Download presentation
Presentation is loading. Please wait.

1
Chapter 4-2 Transpiration – diffusion of water vapor

2
Transpiration – diffusion of water vapor Diffusion is primary means of any further movement of the water out of the leaf. That is water movement is controlled by the concentration gradient of water vapor. Two factors determine transpiration : Difference in water vapor concentration (Cwv) Resistance of the pathway for vapor diffusion (Diffusional resistance) Leaf stomatal resistance ( r s) Boundary layer resistance ( r b)
 Resistance of the pathway for vapor diffusion (Diffusional resistance) Leaf stomatal resistance ( r s) Boundary layer resistance ( r b).")
3
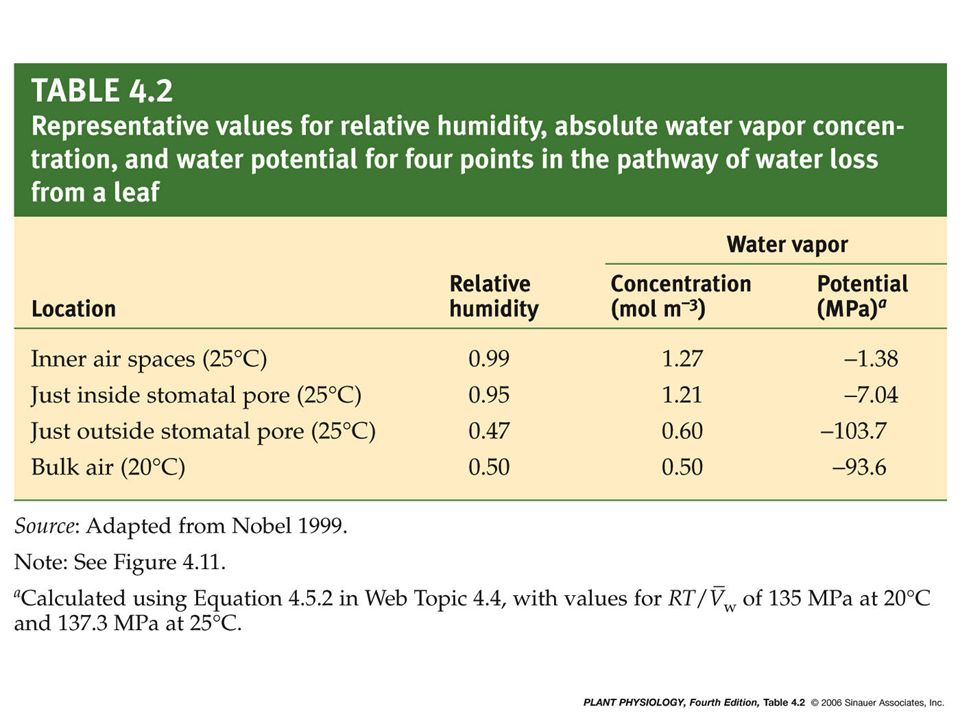
E = Cwv (leaf) – Cwv (air) r s + r b E : transpiration rate (mol/m 2 /s) r s : resistance at the stomatal pore (s/m) (leaf stomatal resistance) r b : resistance due to the boundary layer boundary layer : the layer of unstirred air at the leaf surface (boundary layer resistance) However, it is difficult to measure the Cwv (leaf). Sometimes vapor pressure are used instead of concentrations, and the difference is called “water vapor pressure deficit”. Water vapor pressure deficit = Pwv (leaf) - Pwv (air)
 - Pwv (air).")
5
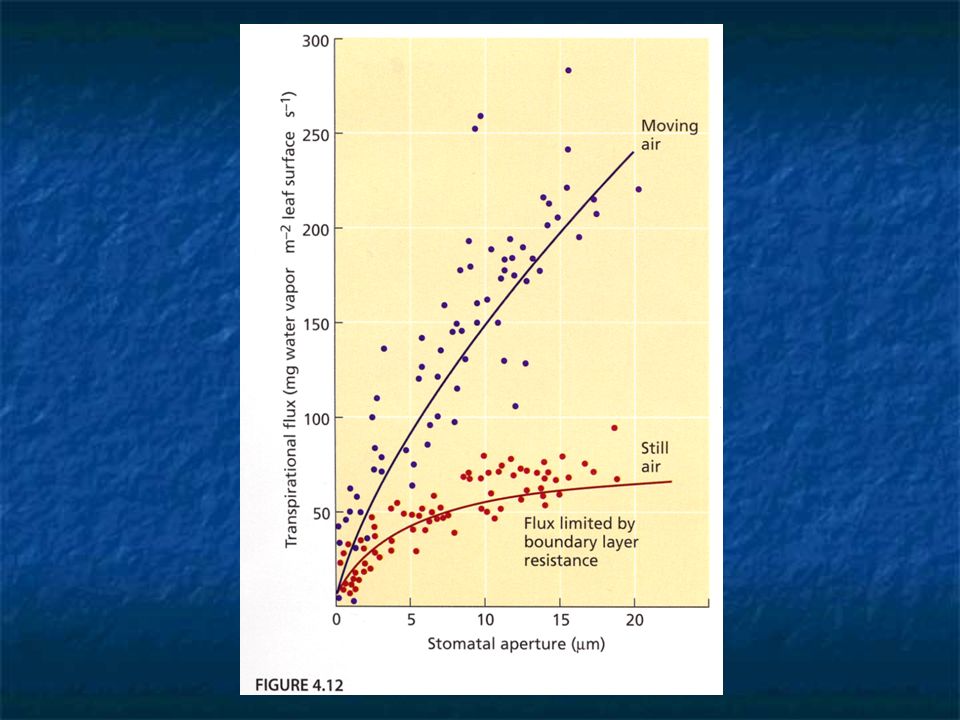
Air boundary layer A thin film of still air on the surface of leaf and its resistance to water vapor diffusion is proportional to its thickness. The thickness of the boundary layer is determined primarily by wind speed.

7
When air surrounding the leaf is still, increases in stomatal aperture have little effect on transpiration rate. The thickness of the boundary layer is the primary deterrent to water vapor loss from the leaf. When wind velocity is high, the stomatal resistance has the largest amount of control over water loss.

8
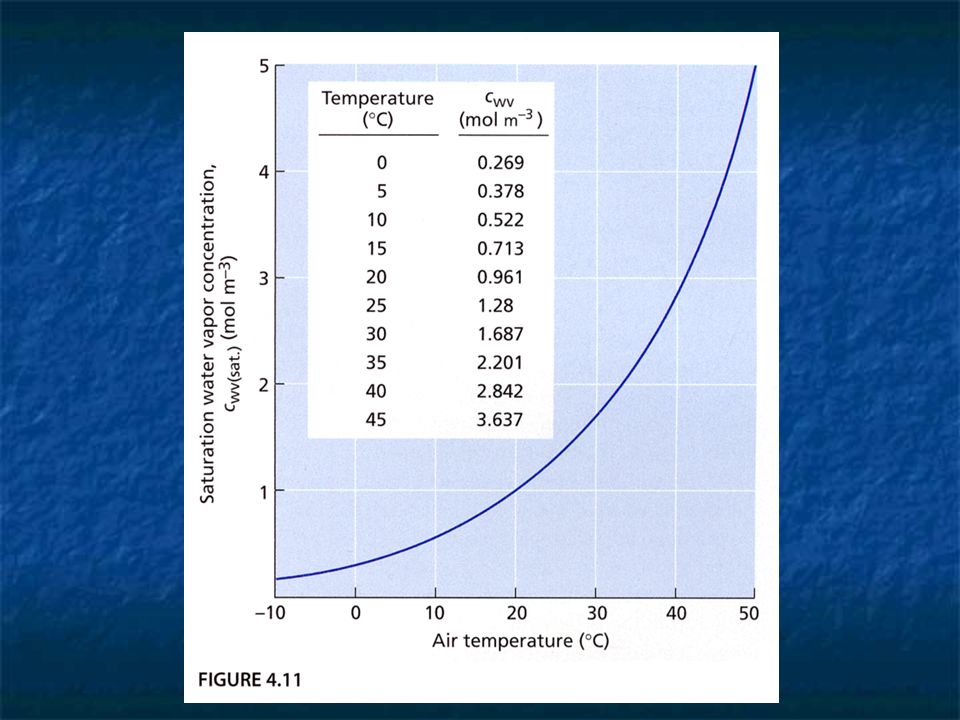
Relative humidity (RH) RH = Cwv / Cwv (sat.) The Cwv (sat.) is strongly dependent on temp.
 RH = Cwv / Cwv (sat.) The Cwv (sat.) is strongly dependent on temp.")
11
Water loss Water loss is regulated by : Difference in water vapor concentration The pathway resistance

12
The concentration gradient for CO 2 uptake is smaller than the concentration gradient driving water loss. How can plant prevent water loss without simultaneously excluding CO 2 uptake? Temporal regulation – When water is abundant : At night, no photosynthesis Stomata close, preventing unnecessary loss of water. Sunny morning, photosynthesis is demanding, supply of water is abundant. Stomata are wide open, decreasing the stomatal resistance to CO 2 diffusion. When soil water is less abundant The stomata will open less or even remain closed on a sunny morning.

13
Transpiration ratio A measure of the relationship between water loss and carbon gain Transpiration ratio = moles of H 2 O transpired moles of CO 2 fixed The factors that cause large ratio of H 2 O efflux to CO 2 influx : The concentration gradient driving water loss is about 50 times larger than driving CO 2 influx. CO 2 diffuse slower than water does. MW of CO 2 > MW of H 2 O CO 2 has longer diffusion path membrane -> cytoplasm -> chloroplast envelop -> chloroplast

14
In dicot and non grass monocot Usually no subsidiary cell Kidney-shaped guard cells Grass stomatal complex pore Stomata

15
Oat 35 34

16
Ventral wall (thicker ~ 5 m) dorsal wall (thinner)
 dorsal wall (thinner)")
17
Ion and organic molecules Guard cell s Decrease ( s = -RTCs) w decrease Water moves into the guard cell p increase Stoma open
 w decrease Water moves into the guard cell p increase Stoma open")
19
The mechanism of guard cell movement

20
H 2 O CO 2 Light energy The three principal raw materials for photosynthesis

21
Monocot leaf does not have palisade and spongy mesophyll cells. Palisade cells generally have larger numbers of chloroplasts than spongy mesophyll cells. The conc. of chlorophyll is also higher in palisade cells than in spongy cell (1.5 ~ 2.5 times higher). Sieve effect : Since there is a significant proportion of the cell volume that does not contain chloroplast, a substantial amount of light may pass through the first cell without being absorbed.
. Sieve effect : Since there is a significant proportion of the cell volume that does not contain chloroplast, a substantial amount of light may pass through the first cell without being absorbed..")
23
The major role of stomata is to allow entry of CO 2 into leaf for photosynthesis while preventing excessive loss of water. However, plant can not uptake CO 2 and simultaneously preventing water loss. Stomatal complex (stomatal apparatus) Stoma Guard cells Subsidiary cells
 Stoma Guard cells Subsidiary cells.")
24
Elliptic type Graminaceous type Kidney-shaped Dumbbell-shaped

25
The rate of CO 2 diffusion is proportional to the diameter of the pores, not the area. That is, as the pore size decrease, the diffusion rate ( g CO 2 /hr) of CO 2 decrease proportionally. However, as the pore size decrease, the efficiency of CO 2 diffusion per unit area increases several fold.
 of CO 2 decrease proportionally. However, as the pore size decrease, the efficiency of CO 2 diffusion per unit area increases several fold..")
27
Are the stomata on the upper or lower leaves?

28
Oat 35 34

30
Ventral wall (thicker ~ 5 m) dorsal wall (thinner)
 dorsal wall (thinner)")
31
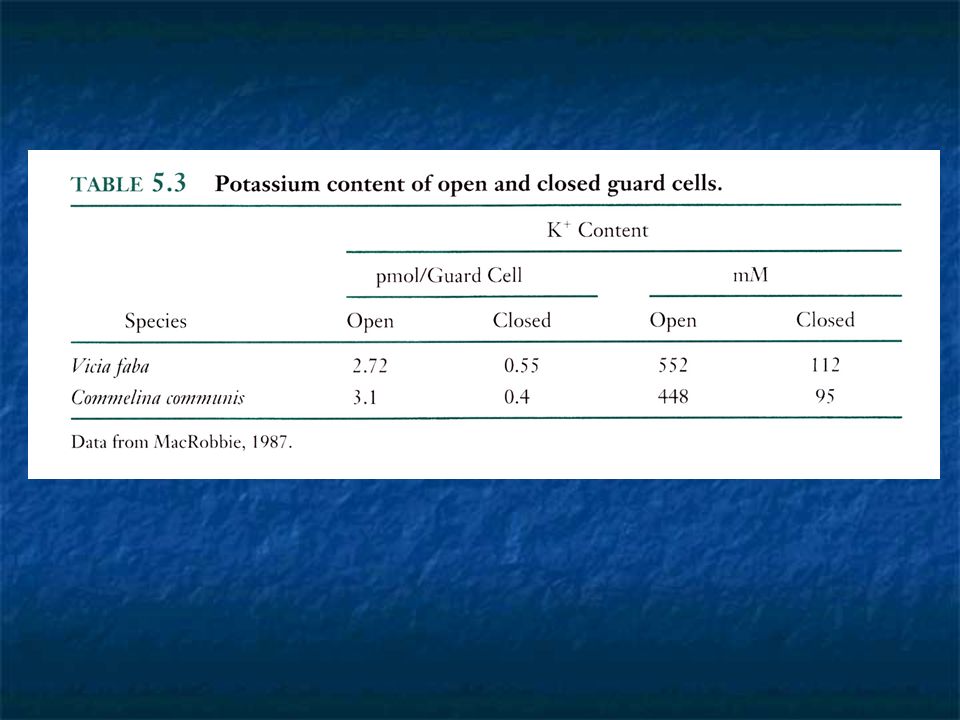
The mechanism of guard cell movement Driving force -> osmotic uptake of water -> increase of hydrostatic pressure -> push the thin dorsal walls outward into the neighboring epidermal cells. -> cause cell to arch. In dumbbell-shape guard cells, the bulbous ends of the cells push against each other as they swell, driving the central handles apart in parallel and widening the pore. In 1960, it became evident that K + levels are high in open guard cells and very low in closed guard cells. Upon opening, large amounts of K + move from the subsidiary and epidermal cells into the guard cells.

32
Ion and organic molecules Guard cell s Decrease ( s = -RTCs) w decrease Water moves into the guard cell p increase Stoma open
 w decrease Water moves into the guard cell p increase Stoma open")
34
Mechanism Proton extrusion Apply fusicoccin -> stimulate H + extrusion -> stimulate stomatal opening Vanadate (VO 3 -, 釩酸鹽 ) -> inhibit proton pump -> inhibit stomatal opening Establish a pH gradient across membrane Influx of K + by passive uptake through K + channels
 -> inhibit proton pump -> inhibit stomatal opening Establish a pH gradient across membrane Influx of K + by passive uptake through K + channels")
35
In most plants, malate is used as counterions to balance K + uptake. In plants whose guard cells lack chloroplast or starch, Cl - is also used as counterions. The evidence for malate as a counterion Malate conc. is high in opened stomata. PEP carboxylase which catalyzed the formation of malate is high in opened stomata. The starch conc. is decreasing in opened stomata that correlates with the increasing amount of malate.

36
ADP + Pi Osmotic potential decrease Water potential decrease Water enter into vacuole Increase turgor Stomata open

37
Mechanism for stomatal close Uptake of Ca +2 into the cytosol Depolarize the membranes Anion channel opened and Cl - and malate released from the vacuole. K + channel opened and K + released from vacuole and subsequently into subsidiary cells.

38
ADP + Pi Osmotic potential increase Water potential increase Water comes out from vacuole decrease turgor Stomata close Ca +2

39
Control of stomatal movement CO 2 level Light Water stress Temperature Circadian rhythms

40
Effect of CO 2 on stomatal movement CO 2 level CO 2 conc. decrease => stomata open => to uptake more CO 2 CO 2 conc. increase => stomata close The response of the stomata is to the intracellular conc. of CO 2 in the guard cells. When CO 2 level decrease or photosynthesis is needed, guard cells will take up water and swell to open the pores, in order to take more CO 2. When CO 2 level increase or the water stress override the photosynthesis, the guard cells will close.

41
Effect of light on stomatal movement Light Light (blue and red light) => stomata open (to admit CO 2 for photosynthesis) Stomata closed by exposure to high CO 2 can be induced to open slowly if placed in the light. CO 2 conc. increase -> stomata close -> under light -> stomata open Stomata normally open at dawn.

42
Dual-beam experiment The pulse of blue light stimulates a significant increase in stomatal conductance (increase stomatal opening).
.")
43
Indirect effect High fluence rate => CO 2 conc. in intercellular space is decreasing due to photosynthesis in mesophyll cells. Light affects stomatal movement through regulation of intercellular CO 2 levels. Closure of the guard cells in the dark can be attributed to the accumulation of respiratory CO 2 inside the leaf. Direct effect Effect of light on stomatal movement

44
Indirect effect Direct effect Blue light has direct effect on stomatal opening. At low fluence rate, blue light causes stomatal opening, but not red light. At high fluence rate, stomatal opening under blue light is consistently higher than under red light. Blue light -> blue/UV-A crytochrome -> proton extrusion -> photosynthesis -> ATP production -> stomatal opening Why ? Light on signal sunflecks Effect of light on stomatal movement

45
Effect of water on stomatal movement Hydropassive closure Water loss from the guard cells exceeds the rate of movement into the guard cells. => decrease in turgidity of the guard cells => stomata close Hydroactive closure ABA (abscisic acid, 離層酸 ) referred to as an antitranspirant. Plants sense water deficit => ABA is released into the apoplast and then transferred to the guard cells => stomata close => ABA conc. is increased => more stomata close ABA originally accumulate in the chloroplast -> water stress -> pH in chloroplast decrease and the pH in cytoplasm and apoplast increase -> ABA release into apoplast.
 referred to as an antitranspirant. Plants sense water deficit => ABA is released into the apoplast and then transferred to the guard cells => stomata close => ABA conc. is increased => more stomata close ABA originally accumulate in the chloroplast -> water stress -> pH in chloroplast decrease and the pH in cytoplasm and apoplast increase -> ABA release into apoplast..")
46
Effect of temperature on stomatal movement Temp increase -> stimulate respiration and impair photosynthesis -> CO 2 conc. increase -> stomata close Midday closure temp increase -> photosynthesis reduced water deficit occur -> photosynthesis reduced

47
Effect of circadian rhythms on stomatal movement An endogenous circadian rhythm control stomatal opening, but it is still unclear how the rhythm control stomatal movement.

48
END

Similar presentations









. What is the job of air spaces?>")



, Chapter 23 (p. 603-609)>")


, 23>")
, 18 (p.449-455)>")






