Download presentation
Presentation is loading. Please wait.

1
Module 632 Lecture 8 JCS Mechanism of Muscle contraction

2
MODULE - 632Lecture 8 Muscle Contraction Lecture outcomes: At the end of this lecture a student will be aware : 1) of the major sarcomeric proteins and their molecular properties 2) of the structure of the thick filament 3) of the structure of the thin filament 4) of how the major proteins can be purified/polymerised 5) of the biochemical techniques used to study the actomyosin motor 6) of the crossbridge and its cycle linked to ATP hydrolysis 7) of the evidence that myosin ‘rocks, swings or rotates’ when attached to actin. 8) that the crossbridge model can explain the macroscopic properties of contracting sarcomeres 9) that sarcomere length correlates with specialised muscle function – high-speed or high force.
 that the crossbridge model can explain the macroscopic properties of contracting sarcomeres 9) that sarcomere length correlates with specialised muscle function – high-speed or high force..")
3
IA M ZZ I AI Sarcomere – the contractile unit

4
Myosin containing, thick filament Actin containing, thin filaments acto-myosin “cross-bridges” Acto-myosin in striated muscle : Sarcomere 0.5 m 0.1 m 1 m

6
Sarcomeric proteins : Thick filament:Myosin – the major protein Titin Myosin-binding protein C M line proteins Thin filament:Actin – the major protein Tropomyosin Troponin complex – TnC, TnI, TnI Nebulin Z-disc:Actin -actinin and many other proteins

7
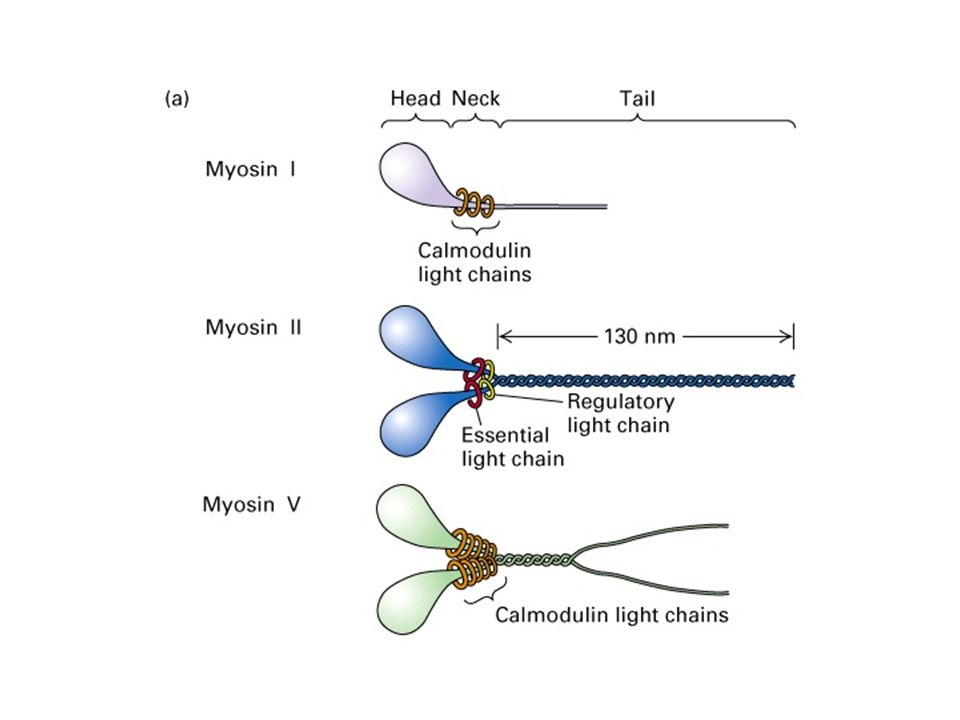
The Myosin Molecule

8
MYOSIN As with all your sarcomeric proteins it is insoluble: Myosin is extracted from muscle using high salt >500mM KCl Once extracted if you reduce the salt the myosin forms insoluble filaments. Why? The myosin molecules (dimers) form side-by-side associations through their rod regions – filaments are bipolar (as are native thick filaments) but have irregular length. To get soluble fragments for biochemistry purified myosin can be cut with proteases, under different conditions to form: Heavy meromyosin (HMM) – two heads joined through the S2 coiled coil – and the rod domain – light meromyosin (LMM) Under different cleavage condition one can release soluble single S1 ‘heads’, now usually referred to as the ‘motor domain’.
 form side-by-side associations through their rod regions – filaments are bipolar (as are native thick filaments) but have irregular length. To get soluble fragments for biochemistry purified myosin can be cut with proteases, under different conditions to form: Heavy meromyosin (HMM) – two heads joined through the S2 coiled coil – and the rod domain – light meromyosin (LMM) Under different cleavage condition one can release soluble single S1 ‘heads’, now usually referred to as the ‘motor domain’..")
9
MHC: (N to C terminal) Green – 27K domain; red – 50K domain (upper and lower); blue 20K domain and lever; ELC – yellow; RLC pink S1 HEAD Rayment et al., 1993
 Green – 27K domain; red – 50K domain (upper and lower); blue 20K domain and lever; ELC – yellow; RLC pink S1 HEAD Rayment et al., 1993")
11
TITIN: VSM contains titin (mol wt > 1 million). N-terminus embedded in Z-disc; runs along the thin filament in I band, but in I-band transfers to the thick (A) filament; coils around thick filament until its C-terminus; at C-term. has a kinase domain that lies within the M-line. Titin consists of serially repeated globular proteins domains with homology to domains in many other proteins: Ig – immunoglobulin domains (PEVK domains) Fn – fibronectin domains
 filament; coils around thick filament until its C-terminus; at C-term. has a kinase domain that lies within the M-line. Titin consists of serially repeated globular proteins domains with homology to domains in many other proteins: Ig – immunoglobulin domains (PEVK domains) Fn – fibronectin domains.")
12
What does titin do? 1) Titin is a probably a ‘ruler’? Determines layout of sarcomere and thick filament length. 2) Titin is elastic – may serve as a spring to centre thick filament array within the sarcomere. The ‘stretchiness’ (compliance) of titin has been studied by optical traps (see later) and by atomic force microscopes
 Titin is elastic – may serve as a spring to centre thick filament array within the sarcomere. The ‘stretchiness’ (compliance) of titin has been studied by optical traps (see later) and by atomic force microscopes.")
13
Myosin binding protein C – binds at one end to thick filament (titin + myosin) and at the other is proposed to attach to the actin: It seems to affect the level of the active state of myosin when activated. M-line proteins – cross-link thick filaments in the M- band; keeps the centered on each other and as a coordinated array. Z-line/discM-line

14
Sarcomeric proteins : Thin filament: F-actin core: The barbed end of the F-actin is embedded into the Z-disc – so each Z-disc contains overlapping F-actins from neighbouring half sarcomeres – which are cross-linked by -actinin (and other proteins) The pointed end of F-actin is capped with tropomodulin (CapZ)
 The pointed end of F-actin is capped with tropomodulin (CapZ)")
15
F-actin is formed by the polymerisation of actin protein monomers

16
Each 7 actin-unit (one-half turn) is associated with: Tropomyosin – -helical coiled-coiled dimer. End to end overlap continuous filament coiled around F- actin helix. Troponin complex – TnC, TnI and TnT – one complex is associated with each tropomyosin dimer Nebulin - A single (maybe 2) molecules of runs the length of each thin filament (controls the length of the thin filament?) - position of nebulin on outside of F-actin core is still not known.
 molecules of runs the length of each thin filament (controls the length of the thin filament ) - position of nebulin on outside of F-actin core is still not known..")
17
Actin Tropomyosin 7-actin repeat structure (14 - F-actin helix is double) in thin filament Structure repeats (half-turn) every 36.5nm
 in thin filament Structure repeats (half-turn) every 36.5nm")
18
IMPORTANT QUESTIONS TO UNDERSTAND HOW MUSCLE CONTRACTS: What is the evidence that crossbridges (myosin bound to actin) are independent force generators? How are the forces/movements generated ? How do the biochemical cycle and the mechanical events correlate?

19
Sliding Filament theory: HE Huxley – electron micrographs (1953) AF Huxley – interference light microscopy (1954)
 AF Huxley – interference light microscopy (1954)")
20
Cross-bridges are independent force generators: AF Huxley, Gordon & Julian (1966)
")
21
Swinging Cross-bridge hypothesis:HE Huxley, 1969 Micrographs and X- ray diffraction of insect flight muscle Reedy, Holmes and Tregear (1966)
")
22
Conclusions from basic fibre experiments: Sarcomeres contain inter-digitating thick and thin filaments: Shortening produces force Shortening of sarcomeres involves the relative sliding of thick and thin filaments Shortening requires – ATP The forces produced are correlated with the overlap of the thick and thin filaments The crossbridges show changes in shape/orientation depending on the nucleotide state – rigor, relaxed, active (disordered)
")
23
Filament sliding causes muscle to shorten myofibril sarcomere Light micrograph Electron micrograph Myosin molecules (purple bars) move over the F-actin (turquoise). This movement is powered by ATP.

24
Biochemical/Biochemical analysis of the crossbridge: Involved many experiments (they still continue) on: a)‘skinned’ fibres (under different conditions of nucleotide, phosphate, tension, length etc.) b)kinetic measurements of isolated proteins – myosin (incl. HMM and S1) and actin - using different biochemical and biophysical assays.
 and actin - using different biochemical and biophysical assays..")
25
Stopped Flow and Quench Flow

26
[P i ] Time Linked assay Quenched flow Time course of free and bound phosphate production when a molar excess of ATP is mixed rapidly with S1 M + ATP M.ATP M.ADP.P M.ADP + P
![[P i ] Time Linked assay Quenched flow Time course of free and bound phosphate production when a molar excess of ATP is mixed rapidly with S1 M + ATP M.ATP M.ADP.P M.ADP + P](http://images.slideplayer.com/14/4452375/slides/slide_26.jpg "[P i ] Time Linked assay Quenched flow Time course of free and bound phosphate production when a molar excess of ATP is mixed rapidly with S1 M + ATP M.ATP M.ADP.P M.ADP + P")
27
Results from biochemical experiments: ATP binding to myosin - very fast; diffusion controlled (10 -6 M -1.s -1 ) Acto-myosin dissociates very rapidly upon binding ATP Hydrolysis of ATP is fast (100 s -1 ) – release of P i is slow (<0.001 s -1 ) Release of Pi is accelerated by presence of F-actin Actomyosin ATPase catalytic site activity (turnover) is about 2s -1
 Acto-myosin dissociates very rapidly upon binding ATP Hydrolysis of ATP is fast (100 s -1 ) – release of P i is slow (<0.001 s -1 ) Release of Pi is accelerated by presence of F-actin Actomyosin ATPase catalytic site activity (turnover) is about 2s -1")
28
Biochemical pathway: Detached Attached ATP Pi ADP Detached Attached Duty ratio: r = (timeattached) / (cycle time) In this example = 75%
 / (cycle time) In this example = 75%")
29
The basic contractile mechanism is highly conserved BUT, you should remember that the rate constants of different biochemical steps can be very different between muscle and myosin types. e.g. Skeletal muscle myosins are much faster (every biochemical step is faster) than smooth muscle myosins Molluscan catch muscle has v.v. slow rate of ADP release
 than smooth muscle myosins Molluscan catch muscle has v.v. slow rate of ADP release.")
30
In vitro motility - assay HMM or S1 F-actin F-actin stabilised and fluorescently labelled with rhodamine-phalloidin

31
In vitro motility of F-actin (white filaments) moving on a surface coated with myosin (unseen) in the presence of ATP.
 moving on a surface coated with myosin (unseen) in the presence of ATP.")
32
Flipping between two EM reconstructions of F-actin decorated with smooth muscle myosin -with and without the presence of ATP Corresponds to transition of A.M.ADP to A.M biochemical states. Only a 20 o change.

33
Molecular model of acto- myosin to show ‘lever arm’ movement. Model is generated from: crystal structure of G-actin modelled into F-actin crystal structures of the S1 head high resolution EM pictures of S1- decorated F-actin atomic (crystal) structures ‘built’ into the ‘mass envelope’ of EM reconstruction of S1+F-actin
 structures ‘built’ into the ‘mass envelope’ of EM reconstruction of S1+F-actin.")
34
60’ 5->10 nm Myosin Subfragment-1 From Rayment et al. (1993) Science 261, 50-58
 Science 261, 50-58")
35
ATP ATP binds cleft opens Loss of ADP. Rigor ATP pocket closes, hydrolysis occurs Myosin detaches from actin Myosin starts to attach to actin Weak binding Strong binding Loss of Pi start of power stroke. The chemo-mechanical cycle :

36
Cartoon of myosin motor function – a rower

37
Effects of sarcomere architecture: Shorter sarcomeres give greater contraction speeds Longer sarcomeres produce greater forces

Similar presentations



















 The Muscle Action Potential ( AP ) Muscle RMP = -90 mV ( same as in nerves.>")
