Download presentation
Presentation is loading. Please wait.

1
Predation-Amensalism Summary
Gause did early predator-prey experiments, and concluded that cycles in nature result from constant migration, because he couldn’t get coexistence in his experiments. Huffaker found habitat complexity allowed coexistence Holling studied Functional response – relationship between prey density and the rate at which an individual predator consumes prey and Numerical response- increase in predator numbers with increases in prey abundance

2
Predation-Amensalism Summary
3 types of functional response curves, I, II, and III Search image- Only when the prey population increases above some threshold level does the predator form a search image and begin to recognize that prey item as a valuable food source. The predator then focuses on and exploits that food source heavily Prudent predation occurs without altruism Predation can cause: changes in size distribution; both decreases and increases in diversity; morphological modifications (spines, mimicry, crypsis)
")
3
Predation-Amensalism Summary
Paine’s exp. led to keystone species concept Optimal foraging-comcerns types of feeding behaviors that maximize food (energy( intake rate Inducible defenses are brought on by predation threat and serve to deter predators Indirect effects of predation: sub-lethal predation; TMII; trophic cascades Parasites are either ecto- or endo-parasites

4
Predation-Amensalism Summary
Parasites exploit host behavior to maximize transmission Host defenses are behavioral, structural or immunological Herbivory is the first step in the transfer of energy in food webs; provides for the cycling of nutrients; and can affect the productivity and structure of plant communities. It increases prevalence of species with: Low nutritive value (low nitrogen) Chemical Defenses (secondary compounds) Structural Defenses (calcareous skeletons) Shifts in functional groups (from fast to slow growers)
 Chemical Defenses (secondary compounds) Structural Defenses (calcareous skeletons) Shifts in functional groups (from fast to slow growers)")
5
Predation-Amensalism Summary
Secondary compounds can deter herbivores but entail tradeoffs in energy allocation Mutualisms- usually have one species providing nutrition while the other provides protection or cleaning services. Can be obligatory or facultative Mutualisms common in tropics and likely evolved from host-parasite relationships Commensalism and trophic amensalism less common

6
Review Questions

7
Community Ecology A community is a group of interacting populations, all living in the same place at the same time the focus is on the interactions between species or populations including competition, predation , succession, invasion, mutualism, predation, etc.

8
Community Structure and Change
Community Structure - a description of the community members (species list) and their relative abundances Community Dynamics - the changes that occur over time and space in a community. (Even though communities have an underlying structure, the structure may change over time
 and their relative abundances. Community Dynamics - the changes that occur over time and space in a community. (Even though communities have an underlying structure, the structure may change over time.")
9
Emergent Properties Properties not predictable from study of component
populations Only apparent at level of community

10
Why is this important? Appropriate unit of study:
- If the community is more than the sum of its parts, then we must study the entire community (holistic approach) - If not then entire picture can be put together from individual pieces (reductionist approach)
 - If not then entire picture can be put together from individual pieces. (reductionist approach)")
11
The Study of Ecological Communities
Properties & patterns Diversity (Number of species) Species’ relative abundances Morphology Succession Processes Disturbances Trophic interactions Competition Mutualism Indirect effects
 Species’ relative abundances. Morphology. Succession. Processes. Disturbances. Trophic interactions. Competition. Mutualism. Indirect effects.")
12
Two Views on Communities
Community as a superorganism (equilibrium community, Clements) Species not replaceable Species need one another to survive Community as a group of individual species (non-equilibrium community, Gleason) Species are replaceable Random association of species
 Species not replaceable. Species need one another to survive. Community as a group of individual species (non-equilibrium community, Gleason) Species are replaceable. Random association of species.")
13
Community Dynamics: Succession
Succession - The change in numbers and kinds of organisms in an area leading to a stable (climax) community. Replacement of communities. Pioneer community - the first community to develop in a successional sequence Sere - any successional community between pioneer and climax community
 community. Replacement of communities. Pioneer community - the first community to develop in a successional sequence. Sere - any successional community between pioneer and climax community.")
14
Types of Succession Primary – situation where barren substrate is available for habitation (inorganic substrates= lava flows/ spreading centers 2. Secondary – occurs in areas where communities have previously existed (after fires or hurricane; much more rapid)
")
15
Succession in community traits
increasing size and longevity of organisms shift from predominantly "r-selected" to predominantly "K-selected" species increasing biomass increasing independence of physical/chemical environment

16
Succession in community traits (2)
decreasing rate of change increasing species diversity and complexity of physical and trophic structures increasing habitat modification and buffering of environmental extremes increasing complexity of energy and nutrient flows increasingly closed system re-cycling of organic and inorganic materials

17
Opposing Views of Communities
Superorganism View (Clements, 1916) tightly evolved, interacting functions as a single organism developmental process (succession) homeostasis (self maintaining – stable) underlying “balance of nature” Individualistic View (Gleason, 1925) randomly assembled Similar resource requirements
 tightly evolved, interacting. functions as a single. organism. developmental process. (succession) homeostasis. (self maintaining – stable) underlying balance of. nature Individualistic View. (Gleason, 1925) randomly assembled. Similar resource. requirements.")
18
Types of Species Early successional Late Successional good colonizers
rapid growth short lived (r-selected) Late Successional poor colonizers slow growth long lived (k-selected) Early Late
 Late Successional. poor colonizers. slow growth. long lived. (k-selected) Early. Late.")
19
Under Equilibrium Models
• Community returns to same position after disturbance • At equilibrium, processes that structure the community produce no net change

20
Equilibrium Theory Single stable state Multiple stable states

21
Outcomes of integrated view
Equilibrium assumed (not tested) Explained succession Super-organism concept widely accepted Dominated community ecology until the 1950’s and beyond
 Explained succession. Super-organism concept widely accepted. Dominated community ecology until the. 1950’s and beyond.")
22
Non-equilibrium models
Disturbance is the norm rather than the exception Disturbed patches provide opportunities for colonization by dispersive species Patchiness promotes diversity on a larger scale

23
Evidence for each view:
Superorganism: remove plants or autotrophs, the community will disappear mutualisms and symbiotic relationships are common (example: herbivore gut bacteria) Non-equilibrium high-level consumers can sometimes be removed without major effects on community disturbances often play a role in determining community structure; these are random
 Non-equilibrium. high-level consumers can sometimes be removed without major effects on community. disturbances often play a role in determining community structure; these are random.")
24
Alternative succession models
Connell and Slatyer (1977) – outlined 3 models: 1. Facilitation – Clementsian succession 2. Tolerance 3. Inhibition Based on effect of initial spp. on subsequent spp.
 – outlined 3 models: 1. Facilitation – Clementsian succession. 2. Tolerance. 3. Inhibition. Based on effect of initial spp. on subsequent spp.")
25
Facilitation Model E Early Stand E L Recruitment Growth Disturbance
Late Successionals only E L Mixed Stand Mortality

26
Tolerance Model E L Mixed Stand E L Recruitment Growth Disturbance
Late Successionals only Mortality

27
Inhibition Model E L Mixed Stand E L Recruitment Growth Disturbance
Late Successionals only L Mortality E E L

28
Succession Can occur without invoking the existence of a “Super-organism” Sequential replacement a consequence of individual species properties

29
Physical disturbance

30
What are the components of disturbance?
The frequency of a disturbance The intensity of the disturbance The timing of the disturbance Influences the availability of larvae to recolonize the disturbed area

31
Intermediate Disturbance Hypothesis
(Connell 1972) Disturbance (eg, tree falls, storms) creates patchiness and new space to be colonized Patchwork is created across the landscape with - early and late successional species - inferior and superior competitors This theory is a non-equilibrium view of how natural communities are structured because landscape is a patchwork of different stages of succession.
 Disturbance (eg, tree falls, storms) creates patchiness. and new space to be colonized. Patchwork is created across the landscape with. - early and late successional species. - inferior and superior competitors. This theory is a non-equilibrium view of how. natural communities are structured because landscape is a. patchwork of different stages of succession.")
32
Intermediate Disturbance Hypothesis (2)
Disturbance is critically important in structuring communities because it can prevent competitively dominant species from excluding others. Weak/infrequent disturbances are insufficient to prevent competitive exclusion Intense/frequent disturbances exclude species sensitive to disturbance Highest diversity might therefore be expected at intermediate frequency or intensities of disturbance

33
Intermediate Disturbance Hypothesis (Connell)
")
34
Top-down vs. bottom-up control
Community structure could be controlled from the bottom-up by nutrients: predators herbivores community structure can be changed by manipulating the lower levels nutrients autotrophs numbers of autotrophs are limited by mineral nutrients

35
Community structure could be controlled top-down by predators (trophic cascade model)
herbivores predators numbers of herbivores are controlled by predators predicts a series of +/- effects if upper levels are manipulated autotrophs nutrients

36
Reintroduction and protection of otters has reduced urchin barrens
Trophic cascades Predation by orcas has increased urchin barrens Reintroduction and protection of otters has reduced urchin barrens

38
Species-area relationships
Species-area curve - the larger the geographic area, the greater the number of species Larger areas have more diverse habitat This can be used to predict how habitat loss may affect key species fig 53.25

39
Species Area Relationships
As a rule of thumb for every 10x increase in habitat area you can expect a doubling in species number this relationship is best described by the regression formula S=cAz where: S = the number of species, c= a constant measuring the number of species/unit area, A= habitat area, and z is another constant measuring the shape of the line relating S & A

40
Often linearized ln (S ) and ln (A ) ln (S ) = ln (c ) + z ln (A )
z is now the slope ln (c ) is now the intercept ln (S ) ln (A )
 is now the intercept. ln (S ) ln (A )")
45
Island Biogeography (MacArthur and Wilson, 1960’s)
The number of species on an island is in a dynamic equilibrium determined by imm. and ext. rates immigration rate decreases with Sp. N since it becomes more likely that immigrants will not be new species extinction rate increases with Sp. N because of greater incidence of competitive exclusion equilibrium reached when immigration and extinction rates are equal equilibrium number is correlated with area and distance from mainland fig 53.26a

47
Area effect B A Mainland

48
Area Effect Island size influences immigration and extinction rates because…… larger islands are more likely to be found by immigrants which increases immigration rate organisms are less likely to go extinct on larger islands because there is more available habitat equilibrium number is higher on larger islands because of both higher immigration and lower extinction fig 53.26b

49
B Distance effect A Mainland

50
Distance Effect Distance from the mainland influences immigration and extinction rates given islands of the same size, immigration will be higher on near islands since they are more likely to be found by immigrants extinction rates the same (same size islands) equilibrium number is higher on near islands because of higher immigration fig 53.26c
 equilibrium number is higher on near islands because of higher immigration. fig 53.26c.")
51
Island biogeography is a simple model and we must also take into account abiotic disturbance, adaptive changes, and speciation events

52
Latitudinal species richness gradients
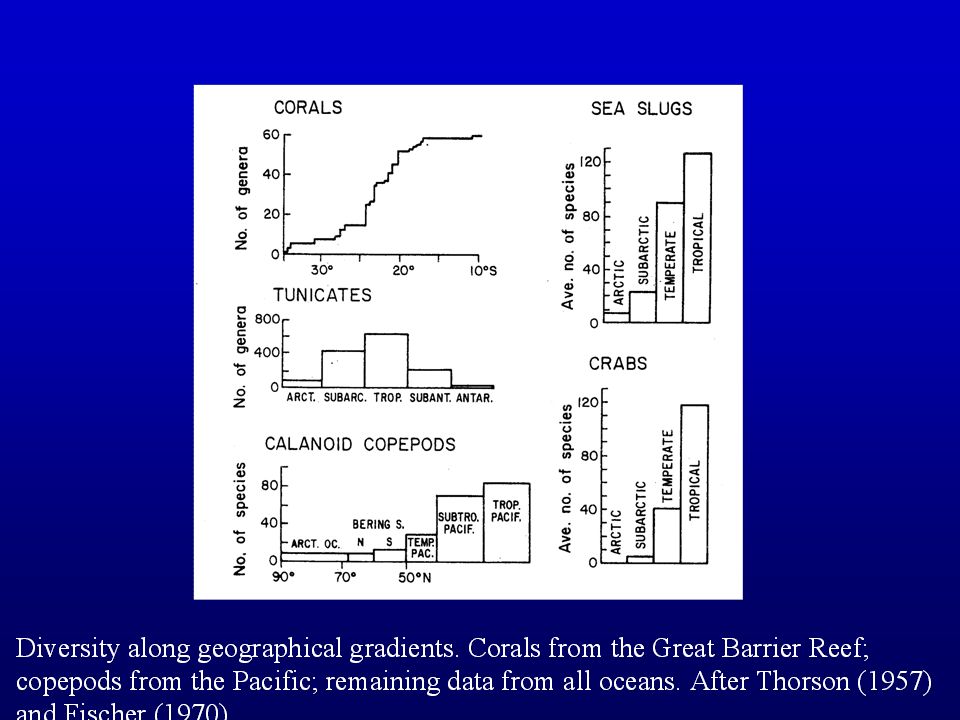
Species richness of many taxa declines from equator to poles Why? NOT CLEAR Land birds Could be evolutionary or ecological factors, or both? fig 53.23

54
Factors Proposed to Explain Latitudinal Diversity Gradients
History (more time permits more speciation Spatial Heterogeneity (more complex habitats provide more niches and permit more species to exist) Competition (competition favors reduced niche breadth, but competition can also eliminate species!) Predation (predation retards competitive exclusion)
 Competition (competition favors reduced niche breadth, but competition can also eliminate species!) Predation (predation retards competitive exclusion)")
55
Factors Proposed to Explain Latitudinal Diversity Gradients(2)
Climate (climatically favorable conditions allow more species to co-exist) Climate Stability (stable climates allow specialization to occur) Productivity (Diversity is limited by the amount of energy that can be partitioned) Disturbance (moderate disturbance retards competitive exclusion= intermediate disturbance hypothesis)
 Climate Stability (stable climates allow specialization to occur) Productivity (Diversity is limited by the amount of energy that can be partitioned) Disturbance (moderate disturbance retards competitive exclusion= intermediate disturbance hypothesis)")
60
Application of biogeographic principles to the design of nature preserves. In each pair of figures the design on the left is preferred over that on the right, even though both incorporate the same area. The concepts are: A, a continuous reserve is better than a fragmented one; B, the ratio of area to perimeter should be maximized; C, distance between refuges should be minimized; and D, dispersal corridors should be provided between fragments. (from Ecology and Evolution of Communities, ed. M. L. Cody and J. M. Diamond,

61
Ecosystems Ecology

62
Food Chains The energy flow from one trophic level to the other is the food chain A food chain involves one type of organism at each trophic level Producers (Autotrophs) Primary Consumers – eat producers Secondary Consumers – eat the primary consumers Tertiary Consumers – eat the secondary consumers Decomposers – bacteria and fungi that break down dead organisms and recycle materials
 Primary Consumers – eat producers. Secondary Consumers – eat the primary consumers. Tertiary Consumers – eat the secondary consumers. Decomposers – bacteria and fungi that break down dead organisms and recycle materials.")
63
What is a Food Web? Describes which organisms in communities eat other kinds of organisms Community food web is a description of feeding habits of a set of organisms based on taxonomy, location or other criteria Webs were derived from natural History approaches to describing community structure

64
What is a Food Web (2)? Food webs portray flows of matter and energy within the community Web omits some information about community properties e.g., minor energy flows, constraints on predation, population dynamics

65
Food Webs: Methods Identify component species
Sample to determine who is eating whom Sampling and gut analysis to quantify frequency of encounters Exclosures and removals of species to determine net effects Stable isotopes Mathematical models

66
Descriptive Food Webs

67
Interaction or functional food webs depict the most influential link or dynamic in the community

68
What is a Food Web (cont.): Complexity meets reality
Fallacy of linear food chains as a adequate description of natural food webs Food webs are reticulate Discrete homogeneous trophic levels an abstraction or an idealism omnivory is rampant ontogenetic diet shifts (sometimes called life history omnivory) environmental diet shifts spatial & temporal heterogeneity in diet
 environmental diet shifts. spatial & temporal heterogeneity in diet.")
69
Are trophic levels useful?
Even if organisms are not strict herbivores, primary carnivores, etc., as long as they are mostly feeding at one trophic level, the concept can have value (e.g., trophic cascade concept).
.")
70
What is a food web (cont.)?
Modern Approaches to Food Web Analysis Connectivity relationships Importance of predators and interaction strength in altering community composition and dynamics

71
Energy flow through ecosystems
Energy transfer between trophic levels is not 100% efficient, and energy is lost as it passes up a food chain. Herbivores eat a small proportion of total plant biomass; they also use only a small proportion of plant material consumed for their growth. The rest is lost in feces or respiration Thus, less energy is available for the next trophic levels

72
Trophic Basis of Production
Assimilation efficiency varies with resource 10% for vascular plant detritus 30% for diatoms and filamentous algae 50% for fungi 70% for animals 50% for microbes (bacteria and protozoans) 27% for amorphous detritus Net Production Efficiency production/assimilation ~ 40%
 27% for amorphous detritus. Net Production Efficiency. production/assimilation ~ 40%")
73
Marine Ecology: Food Webs
Ecological efficiency is defined as the energy supply available to trophic level N + 1, divided by the energy consumed by trophic level N. You might think of it as the efficiency of copepods at converting plants into fish food. In general, only about 10% of the energy consumed by one level is available to the next.=, but this can vary substantially. Difficult to measure so scientists focus on measures of assimilation efficiency for selected groups of animals.

74
Food Webs A pyramid of biomass represents the amount of energy, fixed in biomass, at different trophic levels for a given point in time The amount of energy available to any trophic level is limited by the amount stored by the level below. Because energy is lost in the transfer between levels, there is successively less total energy at higher trophic levels.

75
Food Webs in the Ocean The oceans can be an exception, because the total amount of biomass in algae is usually small. A pyramid of biomass for the oceans can appear inverted However, a pyramid of energy, which shows rates of production rather than biomass, must have the pyramid shape. Algae can double in days, while zooplankton might double in months, and fish might only reproduce once a year. Thus, a pyramid of energy takes into account turnover rate, and can never be inverted.

76
Decomposition and Mineralization
Most material is derived from plants Involves: Release of chemical energy Mineralization (= organic --> inorganic) Note immobilization = reverse of mineralization Net mineralization rate = mineralization - immobilization
 Note immobilization = reverse of mineralization. Net mineralization rate = mineralization - immobilization.")
77
Terrestrial communities: Nutrient sources
Weathering of rock (K, P, Ca and many others) Fixation of CO2 (photosynthesis) and N2 Dryfall (particles in the atmosphere) Wetfall (snow & rain); contains Oxides of S, N Aerosols particles high in Na, Mg, Cl, S produced by evaporation of droplets Dust particles from fires, volcanoes Ca, K, S
 Fixation of CO2 (photosynthesis) and N2. Dryfall (particles in the atmosphere) Wetfall (snow & rain); contains. Oxides of S, N. Aerosols. particles high in Na, Mg, Cl, S. produced by evaporation of droplets. Dust particles from fires, volcanoes. Ca, K, S.")
78
Terrestrial communities: Nutrient losses
Release to atmosphere CO2 from respiration Volatile hydrocarbons from leaves Aerosols NH3 (decomposition), N2 (denitrification) Loss in streamflow Dissolved nutrients Particles
, N2 (denitrification) Loss in streamflow. Dissolved nutrients. Particles.")
79
Oceans No outflow Detritus sinks --> mineralization --> nutrients end up Being carried back to surface in upwelling currents, o Trapped in bottom sediments (e.g., phosphorus: 1% lost to sediment with each cycling)
")
80
CARBON CYCLE 4 PROCESSES MOVE CARBON THROUGH ITS CYCLE: Biological
CO2 4 PROCESSES MOVE CARBON THROUGH ITS CYCLE: Biological Geochemical Mixed biochemical Human Activity CO2 80

81
Figure 22.5 The largest pool in rocks is not shown In oceans, CO2 is mostly as bicarbonate and carbonate ions There could be a net terrestrial sink…

82
NITROGEN CYCLE Nitrogen-containing nutrients include: Ammonia (NH3)
in Atmosphere Nitrogen-containing nutrients include: Ammonia (NH3) Nitrate (NO3-) Nitrite (NO2-) ORGANISMS NEED NITROGEN TO MAKE AMINO ACIDS FOR BUILDING PROTEINS!!! N03- & N02- NH3 82
 Nitrate (NO3-) Nitrite (NO2-) ORGANISMS NEED NITROGEN TO MAKE AMINO ACIDS FOR BUILDING PROTEINS!!! N03- & N02- NH")
83
The nitrogen cycle Figure 22.6
Nitrogen is generally available as either nitrate or ammonium N2 fixation in agricultural systems: rhizobium bacteria associated with approx 200 specis of leguminous plants Non-agricultural systems: 12,000 species plus cynaobacteria

84
PHOSPHORUS CYCLE PHOSPHORUS FORMS PART OF IMPORTANT LIFE-SUSTAINING MOLECULES (ex. DNA & RNA) 84
 84")
85
The phosphorus cycle Figure 22.9 No gaseous component
Natural scarcity (limiting nutrient in aquatic envrionments) Main reservoirs: rocks and minerals Realesed by weatering, erosion, runoff (fertilizers) In soils only a small fraction available to plants
 Main reservoirs: rocks and minerals. Realesed by weatering, erosion, runoff (fertilizers) In soils only a small fraction available to plants.")
86
We’re in the Driver’s Seat - Human Activities Dominate Many Biogeochemical Cycles

88
Disturbance simplified
The greater the disturbance the more habitat that will be opened up

90
Disturbance opens space; slate wiped clean
Time 0 Disturbance opens space; slate wiped clean Time 1 Only certain species can establish themselves in open space; Opportunists, Fugitives, Weeds No special requirements for first colonizers First colonists make environment less suitable for their own further recruitment, but this has little or no effect on other species Time 2 First colonists modify environment so it becomes less suitable for their further recruitment but more suitable for other species First colonists make environment less suitable for all subsequent species Time 3 Process continues until residents no longer facilitate recruitment of other species Process continues until no species can invade and grow in presence of residents First colonists continue to hold space and exclude all others (First Come, First Served) Model FACILITATION TOLERANCE INHIBITION
 Model. FACILITATION. TOLERANCE. INHIBITION.")
94
F.E. Clements (1916, 1936) idea of succession
Sere Climax Ecosystem = superorganism

95
Succession b A c B c C Early Colonizing Mid Mixed Late Climax Time

Similar presentations



























 – PREVALENT.>")



:>")
 a. What are 2 reasons described that plant.>")

