Download presentation
Presentation is loading. Please wait.

1
Innate Defenses Complement The complement system is a biochemical cascade that helps clear pathogens from an organism It is part of the innate immune system The complement system consists of a number of small proteins found in the blood, normally circulating as inactive zymogens When these proteins are stimulated by one of several triggers, proteases in the system cleave specific proteins to release cytokines and initiate an amplifying cascade of further cleavages The end-result of this activation cascade is massive amplification of the response and activation of the cell- killing membrane attack complex

2
Innate Defenses Complement Proteins Over 20 proteins and protein fragments make up the complement system The complement proteins account for about 5% of the globulin fraction of blood serum The proteins and glycoproteins that constitute the complement system are synthesized by the liver hepatocytes. But significant amounts are also produced by tissue macrophages, blood monocytes and epithelial cells of genitourinal tract and gastrointestinal tract

3
Innate Defenses Complement Classical pathway Three biochemical pathways activate the complement system: the classical complement pathway, the alternative complement pathway, and the mannose-binding lectin pathway Classical complement pathway typically requires antibodies for activation (natural antibodies & specific immune response), whereas the alternative and mannose-binding lectin pathways can be activated by C3 hydrolysis or antigens without the presence of antibodies (non-specific immune response).
, whereas the alternative and mannose-binding lectin pathways can be activated by C3 hydrolysis or antigens without the presence of antibodies (non-specific immune response).")
4
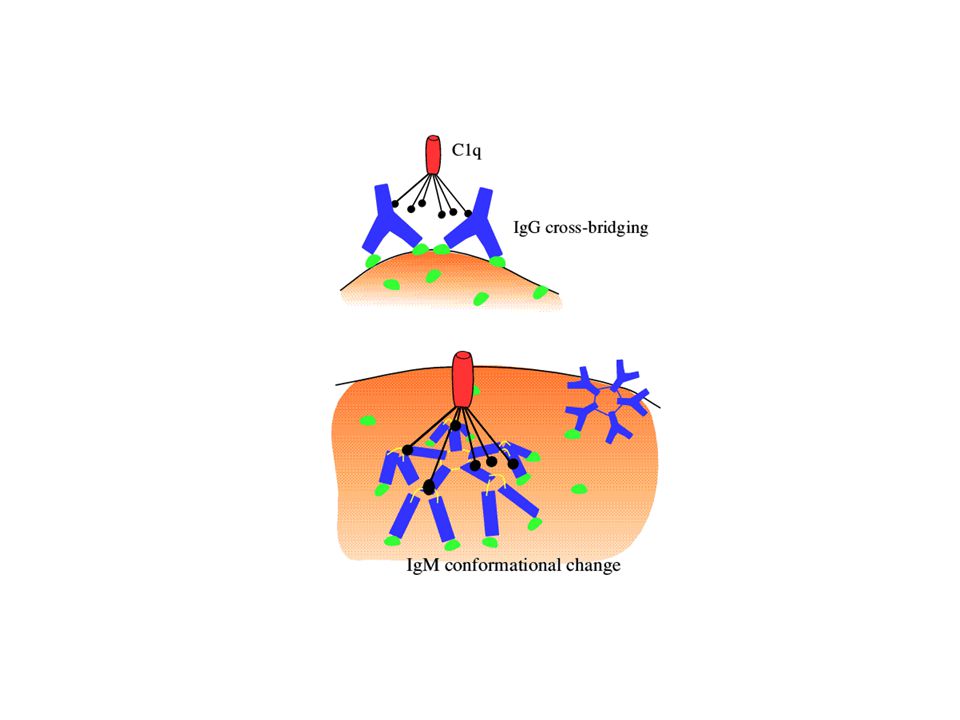
Innate Defenses Complement Classical pathway Classical pathway: is triggered by activation of the C1- complex (C1q, two molecules of C1r, and two molecules of C1s thus forming C1qr2s2) Activation of the C1-complex occurs when C1q binds to IgM or IgG complexed with antigens or when C1q binds directly to the surface of the pathogen A single IgM ('Natural' antibody or other IgM) can initiate the pathway, while multiple IgGs are needed) Such bindings lead to conformational changes in the C1q molecule, which leads to the activation of two C1r (a serine protease) molecules
 Activation of the C1-complex occurs when C1q binds to IgM or IgG complexed with antigens or when C1q binds directly to the surface of the pathogen A single IgM ( Natural antibody or other IgM) can initiate the pathway, while multiple IgGs are needed) Such bindings lead to conformational changes in the C1q molecule, which leads to the activation of two C1r (a serine protease) molecules")
6
Innate Defenses Complement Classical pathway They then cleave C1s (another serine protease) The C1r2s2 component now splits C4 and then C2, producing C4a,C4b,C2a,and C2b C4b and C2a bind to form the classical pathway C3-convertase (C4bC2a complex) C3-convertase (C4b2a complex) promotes cleavage of C3 into C3a and C3b C3b later joins with C4b2a (the C3 convertase) to make C5 convertase (C4bC2aC3b complex) The inhibition of C1r and C1s is controlled by C1-inhibitor C3-convertase can be inhibited by Decay accelerating factor (DAF)
 The C1r2s2 component now splits C4 and then C2, producing C4a,C4b,C2a,and C2b C4b and C2a bind to form the classical pathway C3-convertase (C4bC2a complex) C3-convertase (C4b2a complex) promotes cleavage of C3 into C3a and C3b C3b later joins with C4b2a (the C3 convertase) to make C5 convertase (C4bC2aC3b complex) The inhibition of C1r and C1s is controlled by C1-inhibitor C3-convertase can be inhibited by Decay accelerating factor (DAF)")
8
Innate Defenses Complement Alternative pathway It does not rely on a pathogen-binding antibodies like the other pathways Alternative complement pathway: is triggered by spontaneous C3 hydrolysis directly due to the breakdown of the thioester bond via condensation reaction (C3 is mildly unstable in aqueous environment) to form C3a and C3b C3b is then capable of covalently binding to a pathogenic membrane surface if it is near enough If there is no pathogen in the blood, the C3a and C3b protein fragments will be deactivated by rejoining with each other
 to form C3a and C3b C3b is then capable of covalently binding to a pathogenic membrane surface if it is near enough If there is no pathogen in the blood, the C3a and C3b protein fragments will be deactivated by rejoining with each other")
9
Innate Defenses Complement Alternative pathway Upon binding with a cellular membrane, C3b is bound by factor B to form C3bB This complex in presence of factor D will be cleaved into Ba and Bb Bb will remain covalently bonded to C3b to form C3bBb which is the alternative pathway C3-convertase. The C3bBb complex, which is "hooked" onto the surface of the pathogen, will then act like a "chain saw," catalyzing the hydrolysis of C3 in the blood into C3a and C3b This positively affects the number of C3bBb hooked onto a pathogen

10
Innate Defenses Complement Alternative pathway After hydrolysis of C3, C3b complexes to become C3bBb, which cleaves C5 into C5a and C5b C5b with C6, C7, C8, and C9 (C5b6789) complex to form the membrane attack complex The membrane attack complex, also known as MAC, which is inserted into the cell membrane, "punches a hole," and initiates cells lysis. C5a and C3a are known to trigger mast cell degranulation IgA is associated with activating the alternative path

11
Innate Defenses Complement Lectin pathway Lectin pathway (MBL - MASP): Mannose-binding lectin pathway The lectin pathway is homologous to the classical pathway, but with the opsonin, mannose-binding lectin (MBL), and ficolins, instead of C1q Ficolins are homologous to MBL and function via MASP in a similar way. In vertebrates without an adaptive immune system, ficolins are expanded and their binding specificities diversified to compensate for the lack of pathogen-specific recognition molecules. This pathway is activated by binding mannose-binding lectin to mannose residues on the pathogen surface

12
Innate Defenses Complement Lectin pathway This activates the MBL-associated serine proteases, MASP-1, and MASP-2 (very similar to C1r and C1s, respectively),which can then split C4 into C4a and C4b and C2 into C2a and C2b C4b and C2a then bind together to form the C3-convertase, as in the classical pathway
,which can then split C4 into C4a and C4b and C2 into C2a and C2b C4b and C2a then bind together to form the C3-convertase, as in the classical pathway")
13
Innate Defenses Complement Complement also serves to activate the acute inflammatory response In all three pathways, a C3-convertase cleaves and activates component C3, creating C3a and C3b and causing a cascade of further cleavage and activation events C5b initiates the membrane attack pathway, which results in the membrane attack complex (MAC), consisting of C5b, C6, C7, C8, and polymeric C9 MAC is the cytolytic endproduct of the complement cascade; it forms a transmembrane channel, which causes osmotic lysis of the target cell. C3b binds to the surface of pathogens leading to greater internalization by phagocytic cells by opsonization

14
Innate Defenses Complement C5a is an important chemotactic protein, helping recruit inflammatory cells such as neutrophils and activates vascular endothelium directly C5a also activates mast cells which amplify the inflammatory signals by releasing their preformed vasoactive mediators. Both C3a and C5a have anaphylatoxin activity, directly triggering degranulation of mast cells as well as increasing vascular permeability and smooth muscle contraction Kupffer cells and other macrophage cell types help clear complement-coated pathogens.

Similar presentations



 Activation pathways Effector functions Regulation of complement activation.>")
BB Fall 2007.>")






BB Unit 1 Nature of the Immune System Part 9 Complement.>")

 It has made the self/nonself discrimination on an evolutionary time-scale It uses.>")
 Harvianto Siman Santosa ( )>")

 MLIM-101 Prepared by: Dr. Mohamed S. Abdel-Latif.>")



 Complement Complement refers, historically, to fresh serum capable of lysing antibody (Ab)-coated cells. This.>")