Download presentation
Presentation is loading. Please wait.

1
Bio 211- Anatomy and Physiology I Today’s topics Muscular system

2
Organization of muscles and the muscular system Muscle function: 1.Movement – Generate force to move bones Generate force to move body contents 2.Stability – Muscle tone resists force of gravity and helps stabilize some joints Posture 3.Communication – Speaking, writing, facial expressions, etc… 4.Control of body openings – SPHINCTER muscles control openings of eyes, mouth, digestive system, urethra, anus 5.Heat production - Muscles generate a lot of heat when contracting (85% of body heat) Keeps body around 98.6 degrees – important for enzyme function! These functions may be carried out by skeletal, smooth, or cardiac muscle, but we’ll mostly focus on skeletal muscle

3
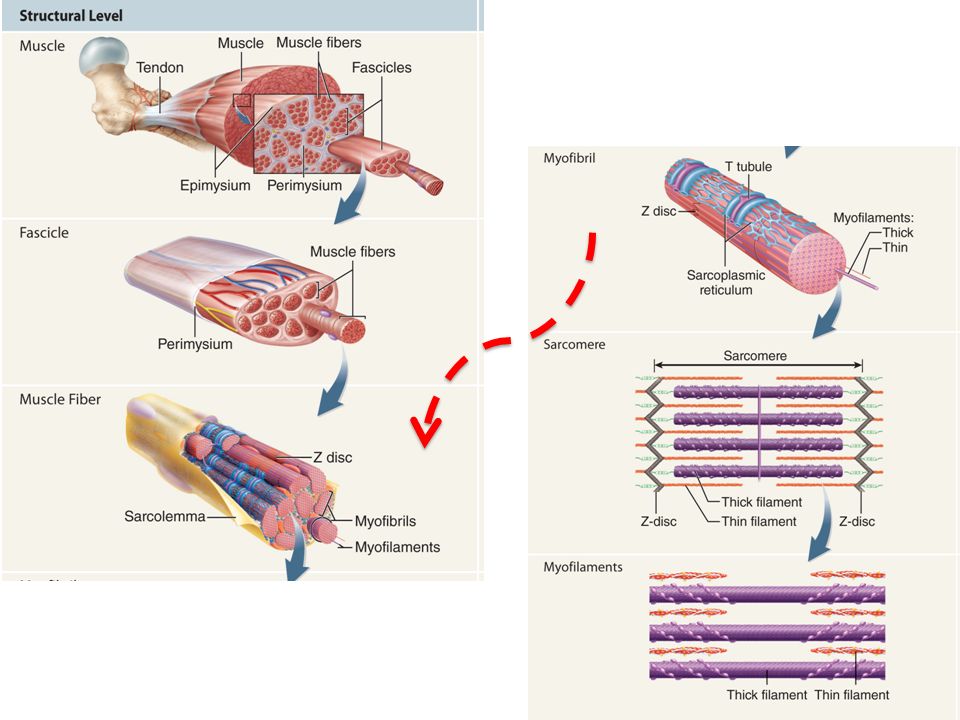
Connective tissues of a muscle A muscle (biceps brachii, pectoralis, etc..) is an organ made up of 1,000s of individual muscle cells (MUSCLE FIBERS) that work together to generate force Muscles contain a great deal of connective tissue Keeps muscle fibers organized Allows attachment to bone/other structures
 is an organ made up of 1,000s of individual muscle cells (MUSCLE FIBERS) that work together to generate force Muscles contain a great deal of connective tissue Keeps muscle fibers organized Allows attachment to bone/other structures")
4
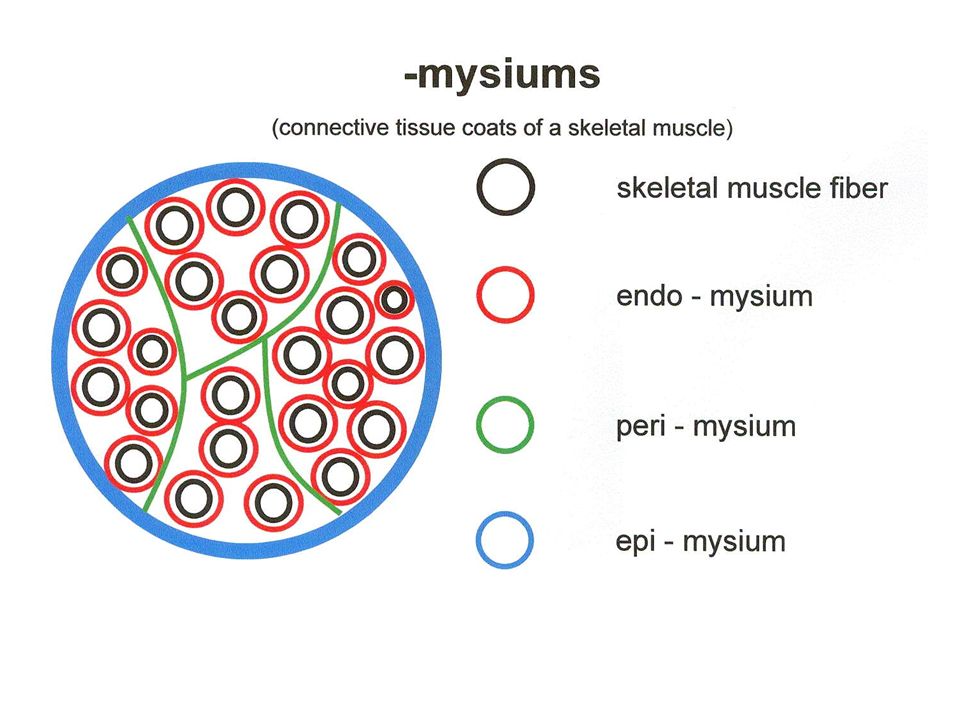
Connective tissues of a muscle Innermost Thinnest Outermost Thickest 1.ENDOMYSIUM : areolar connective tissue that surrounds EACH muscle fiber Muscle fibers are grouped together in bundles called FASCICLES 2.PERIMYSIUM : Thicker connective tissue that surrounds EACH FASCICLE Fascicles can be seen with the naked eye as “grains” 3.EPIMYSIUM : Thicker connective tissue that surrounds the ENTIRE MUSCLE Keeps fascicles bundled together 4.FASCIA : Connective tissue that surrounds and separates muscles from other tissues DEEP FASCIA – separates layers of muscles from one another SUPERFICIAL FASCIA – contains adipose tissue and separates muscles from overlying skin

6
Connective tissues of a muscle Attachment of muscle to bones 1.DIRECT : connective tissue surrounding muscle (epimysium) fuses directly with the periosteum of the bone Intercostal and some facial muscles attach this way 2.INDIRECT : Epimysium and deep facia transitions into a tendon which then inserts into the periosteum of a bone Very common, creates a physical gap between muscle and bone 3.APONEUROSIS : Broad, sheet-like tendon that fuses muscle to bone (abdominal muscles) Aponeurosis of external oblique
 fuses directly with the periosteum of the bone Intercostal and some facial muscles attach this way 2.INDIRECT : Epimysium and deep facia transitions into a tendon which then inserts into the periosteum of a bone Very common, creates a physical gap between muscle and bone 3.APONEUROSIS : Broad, sheet-like tendon that fuses muscle to bone (abdominal muscles) Aponeurosis of external oblique")
7
Muscle anatomy terminology Most muscles of the body are attached to different bones on either end. The contraction of the muscle causes the movement of one of the bones ORIGIN – the stationary attachment point of a muscle to a bone – this bone does not move when muscle is contracted INSERTION : the moveable attachment point of a muscle to a bone – this bone moves when the muscle contracts

8
Actions of groups of muscles Very often, two or more muscles act together (or against each other) to produce movement at a joint AGONIST : muscle that produces the most force during a particular movement SYNERGIST : muscle that aids the agonist in producing a given movement Usually insertion is different than that of the agonist – helps ensure a stable joint and prevent damage to a muscle or joint ANTAGONIST : muscle that opposes the agonist – creates movement in a joint that is in the opposite direction of the agonist FIXATOR : a muscle that prevents movement of a bone – Ex.: rhomboids prevent movement of the scapula when the biceps brachii contracts
 to produce movement at a joint AGONIST : muscle that produces the most force during a particular movement SYNERGIST : muscle that aids the agonist in producing a given movement Usually insertion is different than that of the agonist – helps ensure a stable joint and prevent damage to a muscle or joint ANTAGONIST : muscle that opposes the agonist – creates movement in a joint that is in the opposite direction of the agonist FIXATOR : a muscle that prevents movement of a bone – Ex.: rhomboids prevent movement of the scapula when the biceps brachii contracts")
9
Skeletal muscle histology Reminder – generally, skeletal muscle is VOLUNTARY, while all smooth and cardiac muscle is INVOLUNTARY There are a few exceptions like the muscles of the diaphragm which are skeletal AND involuntary (more than one, located near plasma membrane) (long and cylindrical) (thin C.T. that surrounds EACH muscle fiber) (due to overlapping of actin and myosin) Muscle fibers have several nuclei because they come from the fusion of several MYOBLASTS Occurs during embryonic development Undifferentiated myoblasts exist in adults as SATELLITE CELLS (a form of stem cell)
 (due to overlapping of actin and myosin) Muscle fibers have several nuclei because they come from the fusion of several MYOBLASTS Occurs during embryonic development Undifferentiated myoblasts exist in adults as SATELLITE CELLS (a form of stem cell).")
10
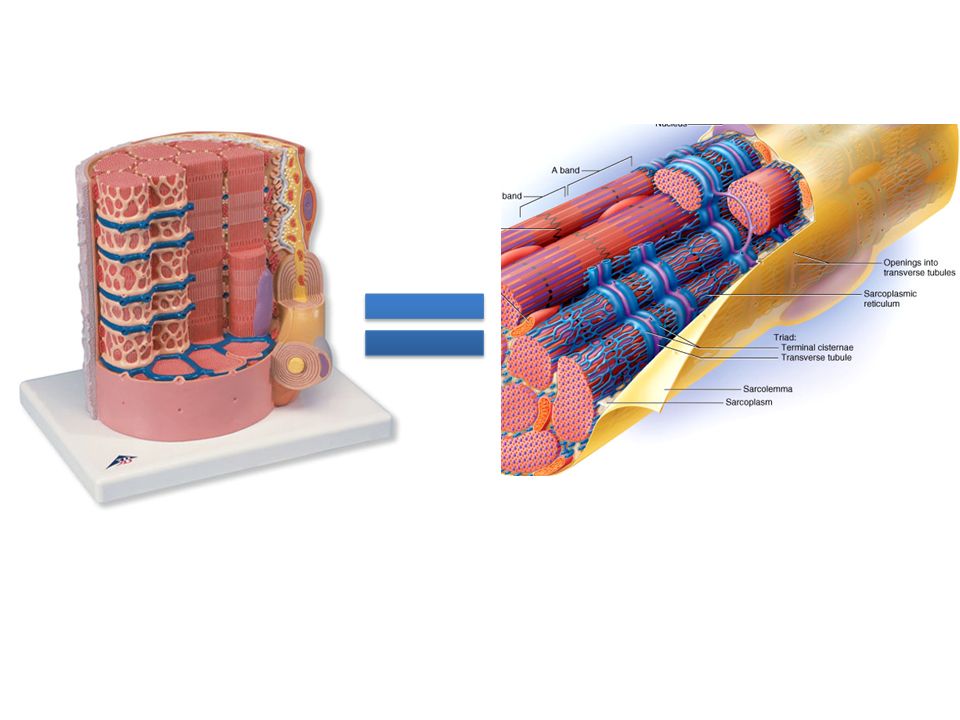
Microscopic anatomy of a muscle fiber Like all cells, muscle cells have a nucleus, plasma membrane, cytoplasm, etc… In muscle cells these structures have special names: Plasma membrane = SARCOLEMMA Cytoplasm = SARCOPLASM Endoplasmic reticulum = SARCOPLASMIC RETICULUM - VERY important to muscle fiber function because it acts as the internal storage site of Ca 2+ (needed for muscle contraction) NOTE: this is ONE individual muscle fiber
 NOTE: this is ONE individual muscle fiber")
11
Microscopic anatomy of a muscle fiber Most of the interior of the cell is packed with protein filaments needed for contraction– bundles of filaments (called MYOFILAMENTS) combine to form MYOFIBRILS Larger muscle fibers will contain more myofibrils The sarcoplasmic reticulum forms a network around each myofibril (allows rapid and even distribution of Ca 2+ ions) Dilations of the S.R. are known as TERMINAL CISTERNAE (contain a great deal of Ca 2+ ) Myofilaments (Thick/thin/elastic filaments) Myofibrils Muscle Fibers Muscle Tissue
 Myofilaments (Thick/thin/elastic filaments) Myofibrils Muscle Fibers Muscle Tissue.")
13
Microscopic anatomy of a muscle fiber TRANSVERSE TUBULES (T-tubules) small tubes that are created by the infolding of the sarcolemma The T-tubule has two terminal cisternae on either side to form a TRIAD The T-tubule carries electrical signals (nerve impulses) to the interior of the cell and triggers the release of Ca 2+ from the SR and terminal cisternae (needed for contraction) Nerve impulse
 small tubes that are created by the infolding of the sarcolemma The T-tubule has two terminal cisternae on either side to form a TRIAD The T-tubule carries electrical signals (nerve impulses) to the interior of the cell and triggers the release of Ca 2+ from the SR and terminal cisternae (needed for contraction) Nerve impulse")
15
Bio 211- Anatomy and Physiology I Today’s topics Muscular system

16
Molecular structure of myofibrils Each MYOFIBRIL consists of a bundle of long, parallel myofilaments : Thick filaments, thin filaments, elastic filaments Thick Filaments – Made up of hundreds of protein molecules called MYOSIN – shaped like a golf club Myosin molecules are arranged end-to-end in a spiral fashion with the “heads” facing outward Thin filaments – Made up of long winding strands of the protein ACTIN Every molecule of actin has an ACTIVE SITE that can potentially bind to the myosin head When a muscle is relaxed, the active sites of actin are covered up by two proteins called TROPOMYOSIN and TROPONIN – myosin can’t bind to actin and cause contraction Elastic filaments – Smallest of the filaments – made up of protein called TITIN A stretchy filament that anchors the thick filaments in place

17
Molecular structure of myofibrils Myosin and actin are considered CONTRACTILE PROTEINS since they are responsible for the actual shortening of the muscle fiber (contraction) Tropomyosin and troponin are considered REGULATORY PROTEINS since their job is to regulate when and if a muscle cell will contract There are also a number of accessory proteins that also play a role in muscle fiber contraction DYSTROPHIN – a HUGE protein that anchors actin filaments to the endomysium Links the shortening of myofilaments to sarcolemma and external CT This protein is mutated and non-functional in the disease MUSCULAR DYSTROPHY – patients exhibit progressive loss of skeletal muscle function
 Tropomyosin and troponin are considered REGULATORY PROTEINS since their job is to regulate when and if a muscle cell will contract There are also a number of accessory proteins that also play a role in muscle fiber contraction DYSTROPHIN – a HUGE protein that anchors actin filaments to the endomysium Links the shortening of myofilaments to sarcolemma and external CT This protein is mutated and non-functional in the disease MUSCULAR DYSTROPHY – patients exhibit progressive loss of skeletal muscle function")
18
Organization of filaments in a myofibril The thick, thin, and elastic filaments are arranged in a VERY specific, ordered fashion that allows contraction to occur efficiently Z-DISK is a large protein that the thin filaments and elastic filaments are attached to Z-disk is also attached to sarcolemma – helps translate contraction of myofibrils to contraction of muscle fiber Thick filaments are suspended between the thin filaments by elastic filaments Two Z-disks with the associated myofilaments in between is called a SARCOMERE – the functional unit of contraction in a myofibril Muscle cell stimulation brings the Z-disks closer together, shortening the length of the sarcomere = CONTRACTION!!

19
Organization of filaments in a myofibril Remember that skeletal muscle contains STRIATIONS – alternating bands of light and dark The I-band (light band) of the sarcomere contains only elastic filaments and thin filaments – more light passes through when looking under microscope The A-band (dark band) contains overlapping thin, thick, and elastic filaments LIGHT = I -band DARK = A-band The H-band is the region in the middle where there are only thick filaments since the thin filaments do not extend that far
 of the sarcomere contains only elastic filaments and thin filaments – more light passes through when looking under microscope The A-band (dark band) contains overlapping thin, thick, and elastic filaments LIGHT = I -band DARK = A-band The H-band is the region in the middle where there are only thick filaments since the thin filaments do not extend that far")
20
Organization and association of sarcomere elements with linking proteins, sarcolemma, and endomysium Linking proteins such as dystrophin allows muscle cell to translate shortening of sarcomere into shortening of muscle cell and shortening of muscle tissue

21
Role of the nervous system in muscle contraction A stimulus from the nervous system is needed to “excite” each muscle fiber to contract Specialized neurons that trigger muscle contraction are known as MOTOR NEURONS As the axon of a motor neuron approaches the muscle it branches out into hundreds of smaller fibers – allows one neuron to stimulate MANY different muscle fibers This arrangement helps coordinate the timing of the contractions of the 1000s of muscle fibers found in a muscle Although each neuron can stimulate 100s of muscle fibers, one muscle fiber only receives input from one neuron – prevents the muscle fiber from getting “mixed signals”

22
Role of the nervous system in muscle contraction When a motor neuron sends an impulse, ALL of the muscle fibers associated with it contract at the same time – this group of muscle fibers is called a MOTOR UNIT Large muscles may have 100s of different motor units Overall strength of muscle contraction depends on # of motor units stimulated The muscle fibers of a motor unit are spread out over a large area This allows a muscle to contract uniformly during a weak contraction – don’t only want one little area to contract (very inefficient) The process of adding more motor units during a strong contraction = RECRUITMENT Ability to vary # of motor units AND size of motor units allows us to vary force of contraction!!!
 The process of adding more motor units during a strong contraction = RECRUITMENT Ability to vary # of motor units AND size of motor units allows us to vary force of contraction!!!")
23
Role of the nervous system in muscle contraction Having many motor units is important for repeated or sustained muscle contractions Eventually, some motor units fatigue and others will take over (like shift work) Not all motor units contain the same # of muscle fibers Activation of smaller motor units allow for weaker muscle contractions Activation of larger motor units lead to strong contractions Weak nerve impulse = fewer and smaller motor units → weak contraction, fine movement Strong nerve impulses = more, larger motor units → strong contraction, large movements
 Not all motor units contain the same # of muscle fibers Activation of smaller motor units allow for weaker muscle contractions Activation of larger motor units lead to strong contractions Weak nerve impulse = fewer and smaller motor units → weak contraction, fine movement Strong nerve impulses = more, larger motor units → strong contraction, large movements")
24
The neuromuscular junction How does a nerve “tell” a muscle when and how to contract??? Neurons “talk” to muscle fibers via the NUEROMUSCULAR JUNCTION Interaction between a motor neuron and a muscle fiber is a SYNAPSE – cells are separated by a small gap called the SYNAPTIC CLEFT Neurons communicate with muscle cells using a NEUROTRANSMITTER: ACETYLCHOLINE (ACh) Acts like a chemical messenger that carries the signal from the neuron to the muscle cell
 Acts like a chemical messenger that carries the signal from the neuron to the muscle cell.")
25
The neuromuscular junction Motor neurons a large bulb at their ends called a SYNAPTIC KNOB (or SYNAPITC BULB) This knob contains vesicles filled with ACh Region of the sarcolemma that contains ACh receptors is called MOTOR END PLATE ACh is released by motor neuron and travels across synaptic cleft ACh binds to receptors on sarcolemma setting off chain of events leading to muscle contraction ACh is degraded by ACETYLCHOLINESTERASE, stimulation stops and allows muscle to relax
 This knob contains vesicles filled with ACh Region of the sarcolemma that contains ACh receptors is called MOTOR END PLATE ACh is released by motor neuron and travels across synaptic cleft ACh binds to receptors on sarcolemma setting off chain of events leading to muscle contraction ACh is degraded by ACETYLCHOLINESTERASE, stimulation stops and allows muscle to relax")
26
Excitation of cells of the muscular system Motor neurons and muscle fibers are both known as ELECTRICALLY EXCITABLE The voltage across the cell membrane changes when the cell is stimulated Normally, a resting cell contains a negative charge INSIDE the cell as compared to the positive charge OUTSIDE Negatively charged ions, proteins, and DNA inside, lots of Na+ outside There is SOME K+ inside the cell but not enough to neutralize the negative charge When there is a charge difference across the cell membrane, the cell is said to be POLARIZED Negative = iNside, pOsitive = Outside The electrochemical Na+ gradient (more outside, less inside) is maintained by the Na+/K+ ATPase DNA, protein, phosphate Na+ _ _ _ _ _ _ _ K+
 is maintained by the Na+/K+ ATPase DNA, protein, phosphate Na+ _ _ _ _ _ _ _ K+")
27
Excitation of cells of the muscular system Stimulation of a muscle cell leads to DEPOLARIZATION of the cell membrane Stimulus causes Na+ ion channels to open Na+ rushes in (down conc. gradient) For a BRIEF period the inside of the cell becomes positively charged Depolarization causes Na+ channels to close and K+ channels to open K+ rushes out (down its conc. gradient) Loss of (+) charged K+ allows inside of cell to regain negative charge Return of membrane charge (− inside, + outside) is called REPOLARIZATION The change of the cell interior from (–) to (+) and then back to (-) is known as an ACTION POTENTIAL DNA, protein, phosphate Na+ _ _ _ _ _ _ K+ DNA, protein, phosphate Na+ _ _ _ _ _ _ K+ DNA, protein, phosphate Na+ _ _ _ _ _ _ _ K+ POLARIZED REPOLARIZED DEPOLARIZED
 For a BRIEF period the inside of the cell becomes positively charged Depolarization causes Na+ channels to close and K+ channels to open K+ rushes out (down its conc. gradient) Loss of (+) charged K+ allows inside of cell to regain negative charge Return of membrane charge (− inside, + outside) is called REPOLARIZATION The change of the cell interior from (–) to (+) and then back to (-) is known as an ACTION POTENTIAL DNA, protein, phosphate Na+ _ _ _ _ _ _ K+ DNA, protein, phosphate Na+ _ _ _ _ _ _ K+ DNA, protein, phosphate Na+ _ _ _ _ _ _ _ K+ POLARIZED REPOLARIZED DEPOLARIZED.")
28
Localization of an action potential An action potential is a LOCAL occurrence at a specific region of the cell membrane Only a small part of the muscle cell membrane is depolarized at a given time Think of “The Wave” at a sports event NERVE OR MUSCLE FIBER DEPOLARIZATION NERVE OR MUSCLE FIBER REPOLARIZATION Depolarization of a region is IMMEDIATELY followed by repolarization Necessary to allow a nerve or muscle cell to be re-stimulated Depolarization of one region triggers the depolarization of a region next to it Allows action potentials to be self-propagating

29
Excitation of skeletal muscle Nerve impulse arrives at synaptic bulb Entrance of Ca 2+ into synaptic bulb triggers exocytosis of ACh-containing vesicles ACh travels across synaptic cleft and binds to ACh receptors on sarcolemma The region of the muscle fiber where ACh binds is called the MOTOR END PLATE-the muscle part of the neuromuscular junction Motor end plate

30
Excitation of skeletal muscle The ACh receptors actually play two roles : ACh receptor AND ion channel LIGAND-GATED ION CHANNEL – opens up when ACh binds to it Binding of ACh (ligand) to its receptor causes ion channel to open Na+ ions rush IN, causing voltage across membrane to reverse (depolarize) This LOCAL fluctuation of membrane voltage (at the motor end plate) leads to the activation of VOLTAGE-GATED ion channels on the REST of the muscle fiber
 to its receptor causes ion channel to open Na+ ions rush IN, causing voltage across membrane to reverse (depolarize) This LOCAL fluctuation of membrane voltage (at the motor end plate) leads to the activation of VOLTAGE-GATED ion channels on the REST of the muscle fiber")
31
OUTSIDE (+) INSIDE (-) Ligand Na+ Ligand-gated -vs- Voltage gated ion channels Ligand Na+ Ligand gated ion channels are ion channels that open up in response to a ligand (ACh) binding to them These channels will only be found at the motor end plate of a muscle cell When ACh binds to the channel it opens up letting Na+ enter the cell Voltage gated ion channels are ion channels that require a change in membrane voltage to open and cause depolarization These channels sense changes in membrane voltage (a nearby depolarization) that triggers them to open up and let Na+ enter the cell Found over the rest of the muscle cell (but not at motor end plate) These channels are responsible for propagating the action potential over the surface of a muscle cell Motor end plateThe rest of the muscle cell
 INSIDE (-) Ligand Na+ Ligand-gated -vs- Voltage gated ion channels Ligand Na+ Ligand gated ion channels are ion channels that open up in response to a ligand (ACh) binding to them These channels will only be found at the motor end plate of a muscle cell When ACh binds to the channel it opens up letting Na+ enter the cell Voltage gated ion channels are ion channels that require a change in membrane voltage to open and cause depolarization These channels sense changes in membrane voltage (a nearby depolarization) that triggers them to open up and let Na+ enter the cell Found over the rest of the muscle cell (but not at motor end plate) These channels are responsible for propagating the action potential over the surface of a muscle cell Motor end plateThe rest of the muscle cell")
32
Excitation/Contraction coupling This is the process by which we translate the action potential on the sarcolemma to the contraction of myofilaments inside the muscle cell Action potentials that leave the motor end plate spread out over the entire surface of the sarcolemma and travel down T-tubules Dependent on voltage gated ion channels These action potentials trigger the opening of Ca 2+ channels in the SR allowing Ca 2+ to exit the SR and enter the sarcoplasm (where the myofilaments are located)
")
33
Excitation/Contraction coupling Ca 2+ released from the SR binds to troponin on the thin filaments and causes troponin to change shape and expose active sites on actin molecules The exposure of the active sites on actin (thin filaments) makes them available to bind to the heads of the myosin molecules (thick filaments)
 makes them available to bind to the heads of the myosin molecules (thick filaments)")
34
Contraction This is the process by which the muscle fiber generates tension (force) and is capable of contraction Sliding filament theory States that during contraction, myofilaments (thick and thin) slide past each other thereby generating tension and pulling the Z-disks closer to each other → shortening of the sarcomere
 and is capable of contraction Sliding filament theory States that during contraction, myofilaments (thick and thin) slide past each other thereby generating tension and pulling the Z-disks closer to each other → shortening of the sarcomere")
35
Contraction In the 1 st step of contraction, the myosin head binds one molecule of ATP The enzyme MYOSIN ATPase hydrolyzes ATP, releasing energy (ATP→ADP+P +ENERGY) This energy is used to activate the myosin head The myosin head “reaches” out to bind to the active site of actin In the 2 nd step, the myosin head binds to the exposed active site of actin, creating a CROSS-BRIDGE
 This energy is used to activate the myosin head The myosin head reaches out to bind to the active site of actin In the 2 nd step, the myosin head binds to the exposed active site of actin, creating a CROSS-BRIDGE")
36
Contraction After binding to actin, the myosin head releases the ADP+ P causing it to return to its original conformation This pulls on the thin filament, bringing the Z-disks closer Binding of a new molecule of ATP causes the myosin head to release the actin molecule Hydrolysis of another ATP then allows the myosin head to “reach” out to another actin molecule farther down the thin filament Each “ratcheting” of the myosin head uses one ATP Each myosin head ratchets 5X per second 1000s of myosin heads work together to shorten each muscle fiber – Contraction uses a HUGE amount of ATP!!!

37
Relaxation Once the contraction is finished, the motor neuron stops releasing ACh Remaining ACh bound to the receptors on the sarcolemma is degraded by ACETYLCHOLINESTERASE This prevents action potentials from being generated at the motor end plate Without stimulation, Ca 2+ is pumped back INTO the SR – active transport (uses ATP) As Ca 2+ diffuses away from troponin, the tropomyosin molecules cover up the active sites of actin again – prevents myosin from binding Without myosin binding, the thin filaments slide BACK over the thick filaments and the sarcomere lengthens (returns to resting length)
 As Ca 2+ diffuses away from troponin, the tropomyosin molecules cover up the active sites of actin again – prevents myosin from binding Without myosin binding, the thin filaments slide BACK over the thick filaments and the sarcomere lengthens (returns to resting length)")
38
Summary 1.Arrival of action potential at synaptic bulb of motor neuron causes release of ACh across synaptic cleft (process requires Ca 2+ ) 2.Binding of ACh to sarcolemma at motor endplate creates local action potential (dependent on the availability of ACh from motor neuron) 3.Action potential at motor end plate spreads out over sarcolemma and down T-tubule to SR (these action potentials are generated by voltage gated ion channels and are dependent on the initial ACh-dependent action potential) 4.Release of Ca 2+ from SR allows active sites of actin to be bound by myosin heads 5.“Ratcheting” of the myosin heads is dependent on the energy released from ATP hydrolysis 6.“Ratcheting” of myosin causes thin filaments to slide over the thick filaments – brings Z- disks closer to each other (sarcomere shortening) 7.End of motor neuron stimulation causes Ca 2+ to be pumped back into SR (diffuses away from troponin) – active transport of Ca 2+ requires ATP 8.Myosin releases actin and allows thin filaments to slide back over thick filaments and return to resting position (sarcomere lengthens) Be sure to understand and remember which steps in the entire process require ATP, ACh, or Ca 2+ !!!!!!
 2.Binding of ACh to sarcolemma at motor endplate creates local action potential (dependent on the availability of ACh from motor neuron) 3.Action potential at motor end plate spreads out over sarcolemma and down T-tubule to SR (these action potentials are generated by voltage gated ion channels and are dependent on the initial ACh-dependent action potential) 4.Release of Ca 2+ from SR allows active sites of actin to be bound by myosin heads 5. Ratcheting of the myosin heads is dependent on the energy released from ATP hydrolysis 6. Ratcheting of myosin causes thin filaments to slide over the thick filaments – brings Z- disks closer to each other (sarcomere shortening) 7.End of motor neuron stimulation causes Ca 2+ to be pumped back into SR (diffuses away from troponin) – active transport of Ca 2+ requires ATP 8.Myosin releases actin and allows thin filaments to slide back over thick filaments and return to resting position (sarcomere lengthens) Be sure to understand and remember which steps in the entire process require ATP, ACh, or Ca 2+ !!!!!!")
39
Bio 211- Anatomy and Physiology I Today’s topics Muscular system

40
Action potential and muscle physiology animations Action potential at Neuromuscular junction Action potentials and muscle contraction Crossbridge formation and contraction contraction of the sarcomere

41
What factors are needed and when??? Ca 2+ ACh ATP ACh release: Ca 2+ triggers exocytosis of ACh from synaptic bulb Crossbridge formation: Ca 2+ binds to troponin causing tropomyosin to shift positions and expose active sites of actin Action potential generation: ACh binds to receptors on sarcolemma of motor end plate causing Na+ channels to open and lead to depolarization of sarcolemma Without this initial action potential there can be NO MUSCLE CONTRACTION Maintenance of cell polarization: Na+/K+ ATPase requires ATP to maintain Na+ gradient thereby keeping interior of cell negatively charged Muscle fiber contraction: Myosin heads use ATP to “reach” out and attach to active sites of actin Muscle fiber relaxation: Ca 2+ pumps use active transport to pump Ca 2+ BACK into the SR

42
Length-Tension relationship and muscle tone The amount of force or tension a muscle can generate is related to how contracted or stretched it was prior to stimulation A B C A -OPTIMAL RESTING LENGTH the thin filaments are just overlapping with all the myosin heads → maximal contraction B – When the muscle is overly contracted the thin filaments overlap the thick filaments so much that the thick filaments are nearly butting up against the Z-disk → no room for additional sarcomere shortening C – When the muscle is overly stretched only a SMALL part of the thin filament overlaps with the thick filaments → myosin heads on thick filaments can’t “get a hold” of thin filaments

43
Muscle tone Even at rest, muscles maintain a certain degree of contraction called MUSCLE TONE This partial contraction is the result of a spinal reflex involving stretch receptors buried within the muscle tissue This allows the sarcomeres of the muscle fibers to remain at their optimal length Muscle tone is important for posture, balance, and joint stability Muscles of the trunk and legs keep us centered and upright WITHOUT conscious effort Muscle tone in deltoid and biceps brachii helps stabilize shoulder joint Muscle Spindle Motor neuron

44
Whole muscle physiology In order for a muscle to contract we need a sufficiently strong stimulus that can trigger the contraction of the muscle fibers in a motor unit An single impulse may not be strong enough to trigger a contraction, requiring REPEATED or STRONGER nerve impulses to reach THRESHOLD (i.e., cause enough Ca 2+ release to trigger sarcomere shortening) The period of time between the initial stimulus and the beginning of muscle contraction is called the LATENT PERIOD (only a few milliseconds) The cycle of contraction and relaxation is known as a MUSCLE TWITCH
 The period of time between the initial stimulus and the beginning of muscle contraction is called the LATENT PERIOD (only a few milliseconds) The cycle of contraction and relaxation is known as a MUSCLE TWITCH")
45
Whole muscle physiology During the CONTRACTION phase: Sarcomeres shorten → muscle fibers shorten → whole muscle contracts This phase is relatively short because the SR rapidly reabsorbs the Ca 2+ This is the phase where the muscle generates tension and force (pulling on tendons or bone of origin and insertion) During the RELAXATION phase: Ca 2+ is reabsorbed by the SR Troponin and tropomyosin cover up active sites on actin (no more crossbridges) Thin filaments slide BACK over thick filaments Sarcomere lengthens → muscle fiber lengthens → muscle relaxes
 During the RELAXATION phase: Ca 2+ is reabsorbed by the SR Troponin and tropomyosin cover up active sites on actin (no more crossbridges) Thin filaments slide BACK over thick filaments Sarcomere lengthens → muscle fiber lengthens → muscle relaxes")
46
Strength of Muscle Twitches Earlier we talked about how muscle contraction is dependent on: 1. # of motor units involved 2.Size of motor units involved 3.Strength of nerve impulse We can also alter muscle contraction via strength of muscle fiber contraction ELECTRICAL STIMULATION (excitation) of the fiber is “all-or-none”, but contraction strength varies with: a)Strength of stimulation (nerve impulse or voltage) b)Stimulation frequency c)Degree of stretch before stimulation d)Temperature (warm enhances, but extremes inhibit contraction) e)Muscle pH (low pH inhibits contraction-decreases myosin action) f)Hydration (proper hydration of cell required for organization of myofibrils) Concentration of Ca 2+ ions in the inside of the cell correlates with strength of muscle cell contraction More Ca 2+ = Stronger contraction
 of the fiber is all-or-none , but contraction strength varies with: a)Strength of stimulation (nerve impulse or voltage) b)Stimulation frequency c)Degree of stretch before stimulation d)Temperature (warm enhances, but extremes inhibit contraction) e)Muscle pH (low pH inhibits contraction-decreases myosin action) f)Hydration (proper hydration of cell required for organization of myofibrils) Concentration of Ca 2+ ions in the inside of the cell correlates with strength of muscle cell contraction More Ca 2+ = Stronger contraction.")
47
Strength of Muscle Twitches Strength of stimulation An increase in stimulation strength causes more axons within the motor nerve to be stimulated Once THRESHOLD is reached : Greater # of axons stimulated = Greater nerve impulse = Greater # of motor units stimulated The addition of more motor units involved in a contraction is known as RECRUITMENT Beyond a certain point, additional stimulation (or voltage) will not increase muscle contraction because all fibers are contracting maximally Note : NEURON = one individual neural cell NERVE = a bundle of axons from many neurons Larger nerves contain axons from more neurons
 will not increase muscle contraction because all fibers are contracting maximally Note : NEURON = one individual neural cell NERVE = a bundle of axons from many neurons Larger nerves contain axons from more neurons")
48
Frequency of stimulation Repeated, rapid stimulation causes Ca 2+ levels to increase in the muscle fiber → stronger muscle fiber contraction (more exposed active sites on actin for myosin binding) Rapid stimulation prevents all of the Ca 2+ from being reabsorbed by SR→Ca 2+ accumulates→stronger contraction (TREPPE EFFECT) As a muscle twitches rapidly, it generates heat, allowing myosin heads to “ratchet” more efficiently→stronger contraction Strength of Muscle Twitches
 Rapid stimulation prevents all of the Ca 2+ from being reabsorbed by SR→Ca 2+ accumulates→stronger contraction (TREPPE EFFECT) As a muscle twitches rapidly, it generates heat, allowing myosin heads to ratchet more efficiently→stronger contraction Strength of Muscle Twitches")
49
Frequency of stimulation When stimulation is very rapid (faster than needed for Treppe effect), one muscle contraction begins before the previous one ends – overlapping contractions As a result, successive nerve impulses stimulate a partially contracted muscle fiber → stronger contraction (INCOMPLETE TETANUS) If stimulation rate continues to increase, the muscle does not have time to relax at all leading to a sustained, continuous contraction (COMPLETE TENTANUS) – almost never occurs in our bodies though……..
, one muscle contraction begins before the previous one ends – overlapping contractions As a result, successive nerve impulses stimulate a partially contracted muscle fiber → stronger contraction (INCOMPLETE TETANUS) If stimulation rate continues to increase, the muscle does not have time to relax at all leading to a sustained, continuous contraction (COMPLETE TENTANUS) – almost never occurs in our bodies though……..")
50
Bio 211- Anatomy and Physiology I Today’s topics Muscular system

51
Different types of muscle contractions Not every muscle contraction results in shortening of the muscle length and movement of a body part Muscle contraction technically refers to the generation of force and tension within a muscle Isometric contraction (constant length) – muscle contracts (develops tension) but DOES NOT SHORTEN in length Isotonic contraction (constant force) – muscle contracts (develops tension) and may either lengthen or shorten depending on force of contraction Both of these contractions are in play when we lift a weight At this point muscle tension exceeds the load and the muscle is allowed to shorten
 – muscle contracts (develops tension) but DOES NOT SHORTEN in length Isotonic contraction (constant force) – muscle contracts (develops tension) and may either lengthen or shorten depending on force of contraction Both of these contractions are in play when we lift a weight At this point muscle tension exceeds the load and the muscle is allowed to shorten")
52
Muscle Metabolism By now we know that ATP is required for at least 3 steps in the contraction of a muscle: 1.Maintenance of polarized nerve and muscle cells (Na+/K+ ATPase) 2.Contraction of myofilaments (“ratcheting” of myosin heads) 3.Relaxation of myofilaments (Active transport of Ca 2+ back into SR) Without a constant supply of ATP muscles CANNOT contract – no other energy source exists Muscles can generate ATP in two different ways depending on the supply of oxygen 1.AEROBIC RESPIRATION – when oxygen is available 2.ANAEROBIC FERMENTATION – when oxygen is NOT available
 2.Contraction of myofilaments ( ratcheting of myosin heads) 3.Relaxation of myofilaments (Active transport of Ca 2+ back into SR) Without a constant supply of ATP muscles CANNOT contract – no other energy source exists Muscles can generate ATP in two different ways depending on the supply of oxygen 1.AEROBIC RESPIRATION – when oxygen is available 2.ANAEROBIC FERMENTATION – when oxygen is NOT available")
53
Aerobic Respiration Aerobic respiration in muscle tissue typically involves the oxidation of glucose (sugar) or fatty acids (lipids) Amino acids from protein can be used to synthesize ATP, but is definitely not the preferred source - we’d rather use them to make new proteins!! This process requires oxygen and has the potential to generate much more ATP than anaerobic fermentation Waste product is CO 2 - easy to get rid of via breathing The key step in this process is called the KREBS CYCLE (citric acid cycle) Glucose and fatty acids are converted into ACETYL CoA that enters the Krebs cycle The Krebs cycle takes place here
 Glucose and fatty acids are converted into ACETYL CoA that enters the Krebs cycle The Krebs cycle takes place here.")
54
KREBS CYCLE A complex series of biochemical reactions that uses Acetyl CoA to generate a small amount of actual ATP and a larger amount of intermediates that are later used (in another complex reaction) to synthesize even more ATP End result is about 36 molecules of ATP per molecule of glucose
 to synthesize even more ATP End result is about 36 molecules of ATP per molecule of glucose")
55
Anaerobic fermentation This process of creating ATP is only used when oxygen is not available in sufficient quantities Only produces about 2-4 molecules of ATP per molecule of glucose Waste product is LACTIC ACID – very toxic Lowers pH of muscle – enzymes don’t work well, crossbridge formation is inhibited

56
As we exercise our muscles, the ATP we need comes from different sources depending on how long we work out Sources of muscle ATP Immediate Energy : During brief, intense exercise (like a sprint) we can’t supply muscles with enough oxygen to keep up with ATP demand simply via breathing Muscles adjust by getting oxygen from MYOGLOBIN (carries oxygen in muscle) This is a very short-term solution (10-15 sec. of maximal exercise) Most of the ATP comes from the activities of MYOKINASE and CREATINE KINASE Myokinase takes phosphate group from one ADP and puts it on another molecule of ADP to create ATP Creatine kinase takes phosphate from CREATINE MONOPHOSPHATE and adds it to ADP to create ATP Remember ATP → ADP+ P + ENERGY
 Most of the ATP comes from the activities of MYOKINASE and CREATINE KINASE Myokinase takes phosphate group from one ADP and puts it on another molecule of ADP to create ATP Creatine kinase takes phosphate from CREATINE MONOPHOSPHATE and adds it to ADP to create ATP Remember ATP → ADP+ P + ENERGY.")
57
Short-term energy: After we use up our immediate energy (obtained via myokinase and creatine kinase) our muscles shift to ANAEROBIC FERMENTATION This system can supply muscles with enough ATP for 20-40 seconds of max. effort Not a great solution since it generates little ATP for how much glucose it uses Also produces lots of lactic acid that inhibits muscle contraction Long-term energy: Eventually our breathing and cardiovascular function catch up and can supply enough oxygen for AEROBIC RESPIRATION Aerobic respiration is much more efficient at producing ATP (about 36 ATPs per glucose), also produces waste product that’s easy to get rid of (CO 2 ) Important for endurance athletes
, also produces waste product that’s easy to get rid of (CO 2 ) Important for endurance athletes.")
58
Creatine Monophosphate Creatine supplementation is very popular among many athletes The idea is that by increasing the amount of creatine monophosphate in your muscle will lead to increased ATP availability Creatine monophosphate donates phosphate (through action of creatine kinase) to ADP to make ATP Creatine also attracts water causing muscles to swell (muscle volumization) Makes muscles larger, but no significant increase in muscle strength (requires increase in muscle protein mass-you need more thick and thin filaments!!!) Good or bad?? Some studies have shown benefits of creatine supplements under direction of a PHYSICIAN or EXERCISE PHYSIOLOGIST Many athletes tend to take TOO MUCH !!! 2-5g/day is about all your body will be able to use Excess creatine is excreted by kidneys in your urine – not good for your kidneys and wastes lots of $$ (of course supplement companies want you to use more!)
.")
59
Bio 211- Anatomy and Physiology I Today’s topics Muscular system

60
Muscle fatigue and endurance Remember from last time that the primary route for generating ATP is through AEROBIC RESPIRATION Anaerobic fermentation is inefficient and produces lactic acid as a byproduct FATIGUE is the progressive weakening of a muscle from prolonged, strenuous use Loss of muscle fiber contractility Causes of physiological fatigue 1.Insufficient energy sources Need acetyl CoA for Kreb’s cycle (to make ATP) Muscle contains lots of glycogen → can be used up during long, strenuous exercise Needed to maintain polarized muscle and nerve cells (Na+/K+ ATPase) Required for crossbridge formation, “ratcheting” of myosin heads 2.Lactic acid buildup Anaerobic fermentation produces lactic acid Lower pH → inefficient protein, enzyme function (weak contraction)
 Muscle contains lots of glycogen → can be used up during long, strenuous exercise Needed to maintain polarized muscle and nerve cells (Na+/K+ ATPase) Required for crossbridge formation, ratcheting of myosin heads 2.Lactic acid buildup Anaerobic fermentation produces lactic acid Lower pH → inefficient protein, enzyme function (weak contraction)")
61
4.Hyperpolarization of muscle and nerve cells Rapid or prolonged contraction causes K+ to accumulate outside cell Causes greater positive charge outside cell (hyperpolarized) 5.Insufficient ACh stores Repeated, prolonged contraction can cause synaptic bulb to run out of ACh Without ACh → no action protential at motor end plate → no action potential throughout rest of muscle cell, T-tubules, etc… Called JUNCTIONAL FATIGUE Causes of physiological fatigue Preventing fatigue is dependent on: Oxygen intake Need oxygen to synthesize ATP efficiently (Krebs cycle) Elite athletes can take in more O 2 than most people – greater lung capacity, greater cardiovascular conditioning, possibly even more hemoglobin in RBCs Energy stores Muscle stores a lot of energy in the form of glycogen (long glucose chains) “Carbohydrate loading” – increases glycogen stores in muscle. BUT, also draws in water to muscle adding weight – may not be great for a long distance runner

62
Oxygen Debt As we exercise our muscles, we use up lots of oxygen that needs to be replenished AFTER we exercise, we continue to breathe heavily for at least several minutes (often longer) in order to : 1.Replenish O 2 stores Replenish O 2 that is bound to myoglobin (used up for immediate energy) Replenish systemic O 2. Used up lots of oxygen dissolved in blood, tissue fluids, hemoglobin 2.Need O 2 to regenerate substrates used by myokinase and creatine kinase (creatine phosphate) – used up for immediate energy 3.Oxidation of lactic acid Lactic acid that builds up during short term exercise can be oxidized back into glucose Helps replenish glycogen stores Turns toxic lactic acid into something useful After exercise, our metabolic rate remains high – need more oxygen to meet post- exercise metabolic demands Exercising to lose weight burns energy during and AFTER exercise
 – used up for immediate energy 3.Oxidation of lactic acid Lactic acid that builds up during short term exercise can be oxidized back into glucose Helps replenish glycogen stores Turns toxic lactic acid into something useful After exercise, our metabolic rate remains high – need more oxygen to meet post- exercise metabolic demands Exercising to lose weight burns energy during and AFTER exercise.")
63
Classes of muscle fibers Skeletal muscle fibers can be divided into 2 main categories – SLOW TWITCH and FAST TWITCH Slow and fast twitch fibers differ in function and metabolic needs, and are more suited for certain functions Nearly all muscles contain a mixture of slow and fast twitch fibers (difference is in proportion) All of the muscle fibers from a given motor unit contain ONLY fast or slow twitch – not mixed Slow Twitch muscle fibers These fibers exhibit a slower twitch cycle – about 100 msec/twitch They possess lots of mitochondria (site of ATP generation), myoglobin (O 2 stores in muscle), and are well supplied with blood vessels Well-suited to aerobic respiration Very resistant to physiological fatigue Muscles with lots of slow twitch fibers have a dark red appearance – “red meat” Since these fibers do not fatigue easily, they are found in higher abundance in muscles requiring long, sustained contractions – calf muscles (walking), back and trunk muscles (posture)
 All of the muscle fibers from a given motor unit contain ONLY fast or slow twitch – not mixed Slow Twitch muscle fibers These fibers exhibit a slower twitch cycle – about 100 msec/twitch They possess lots of mitochondria (site of ATP generation), myoglobin (O 2 stores in muscle), and are well supplied with blood vessels Well-suited to aerobic respiration Very resistant to physiological fatigue Muscles with lots of slow twitch fibers have a dark red appearance – red meat Since these fibers do not fatigue easily, they are found in higher abundance in muscles requiring long, sustained contractions – calf muscles (walking), back and trunk muscles (posture)")
64
Fast twitch muscle fibers These fibers twitch more rapidly – about 7.5 msec/twitch These fibers contain less mitochondria, less myoglobin, and fewer blood vessels, BUT MORE components required for anaerobic fermentation and phosphagen system (myokinase/creatine kinase) – needed for immediate and short term energy More susceptible to physiological fatigue Have a lighter appearance than slow twitch – “white meat” Sarcoplasmic reticulum is more able to quickly release/absorb Ca 2+ - more rapid and forceful contractions Reliance on anaerobic fermentation produces a lot of lactic acid – prone to fatigue Ability to produce short, forceful contractions makes them suited for larger, less used (relatively) muscles – arm, leg muscles
 – needed for immediate and short term energy More susceptible to physiological fatigue Have a lighter appearance than slow twitch – white meat Sarcoplasmic reticulum is more able to quickly release/absorb Ca 2+ - more rapid and forceful contractions Reliance on anaerobic fermentation produces a lot of lactic acid – prone to fatigue Ability to produce short, forceful contractions makes them suited for larger, less used (relatively) muscles – arm, leg muscles")
65
Fast twitch Slow twitch White meat –vs.- dark meat What can you tell about the different muscles of a chicken???

66
Cardiac Muscle Striated like skeletal muscle (overlapping thick and thin filaments) Intercalated disks and mechanical junctions allow one cardiac muscle cell to stimulate its neighbors and remain tightly connected – unlike skeletal muscle Cardiomyocytes don’t necessarily need nervous system input (at least not the way skeletal muscles do) Stimulation of specialized cells trigger contraction of muscle cells throughout the heart (via intercalated disks) Twitches are very slow compared to skeletal muscle (about 250 msec) – allows the heart to contract and expel all the blood it contains Relies almost exclusively on aerobic respiration – very resistant to fatigue (GOOD!) Lots of myoglobin, mitochondria, glycogen stores (around nucleus)
 Intercalated disks and mechanical junctions allow one cardiac muscle cell to stimulate its neighbors and remain tightly connected – unlike skeletal muscle Cardiomyocytes don’t necessarily need nervous system input (at least not the way skeletal muscles do) Stimulation of specialized cells trigger contraction of muscle cells throughout the heart (via intercalated disks) Twitches are very slow compared to skeletal muscle (about 250 msec) – allows the heart to contract and expel all the blood it contains Relies almost exclusively on aerobic respiration – very resistant to fatigue (GOOD!) Lots of myoglobin, mitochondria, glycogen stores (around nucleus)")
67
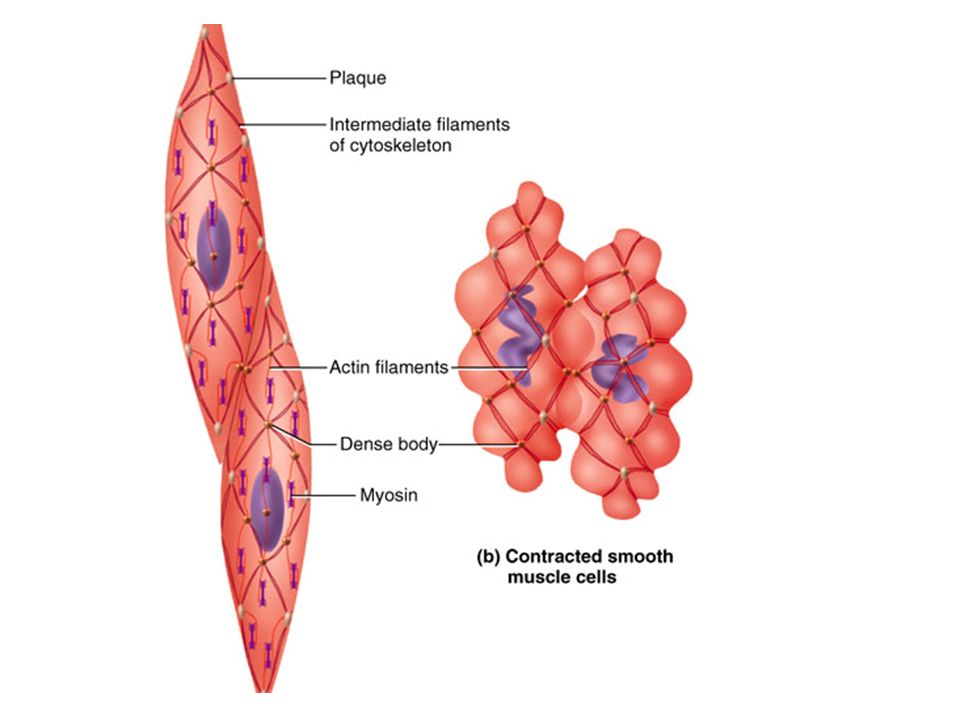
Smooth muscle Tapered cells with NO STRIATIONS Cells DO contain thick and thin filaments, but not arranged in away so that the cell appears striated Thick and thin filaments attach to plasma membrane via a complex internal cytoskeleton and protein masses called DENSE BODIES Thin filaments attach to dense bodies and cytoskeleton instead of Z-disks (skeletal muscle) Need Ca 2+ for contraction, like all muscle Ca 2+ comes primarily from extracellular fluid, NOT FROM SARCOPLASMIC RETICULUM Ca 2+ flows into cell via channels (contraction) Ca 2+ is pumped out during relaxation
 Need Ca 2+ for contraction, like all muscle Ca 2+ comes primarily from extracellular fluid, NOT FROM SARCOPLASMIC RETICULUM Ca 2+ flows into cell via channels (contraction) Ca 2+ is pumped out during relaxation")
69
Stimulation, contraction, relaxation of smooth muscle Like cardiac muscle, smooth muscle is involuntary and doesn’t necessarily need nerve stimulation to contract (but often does) Can also contract in response to hormones, CO 2 levels, changes in pH, stretch, etc… Ca 2+ channels are opened in response to these factors This is how some smooth muscle contraction is regulated in the stomach, intestines, blood vessels, etc… There are 2 main types of smooth muscle: Multiunit smooth muscle – requires innervation from autonomic nervous system (involuntary). Nerve fibers innervate many smooth muscle cells creating a motor unit (like skeletal muscle) Single unit smooth muscle – muscle cells communicate via gap junctions. Nerve fiber releases ACh near one cell and the stimulation is passed to other connected cells (like cardiac cells)
 Single unit smooth muscle – muscle cells communicate via gap junctions. Nerve fiber releases ACh near one cell and the stimulation is passed to other connected cells (like cardiac cells).")
70
Stimulation, contraction, relaxation of smooth muscle Although smooth muscle contains thick and thin filaments and requires Ca 2+ for contraction, it contains no troponin as a regulatory protein Smooth muscle contains a unique protein called calmodulin Associated with THICK FILAMENTS When calmodulin binds Ca 2+ it activates an enzyme that transfers phosphate group from ATP to myosin head (PHOSPHORYLATION=adding phosphate to a protein). Myosin then binds to actin and uses ANOTHER ATP to “ratchet” Process uses 2 ATP molecules as opposed to skeletal muscle (only 1 ATP) Regulation occurs at thick filament, not at thin filament like skeletal muscle When thick filaments “ratchet” they pull on thin filaments that are attached to DENSE BODIES and cytoskeleton → contraction of the muscle cell
 Regulation occurs at thick filament, not at thin filament like skeletal muscle When thick filaments ratchet they pull on thin filaments that are attached to DENSE BODIES and cytoskeleton → contraction of the muscle cell.")
71
Stimulation, contraction, relaxation of smooth muscle As compared to skeletal muscle, smooth muscle twitch is VERY SLOWLY Pumping Ca 2+ into/out of the cell occurs much more slowly without a SR (like skeletal muscle) “Ratcheting” of smooth muscle myosin requires more steps and uses more ATP Phosphorylation/dephosphorylation of myosin is slow For smooth muscle to relax 2 things need to occur: 1.Removal of Ca 2+ - inactivates calmodulin 2. Dephosphorylation of myosin However, in some cases, smooth muscle can stay contracted even in the absence of ATP – allows some muscles to remain contracted without using ATP

Similar presentations



















 –sheet or band of fibrous C.T. under.>")