Download presentation
Presentation is loading. Please wait.

1
Cell Interactions with Biomaterials
Topics: Cell Structure and Components Properties of Cell Components Interaction of Cells with Extracellular Material (ECM) Cell Adhesion and Migration
 Cell Adhesion and Migration.")
2
A successful biomaterial implant must support all the required functions of the attached (or neighboring) cells, including Viability All cell types Communication All cell types Protein synthesis All cell types Proliferation Some cell types Migration Some cell types Activation/differentiation Some cell types Programmed cell death Some cell types

3
Cell Structure 2 types of cells:
Differentiated: perform specific tissue functions Undifferentiated: progenitors for many different cell types Cell tasks are compartmentalized in various organelles. Organelles in all mammal cells include the plasma cell membrane, mitochondria, Golgi apparatus, cytoplasm, lysosome, cytoskeleton, nucleus, and smooth and rough endoplasmic reticulum The Cell

4
Other proteins target specific extracellular molecules
The cell membrane separates the cytoplasm, or cell interior, from the aqueous external environment. It is a bilayered structure made up of phospholipids, or fatty acids with a polar (hydrophilic) head and nonpolar tail. The mitochondria produce the energy for cell functions via a process called oxidative phosphorization. Surrounded by a phospholipid membrane, the mitochondria contains enzymes that help break down molecules Transmembrane proteins span the cell membrane to channel ions into and out of cell and maintain proper cell chemistry In oxidative phosphorization ATP (adenosine triphosphate) is converted to ADP (adenosine diphosphate, an exothermic reaction that releases energy to drive the cellular process. Other proteins target specific extracellular molecules Phospholipid
 head and nonpolar tail. The mitochondria produce the energy for cell functions via a process called oxidative phosphorization. Surrounded by a phospholipid membrane, the mitochondria contains enzymes that help break down molecules. Transmembrane proteins span the cell membrane to channel ions into and out of cell and maintain proper cell chemistry. In oxidative phosphorization ATP (adenosine triphosphate) is converted to ADP (adenosine diphosphate, an exothermic reaction that releases energy to drive the cellular process. Other proteins target specific extracellular molecules. Phospholipid.")
5
The cytoskeleton is made up of three filaments:
Actin microfibrils,~6-8 nm diameter Intermediate filaments ~10 nm diameter Microtubules ~25 nm diameter The Golgi apparatus modifies, sorts and packages proteins for their final destination Made up of proteins, the cytoskeleton Gives the cell shape Can provide cell locomotion Aids in separation/duplication of DNA Lysosomes are specialized vesicles with digestive enzymes

6
The nucleus is the control center for the cell. It contains:
DNA – (deoxyribonucleic acid) that is condensed into chromatin, a complex combination of DNA and protein that makes up chromosomes The nuclear envelope, a bilayer of phospholipid membranes that surrounds the nucleus The outer membrane of the nuclear envelope is contiguous with the endoplasmic reticulum and is connected with the inner membrane at specific locations called nuclear pores. The pores are composed of proteins that form gates to allow only specific molecules in and out of the nucleus Ribosomes, minute round particles composed of RNA and protein that are found in the cytoplasm of living cells and catalyze reactions in which mRNA is used to synthesize proteins. Ribosomes are located in the nucleolus
 that is condensed into chromatin, a complex combination of DNA and protein that makes up chromosomes. The nuclear envelope, a bilayer of phospholipid membranes that surrounds the nucleus. The outer membrane of the nuclear envelope is contiguous with the endoplasmic reticulum and is connected with the inner membrane at specific locations called nuclear pores. The pores are composed of proteins that form gates to allow only specific molecules in and out of the nucleus. Ribosomes, minute round particles composed of RNA and protein that are found in the cytoplasm of living cells and catalyze reactions in which mRNA is used to synthesize proteins. Ribosomes are located in the nucleolus.")
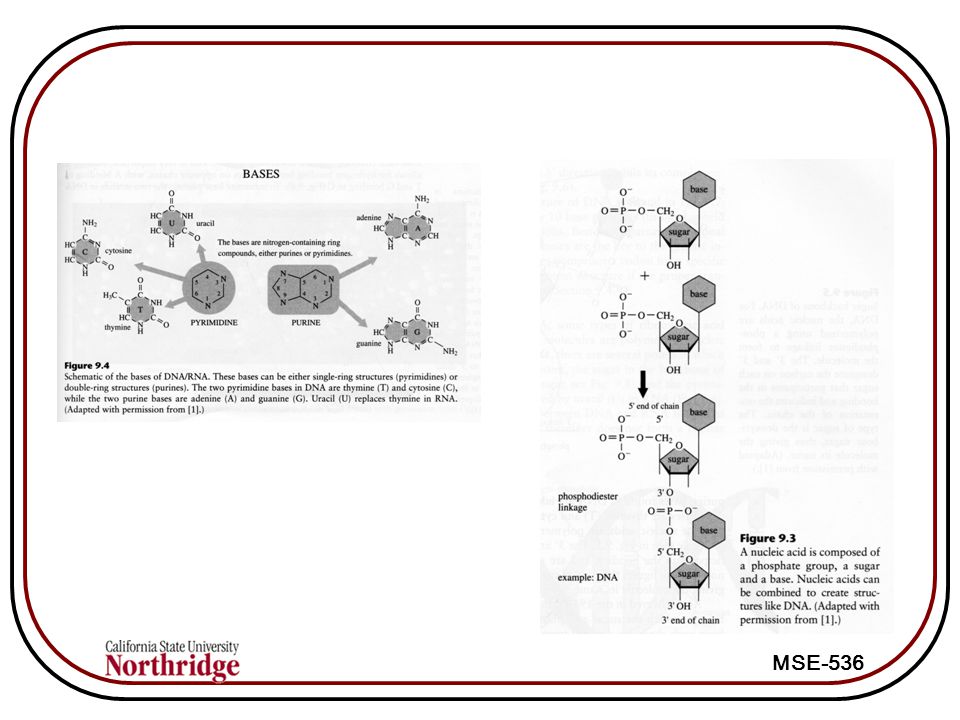
7
DNA DNA (deoxyribonucleic acid) is a polymer of nucleic acid subunits. Each nucleic acid has a phosphate group, a sugar, and a base. DNA contains genes, and forms the template for all proteins synthesized by the cell When a gene is expressed, the cell is actively producing the protein encoded by the gene Genes contain codons which determine the structure of the protein The bases are directed toward the interior, where they form hydrogen bonds with other bases. Bases can be either double ring structures (e.g., purines Adenine (A) or guanine (G)), or single ring (pyrimidines, e.g., thymine (T) or cytosine (C)) DNA
 or guanine (G)), or single ring (pyrimidines, e.g., thymine (T) or cytosine (C)) DNA.")
8
DNA-RNA Protein Synthesis
RNA participates in DNA synthesis and protein production. Three types of RNA: Messenger or mRNA Transfer, or tRNA Ribosomal, or rRNA RNA is similar to DNA, but the sugar in the backbone contains an additional O2, and thymine (T) in DNA has been replaced with uracil (U). RNA is single stranded , and does not form the helical structure of DNA mRNA: messenger RNA; DNA is unzipped, and mRNA strands are synthesized that are complementary to DNA tRNA: serves as an adaptor to combine mRNA strands in the rough ER rRNA: the central component of the ribosome, the function of the rRNA is to provide a mechanism for decoding mRNA into amino acids and to interact with the tRNAs during translation DNA-RNA Protein Synthesis
 in DNA has been replaced with uracil (U). RNA is single stranded , and does not form the helical structure of DNA. mRNA: messenger RNA; DNA is unzipped, and mRNA strands are synthesized that are complementary to DNA. tRNA: serves as an adaptor to combine mRNA strands in the rough ER. rRNA: the central component of the ribosome, the function of the rRNA is to provide a mechanism for decoding mRNA into amino acids and to interact with the tRNAs during translation. DNA-RNA Protein Synthesis.")
9
The outer membrane of the nucleus is connected to the endoplasmic reticulum (ER). The ER is the site of protein synthesis. The ER is made of long, flattened sheets of phospholipids, and may be either rough or smooth. The rough ER has ribosomes attached to the surface that act as catalysts for protein synthesis. The smooth ER is more tubular and does not contain ribosomes. It packages the proteins produced in the nucleolus and rough ER in phospholipids for delivery to the Golgi apparatus. Vesicles transport proteins from the ER to the Golgi, or from the Golgi to the target destination (exocytosis). Specialized vesicles (lyosomes) may also take in and digest particles from the ECM
. Specialized vesicles (lyosomes) may also take in and digest particles from the ECM.")
10
Schematic of the endoplasmic reticulum (ER), which is responsible for protein synthesis. The rough ER, which contains a large number of ribosomes, is attached to the nuclear envelope. The rough ER transforms into the smooth ER away from the nucleus. Pieces of the ER will then split off and transfer to the Golgi apparatus, where the proteins created in the ER are further modified and transported ot their final destinations.

11
Interactions between a cell and its environment can result in cell spreading, migration, communication, differentiation and activation. This is called “outside-in” signaling. Conversely, a cell may secrete molecules or rearrange contacts to alter the ECM. This is called “Inside-out” signaling

12
Types of cell contacts Tight junctions: cells adhere fast to each other – no molecular transport Gap junctions: small hydrophilic channels between cells and membranes Desmosomes: mechanical attachment between two cells. Cells can attach to ECM via hemidesmosomes or focal adhesion. These form strong adhesion to the ECM

13
Cell membrane receptors & ligands
Cellular interactions are facilitated by cell membrane receptors, each of which is specific for a small range of target molecules, or ligands. Common receptor molecules are: Cadherins: responsible for demosomes; a cadherin molecule on one cell binds to a cadherin on another cell. This is homophilic binding Selectins: selectins are like cadherins, but bind to other types of receptors, or heterophilic binding Mucins: participate in heterophilic binding to selectins. Integrins: transmembrane proteins involved in both cell-cell and cell-matrix contacts. Location of cadherins in epithelial cells.. They link to each other to bind cells, and their cytoplasmic regions attach to intermediate filaments linking the ECM to the intracellular environment

14
Cell membrane receptors & ligands
Integrins have two distinct a and b subunits, and are called heterodimers. Variations in the composition of the a and b chains results in selective adhesion to different ligands Other cell adhesion molecules (CAMs) comprise a large group of membrane proteins that mediate cell-cell interactions via both homophilic and heterophilic binding. An example of these other CAMs is the immunoglobulin (Ig) family in the picture at right. The receptors described consist mainly of protein with a small attached carbohydrate (sugar). Similar molecules with small protein and large sugar content are called proteoglycans. Various types of cell membrane receptors: mucins, integrins, selectins, and Ig-cell adhesion molecules (Ig-CAMs).
 comprise a large group of membrane proteins that mediate cell-cell interactions via both homophilic and heterophilic binding. An example of these other CAMs is the immunoglobulin (Ig) family in the picture at right. The receptors described consist mainly of protein with a small attached carbohydrate (sugar). Similar molecules with small protein and large sugar content are called proteoglycans. Various types of cell membrane receptors: mucins, integrins, selectins, and Ig-cell adhesion molecules (Ig-CAMs).")
15
Extracellular Matrix – ECM Environment
Understanding the cell-ECM interaction and response is key to designing new biomaterials The ECM may be thought of as a fiber reinforced composite with fibers made of collagen or elastin, and a matrix made of glycoproteins and proteoglycans ECM video

16
X and Y molecules are often proline and hydroxyproline
Collagen Upon secretion of procollagen from the cell into the ECM, small peptide sequences are cleaved to from the molecule to allow for more efficient packing of collagen molecules into fibrils ( nm). Individual fibers are then assembled into larger fibers (0.5 – 3 mm diameter) Collagen is the most abundant protein in mammals. It is responsible for tissue tensile strength. Collagen is made up of a-polypeptides, or amino acids in a gly-x-y pattern (above). Gly is a small molecule resulting in tight packing X and Y molecules are often proline and hydroxyproline Cell excretes procollagen molecules that self-assemble into fibers (right) Properties of collagen may be manipulated by controlling crosslinking of molecules
. Individual fibers are then assembled into larger fibers (0.5 – 3 mm diameter) Collagen is the most abundant protein in mammals. It is responsible for tissue tensile strength. Collagen is made up of a-polypeptides, or amino acids in a gly-x-y pattern (above). Gly is a small molecule resulting in tight packing. X and Y molecules are often proline and hydroxyproline. Cell excretes procollagen molecules that self-assemble into fibers (right) Properties of collagen may be manipulated by controlling crosslinking of molecules.")
17
Elastins Elastin is responsible for the resiliency and elasticity of the ECM. Elastin is made up of 85% hydrophobic amino acids When relaxed, elastin molecules coil up. When a tensile load is applied they unfold into long chains. The chains are crosslinked to adjacent elastin molecules

18
These molecules attract and interact strongly with water.
Additional ECM molecules include proteoglycans and glycoproteins. These proteins are mainly carbohydrate, with some protein side chains. These molecules attract and interact strongly with water. Carbohydrates in proteoglycans form long chains of polysaccharides called glycosaminoglycans (GAGs) Proteoglycans have several GAGs attached to a protein core. An aqueous environment is favorable to transport and store bioactive molecules. Example of the bottle-brush structure of a large proteoglycan including areas of keratan sulfate and chrondroitin sulfate that are attached to a protein core
 Proteoglycans have several GAGs attached to a protein core. An aqueous environment is favorable to transport and store bioactive molecules. Example of the bottle-brush structure of a large proteoglycan including areas of keratan sulfate and chrondroitin sulfate that are attached to a protein core.")
19
Two molecules that represent glycoproteins are fibronectin and laminin.
Each consists of peptide subunits held together by disulfide bonds and contain many sites for binding to various ECM molecules. Laminin (right) consists of three disulfide linked peptide chains in a loosely woven structure with multiple binding sites Glycoproteins are important in blood clotting (coagulation) since they posses binding domains for heparin and fibrin
 consists of three disulfide linked peptide chains in a loosely woven structure with multiple binding sites. Glycoproteins are important in blood clotting (coagulation) since they posses binding domains for heparin and fibrin.")
20
Cell-Environment Interactions that Affect Cellular Functions
Interactions between cells and cell/ECM can alter cells function and affect gene expression in the nucleus Alterations in gene expression affect four major functions: Cell viability Proliferation Differentiation Protein synthesis and communication Changes in ECM can cause cell death via necrosis or apoptosis due to changes in the local chemistry (e.g. pH), factors that attach to cell membrane leading to cell death. Apoptosis: cell shrinkage followed by fragmentation into vesicles containing small groups of organelles. No inflammatory response Necrosis: cell death from membrane permeability and enzyme leakage, leading to disintegration
, factors that attach to cell membrane leading to cell death. Apoptosis: cell shrinkage followed by fragmentation into vesicles containing small groups of organelles. No inflammatory response. Necrosis: cell death from membrane permeability and enzyme leakage, leading to disintegration.")
21
Mitosis: cell division
Cell classifications Labile: replicate continuously Terminal: terminally differentiated Stable: don’t change once differentiated but can be induced to proliferate The cell cycle is divied into two phases: Mitosis (the M phase) and interphase (G1,S,G2) Mitosis: cell division Interphase: cellular DNA and organelles replicated in preparation for mitosis Go: quiescent phase of stable cells G1: general cell growth S: DNA replicated G2: proteins and structures enabling cell division ar assembled
 and interphase (G1,S,G2) Mitosis: cell division. Interphase: cellular DNA and organelles replicated in preparation for mitosis. Go: quiescent phase of stable cells. G1: general cell growth. S: DNA replicated. G2: proteins and structures enabling cell division ar assembled.")
22
Mitosis is divided into several characteristic periods:
Prophase Metaphase Anaphase Telophase Prophase: dissipation of the nucleolus and formation of the mitotic spindles Metaphase: chromosomes are aligned between two mitotic spindles Anaphase: chromosomes are pulled apart by the spindle microtubules and arrange themselves at the spindle poles Telophase: nuclear envelope begins to reform and the cell starts to undergo cytokinesis Cytokinesis: division of cell cytoplasm

23
Cell Differentiation Progenitor or stem cells can form more than one type of cell The cells produced may be committed or differentiated and can be labile, stable or permanent Or Create additional pluripotent or totipotent cells (produce other or all cell types) Red blood cells generated from hematopoietic stem cell can either replicate itself of differentiate into various cell types. These cells are pluripotent Embryonic stem cells are an example of a totipotent cell
 Red blood cells generated from hematopoietic stem cell can either replicate itself of differentiate into various cell types. These cells are pluripotent. Embryonic stem cells are an example of a totipotent cell.")
24
The commitment and progression of an MSC to and through a specific lineage involves the action of bioactive molecules such as growth factors and cytokines. In the process of differentiation and maturation the cell increases its production of tissue-specific molecules. Terminally differentiated cells may alter their levels of synthesis of matrix molecules to play an increased role in tissue maintenance and homeostasis Mesenchymal stem cells (MSCs) can differentiate into bone, cartilage, muscle, tendon, ligament, and adipose tissue. Differentiation stages can be initiated and controlled by soluble and insoluble elements in their environment and is directly applicable to tissue engineering
 can differentiate into bone, cartilage, muscle, tendon, ligament, and adipose tissue. Differentiation stages can be initiated and controlled by soluble and insoluble elements in their environment and is directly applicable to tissue engineering.")
25
Pixie Dust – Truth or Fiction?
Video 1 Video 2

26
Models for Adhesion, Spreading and Migration
The DLVO theory: (Derjaguin, Landau, Verway and Overbeek) Based on thermodynamics Particles potential energy the sum of attractive and repulsive forces: U = UA + UR Particles approaching a surface reduce their potential energy, and are loosely attached at the secondary minimum – long range electrostatic/van der Waals forces Particles overcoming primary minimum become firmly attached through short range electrostatic forces Model shortcomings: does not include steric repulsion, surface topography/roughness or ligand-receptor interactions
 Based on thermodynamics. Particles potential energy the sum of attractive and repulsive forces: U = UA + UR. Particles approaching a surface reduce their potential energy, and are loosely attached at the secondary minimum – long range electrostatic/van der Waals forces. Particles overcoming primary minimum become firmly attached through short range electrostatic forces. Model shortcomings: does not include steric repulsion, surface topography/roughness or ligand-receptor interactions.")
27
Spreading & Migration Cell spreading: After attachment, cells extend finger-like pseudopodia along surface. The integrin receptors in the cell membrane interact with ligands on the material surface to firmly anchor the cell in place. Cell spreading includes cytoskeleton rearrangement and production/adsorption of adhesive proteins on surface Cell migration: extension of the cell membrane in long pseudopodia is directed by polymerization of actin microfibrils near the leading edge of the cell. (b) the membrane then attaches to the substrate via integrin receptors. (c,d) After the pseudopodia are firmly adhered, there is generation of a contractile force along with a release of rear receptors, leading to forward motion. (e) the integrin receptors are recycled to the leading edge so they may be used again as the process continues.
 the membrane then attaches to the substrate via integrin receptors. (c,d) After the pseudopodia are firmly adhered, there is generation of a contractile force along with a release of rear receptors, leading to forward motion. (e) the integrin receptors are recycled to the leading edge so they may be used again as the process continues.")
28
Tracking cell movements
Plots of cell trajectory, such as the one at right, provide information on cell movement in the form of translocation speed (s) and persistence time (t). Translocation speed is the speed of cell movement over any straight-line portion of the graph, between changes in direction Persistence time is the length of time that the cell moves along the substrate without a drastic change in direction. Measurements of this type are used in mathematical models developed to model cell migration Trajectories of bovine pulmonary artery endothelial cells migrating in a uniform environment. Symbols represent the location of the centroid of each cell at 30 minute intervals. Arrows indicate starting points.
![]()
29
The End

30
Protein Synthesis Receptor-ligand binding can change the function of committed cells which is associated with alterations in the amount of protein synthesized Transcription: Chromatin becomes less compact; RNA “unzips” DNA and synthesizes mRNA strands that are complimentary to DNA then moves into the endoplasmic reticulum (ER) Translation: the process of converting codons from the mRNA to a polypeptide. This takes place in the ER through complex interaction with tRNA. Block diagram of the steps for the creation and modification of proteins. Post Translation: fully formed proteins are combined into various molecules
 Translation: the process of converting codons from the mRNA to a polypeptide. This takes place in the ER through complex interaction with tRNA. Block diagram of the steps for the creation and modification of proteins. Post Translation: fully formed proteins are combined into various molecules.")
31
Summary of the steps of collagen synthesis
Summary of the steps of collagen synthesis. Transcription, translation, synthesis to create the a-chain and the joining of three of thee chains (post translational modification) to create the collagen triple helix occur within the cell. The procollagen molecule is then secreted and assembled into fibrils and finally fibers.
 to create the collagen triple helix occur within the cell. The procollagen molecule is then secreted and assembled into fibrils and finally fibers.")
Similar presentations





 are called _________. Hint: mini-organs.>")






 of animal cells functions.>")









