Download presentation
Presentation is loading. Please wait.

1
Sistema uditivo e vestibolare

2
Il suono è un’onda elastica (ha bisogno di un mezzo per propagarsi), longitudinale (la perturbazione avviene parallelamente alla direzione di propagazione); per la sua esistenza sono, dunque, necessari una sorgente (corpo vibrante) e un mezzo elastico di propagazione (aria, acqua, ecc..). In un mezzo gassoso le onde si possono propagare anche abbastanza lontano.

3
Il suono è, quindi, un modo di trasmissione di energia meccanica che, irradiandosi dalla sorgente attraverso il mezzo di propagazione, arriva ai corpi riceventi. La perturbazione che viaggia nel mezzo consiste, fisicamente, in un susseguirsi di pressioni e depressioni e, quindi, in un’oscillazione di ogni particella in vibrazione attorno ad una sua posizione media, senza spostamento di materia

4
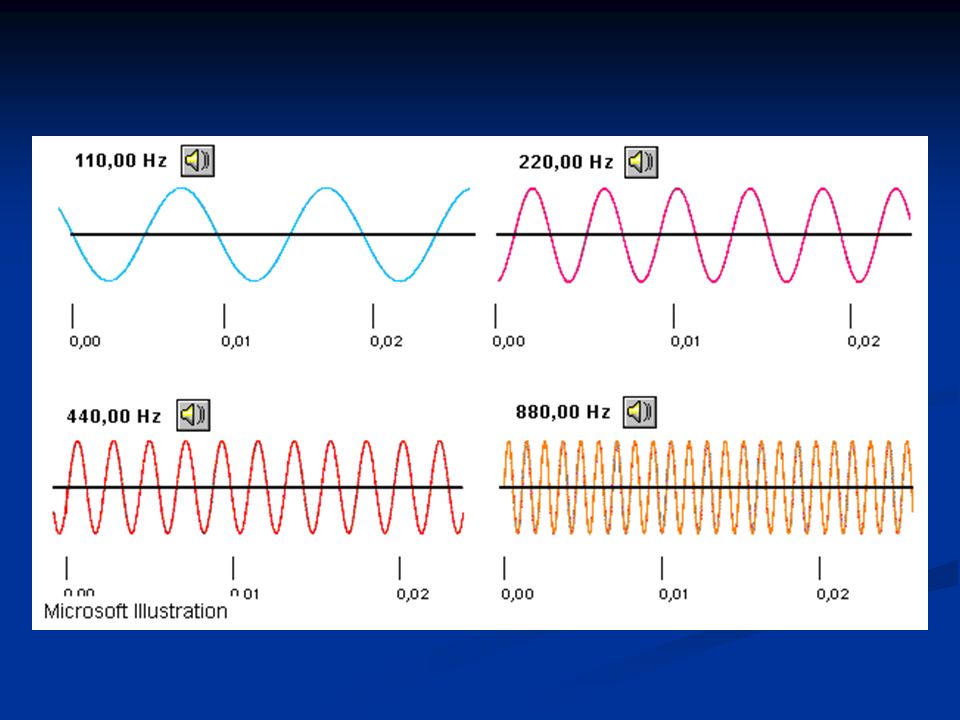
Vediamo quali sono le grandezze fisiche che caratterizzano un suono. Frequenza (ν) Frequenza (ν) E’ il numero di vibrazioni complete che avvengono in un secondo. Si misura in hertz ( Hz ). Il "range" di udibilità dell’orecchio umano è compreso tra i 20 e i 20000 Hz. Ciò significa che, pur esistendo onde sonore che si propagano a frequenze più basse (infrasuoni) o più alte (ultrasuoni), noi non possiamo percepirle.
 Frequenza (ν) E’ il numero di vibrazioni complete che avvengono in un secondo. Si misura in hertz ( Hz ). Il range di udibilità dell’orecchio umano è compreso tra i 20 e i Hz. Ciò significa che, pur esistendo onde sonore che si propagano a frequenze più basse (infrasuoni) o più alte (ultrasuoni), noi non possiamo percepirle..")
6
Lunghezza d’onda (λ) E’ la distanza percorsa dall’onda in un periodo. Ampiezza L’ ampiezza dell’onda rappresenta lo spostamento massimo delle molecole che oscillano intorno alla posizione di equilibrio al passaggio della perturbazione acustica. All’aumentare di questo spostamento aumenta la forza con cui le molecole colpiscono la membrana timpanica e, quindi, l’intensità del suono che percepiamo. L’energia delle vibrazioni varia in un ambito di 10 12 volte, pertanto si preferisce usare il Log

7
Con W= energia del suono in esame e W 0 = energia di riferimento Ambito dinamico: 0-120 dB

8
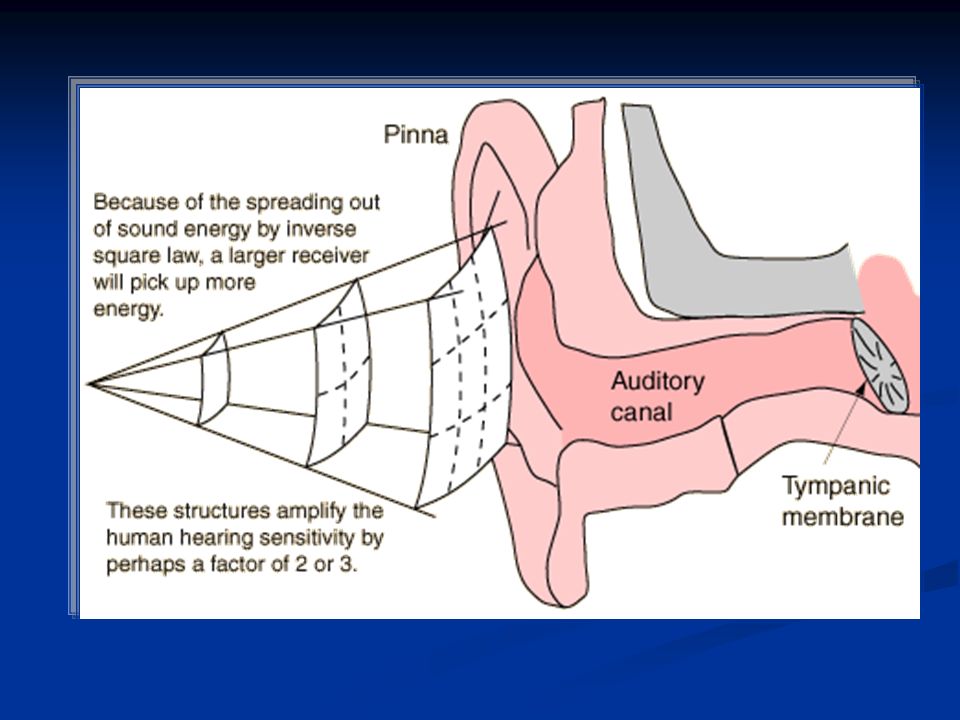
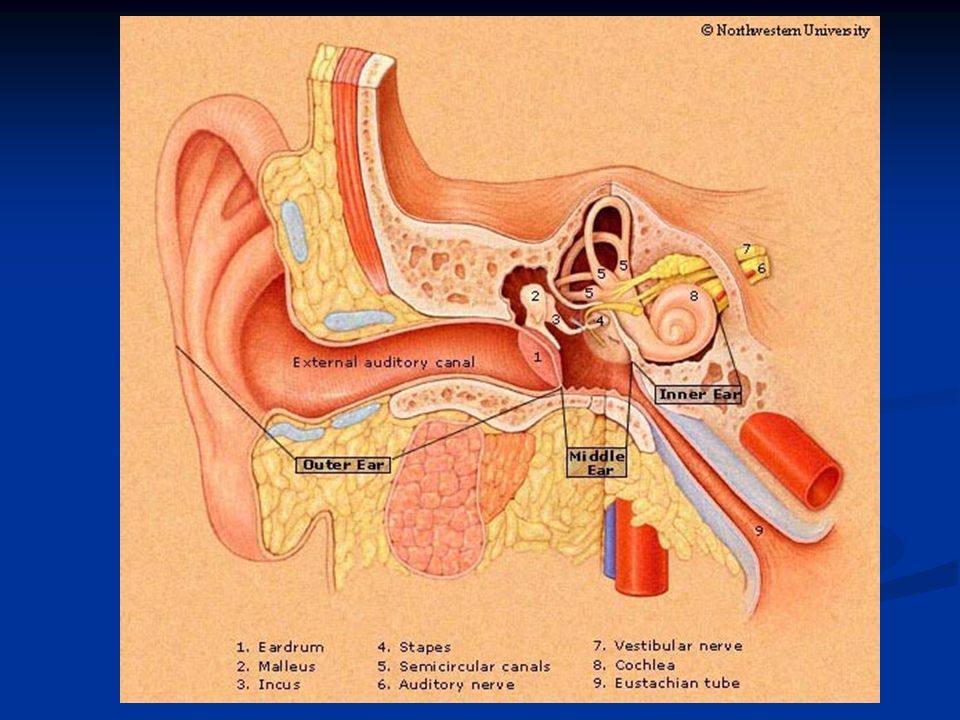
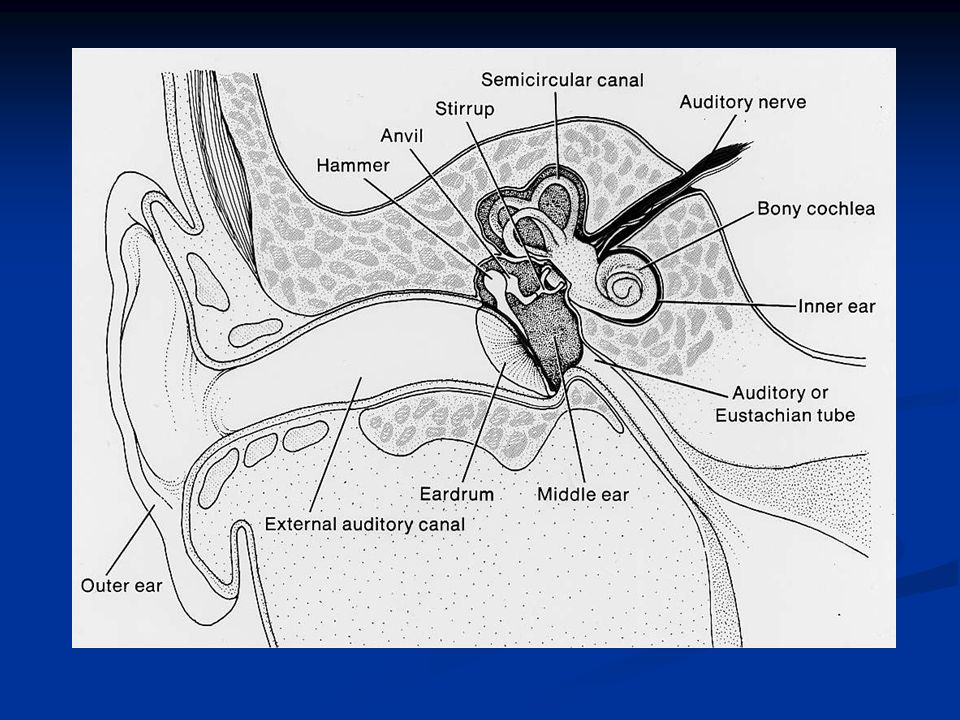
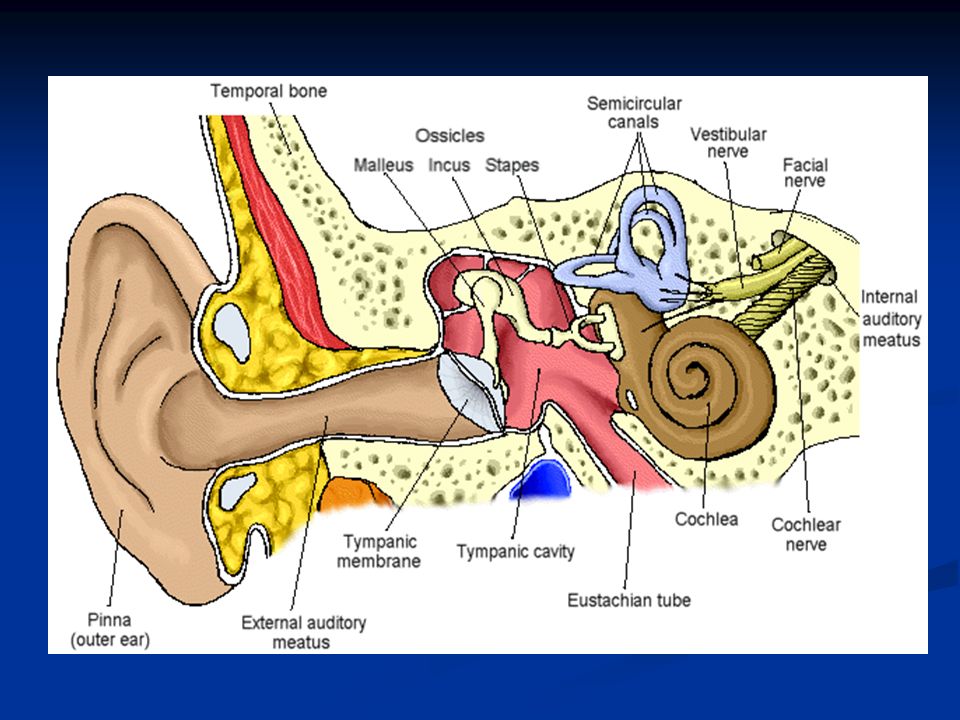
L’organo dell’udito è l’orecchio che si divide in: Oreccchio esterno (attraverso l’osso temporale): padiglione auricolare e meato uditivo esterno Orecchio medio (regione mastoidea del lobo temporale): tromba di Eustachio, catena degli ossicini (martello, incudine e staffa), muscolo stapedio (muscolo della staffa) e muscolo tensore del timpano (inserito sul martello) Orecchio interno: coclea e organo del Corti La funzione dell’orecchio esterno è quella di convogliare le onde sonore al timpano che chiude sul fondo l’orecchio esterno separandolo da quello medio.
: padiglione auricolare e meato uditivo esterno Orecchio medio (regione mastoidea del lobo temporale): tromba di Eustachio, catena degli ossicini (martello, incudine e staffa), muscolo stapedio (muscolo della staffa) e muscolo tensore del timpano (inserito sul martello) Orecchio interno: coclea e organo del Corti La funzione dell’orecchio esterno è quella di convogliare le onde sonore al timpano che chiude sul fondo l’orecchio esterno separandolo da quello medio.")
11
La membrana timpanica ha la forma di un cono ed è mantenuta tesa in modo da poter vibrare quando intercetta un’onda sonora. Ha un elevato potere di smorzamento, cioè quando lo stimolo cessa anche le vibrazioni cessano immediatamente. La membrana timpanica ha una superficie (55mm 2 ) che è circa 15 volte superiore a quella della finestra rotonda (3.5mm 2 ), dando una notevole amplificazione al segnale
 che è circa 15 volte superiore a quella della finestra rotonda (3.5mm 2 ), dando una notevole amplificazione al segnale.")
12
L’orecchio medio è costituito dalla catena degli ossicini: martello, incudine e staffa. Il manico del martello si articola al centro del timpano e la testa del martello all’incudine. L’incudine a sua volta si articola sulla staffa e il piede di questa sulla finestra ovale che chiude l’orecchio medio.

13
L’insieme degli ossicini costituisce un’ulteriore sistema di amplificazione del segnale acustico. Il manico del martello è sempre tenuto indietro dal muscolo tensore del timpano, che mantiene il timpano in tensione. L’onda sonora passando da un mezzo aereo ad un mezzo liquido perderebbe gran parte dell’energia incidente a causa della diversa impedenza acustica dei due mezzi. Il sistema formato da membrana timpanica, catena degli ossicini, membrana della finestra ovale costituisce un sistema amplificatore di pressione compensando la perdita di energia (l’amplificazione è di circa 22 volte fra timpano e finestra ovale).
..")
14
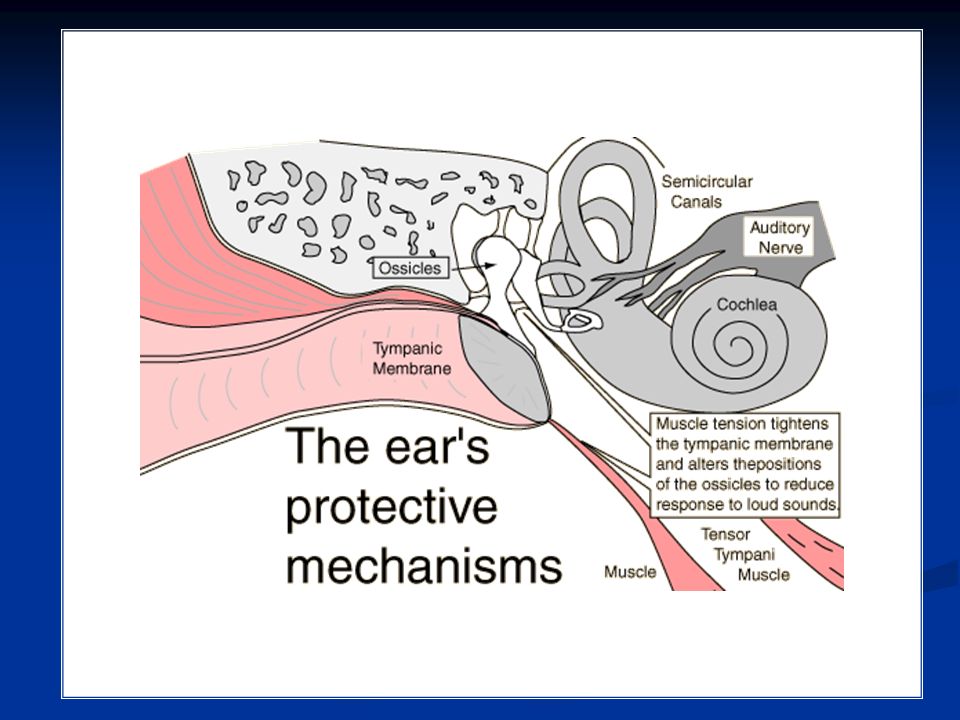
Quindi la differenza di superficie fra timpano e membrana della finestra ovale e catena degli ossicini fornisce un sistema che permette la minima dissipazione possibile di energia nel passaggio di un’onda sonora fra un mezzo aereo (orecchio esterno e medio) e uno liquido (orecchio interno). Quando un suono particolarmente basso viene trasmesso attraverso la catena degli ossicini, si genera un riflesso entro 40-80ms che determina contrazione del muscolo stapedio e del tensore del timpano. Il primo spinge avanti la staffa, mentre il secondo tira indietro il martello.

15
In questo modo le due forze opposte determinano una certa rigidità del sistema degli ossicini che impediscono la trasmissione di suoni troppo bassi. In tal modo: La coclea è protetta da vibrazioni troppo basse che la danneggerebbero Sono mascherati rumori di fondo troppo bassi che determinerebbero un noise di base. Inoltre questo funzionamento tra i due muscoli determina decremento di sensibilità di un soggetto verso la sua propria voce.

19
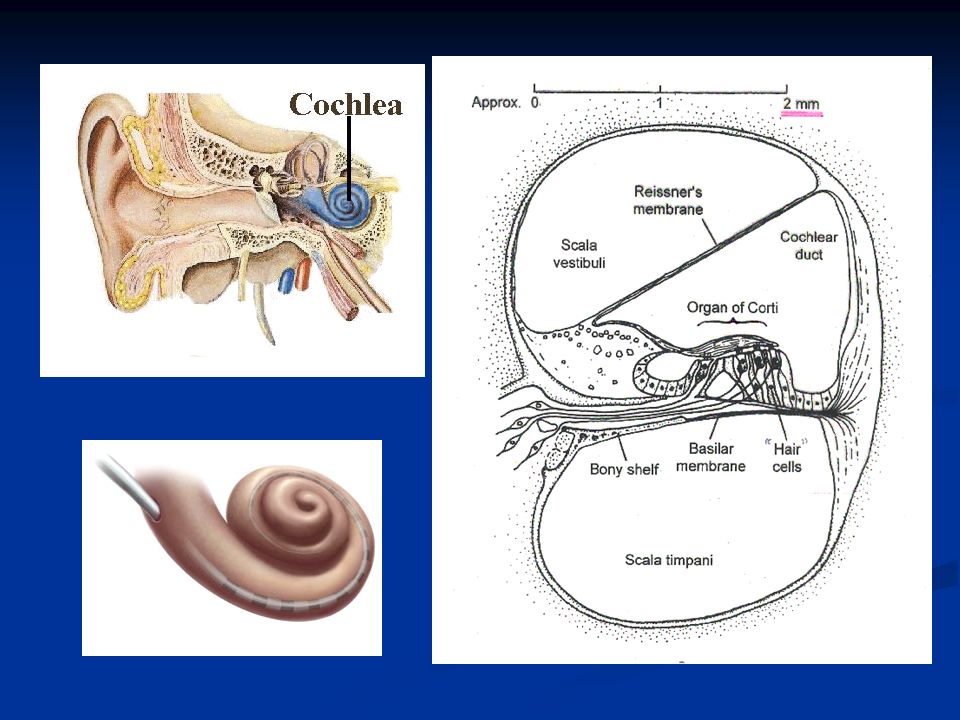
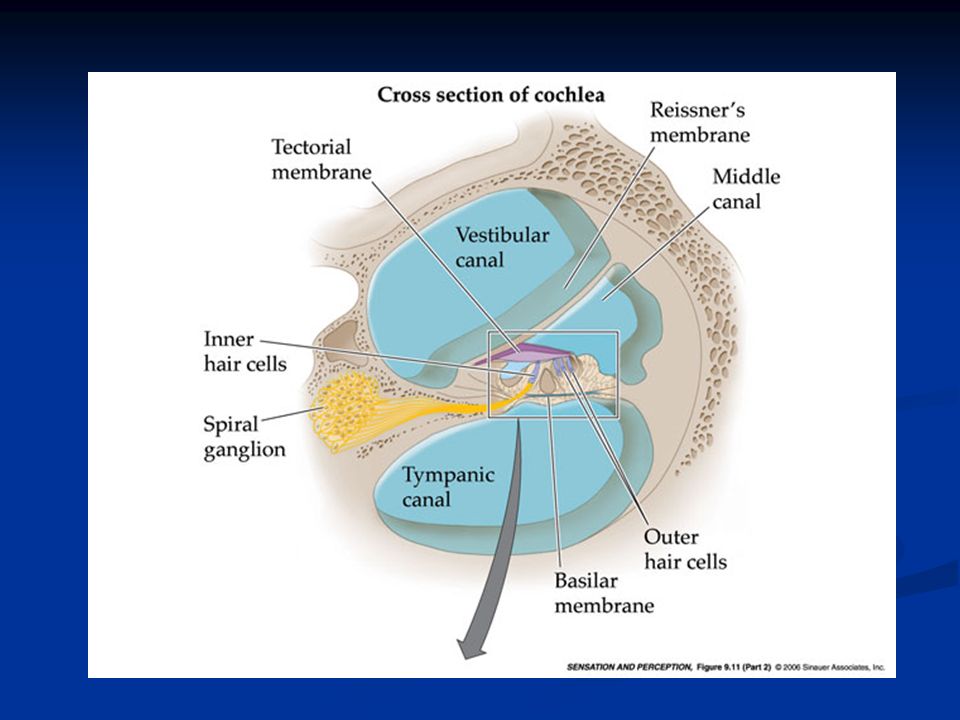
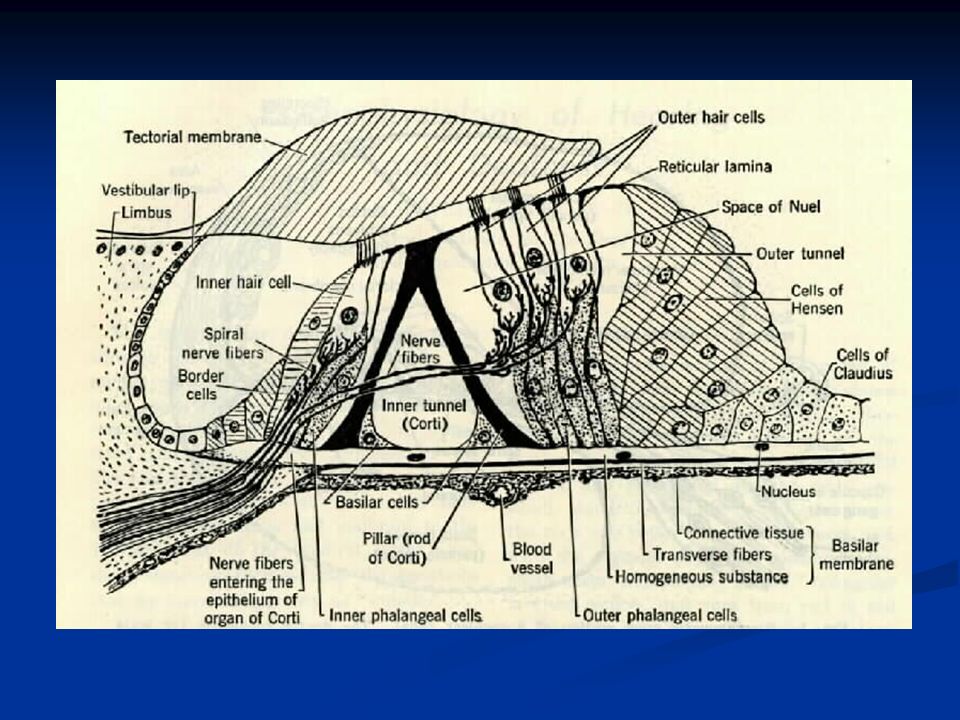
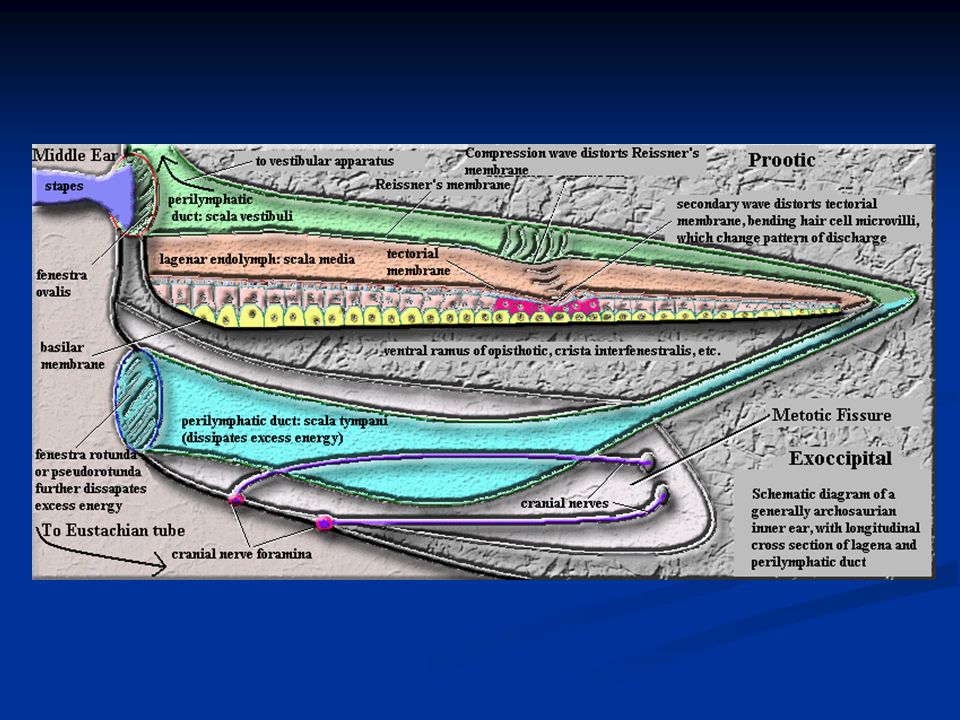
Orecchio interno L’orecchio interno è costituito dalla coclea, arrotolata a forma di spirale sull’osso modiolo. È suddivisa in tre rampe: rampa vestibolare rampa media rampa timpanica La rampa vestibolare e quella media sono separate dalla membrana di Reissner, mentre rampa media e timpanica sono separate dalla membrana basilare sulla cui superficie si trova l’organo del Corti che contiene i recettori uditivi, cioè le cellule cigliate.

21
La scala vestibolare e la scala timpanica decorrono insieme fino al vertice della coclea (ELICOTREMA) dove le due scale sono fra loro in comunicazione
 dove le due scale sono fra loro in comunicazione")
25
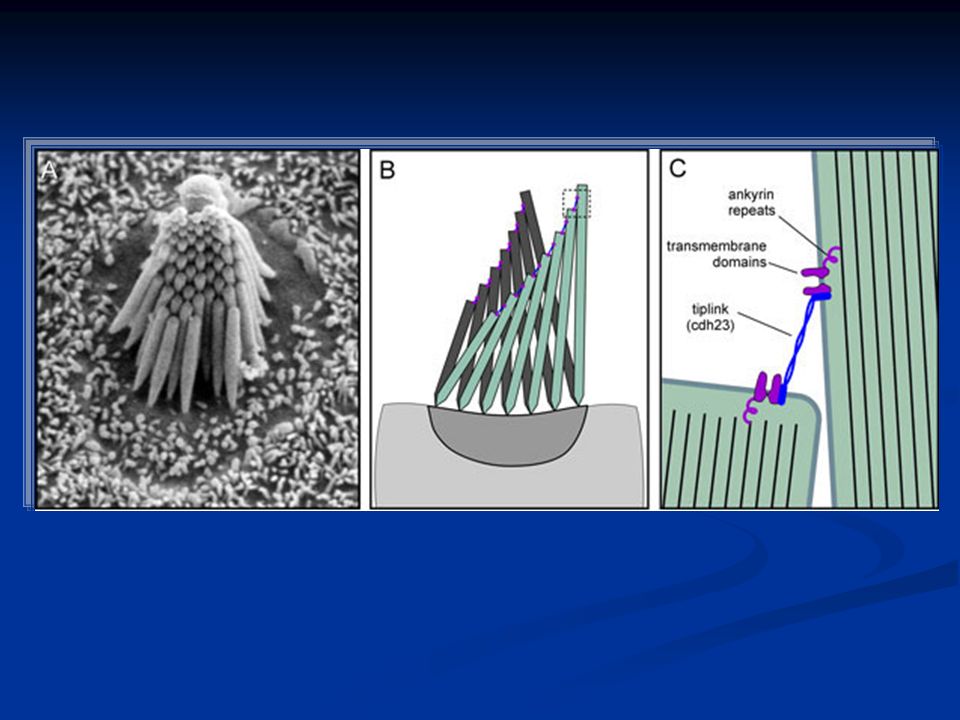
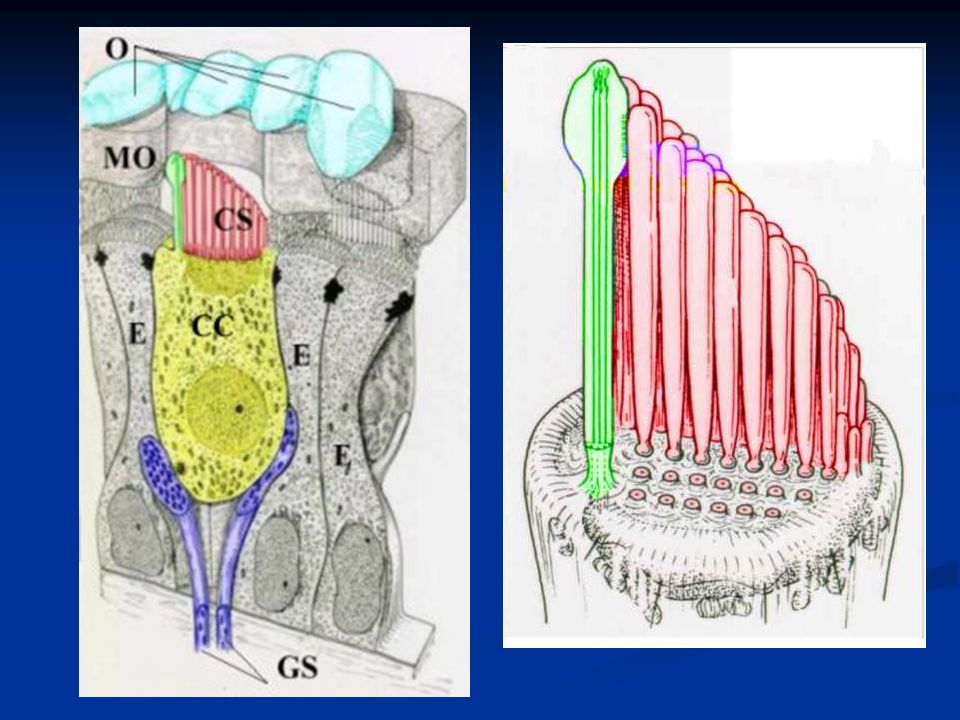
I recettori: cellule cigliate I recettori per il suono sono meccanocettori e sono le cellule cigliate. Si tratta di cellule altamente specializzate caratterizzate sulla parte apicale dalla presenza di ciglia di lunghezza decrescente. Manca nelle cellule cigliate di Mammifero il chinociglio (9 + 2 coppie di microtubuli), mentre sono presenti solo stereociglia con i microtubuli disposti senza organizzazione particolare.
, mentre sono presenti solo stereociglia con i microtubuli disposti senza organizzazione particolare..")
26
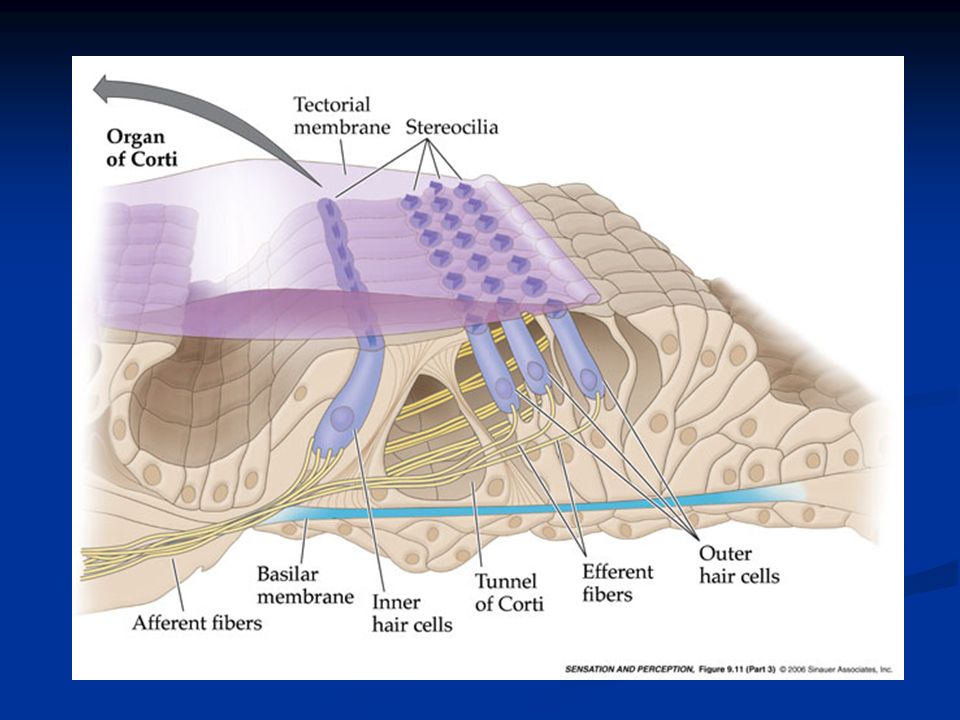
Nella coclea si distinguono tre ordini di cellule cigliate esterne e un ordine di cellule cigliate interne.

27
Cellule cigliate Le cellule cigliate interne convertono la dislocazione meccanica in attività elettrica e svolgono quindi il ruolo di trasduttori inviando segnali alle fibre afferenti (VIII nervo cranico). La maggior parte delle fibre afferenti contrae sinapsi con le cellule c. interne.

28
Le cellule cigliate esterne hanno un ruolo chiave nel raggiungere un’elevata sensibilità e nella sintonizzazione rapida. La maggior parte delle fibre efferenti (dai centri uditivi centrali alla coclea) contraggono sinapsi con le cellule c. esterne
 contraggono sinapsi con le cellule c. esterne.")
29
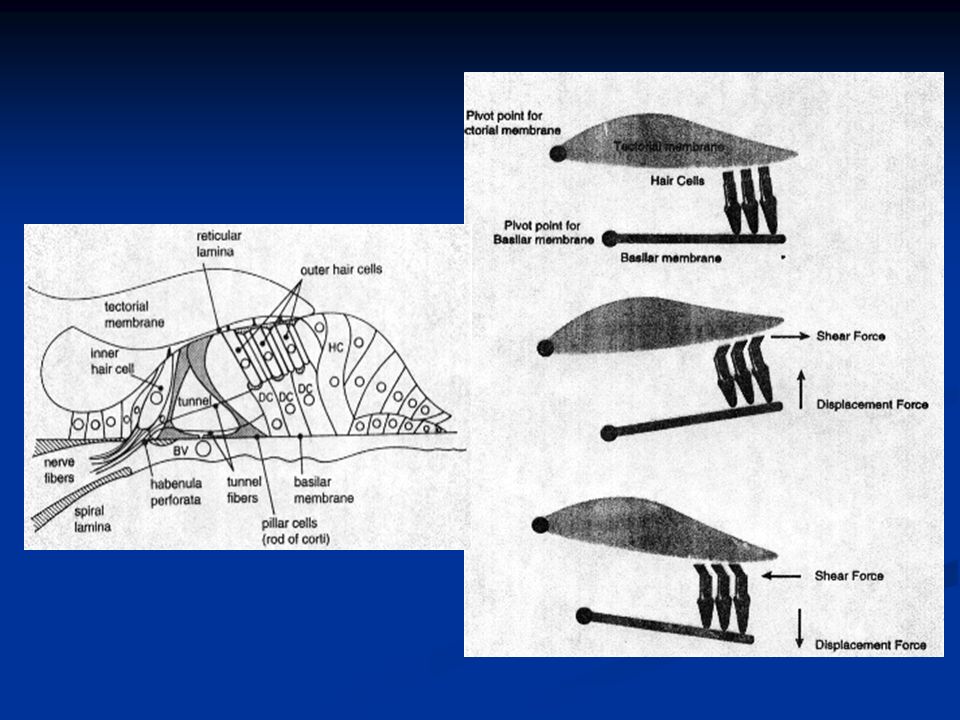
Le stereociglia sono impiantate in una struttura che prende il nome di membrana tectoria, di natura gelatinosa. Quando la membrana basilare su cui sono alloggiate le cellule si muove oscillando, anche le cellule cigliate si muovono relativamente alle stereociglia che invece restano ferme e impiantate nella membrana tectoria.

30
Le cellule cigliate sono fatte in modo tale che quando le stereociglia deflettono dalla più bassa verso la più alta il potenziale di membrana depolarizza, mentre quando deflettono dalla più alta verso la più bassa il potenziale iperpolarizza. In questo modo un’onda sonora che ha un picco positivo e un picco negativo può essere perfettamente codificata in termini di potenziali di membrana.

31
Questo è possibile grazie al fatto che in primo luogo le stereociglia sono legate fra loro da sottili filamenti di actina e poi alla particolare presenza di canali del potassio. Vediamo come.

33
Nell’organo del Corti le cellule cigliate presentano la parte apicale che guarda verso la rampa media dove si trova un liquido detto endolinfa molto ricco di K + (circa 150mM) secreto dalla stria vascolare e la parte basale che guarda verso la rampa timpanica dove si trova un liquido detto perilinfa a normale composizione dei liquidi extracellulari.
 secreto dalla stria vascolare e la parte basale che guarda verso la rampa timpanica dove si trova un liquido detto perilinfa a normale composizione dei liquidi extracellulari.")
34
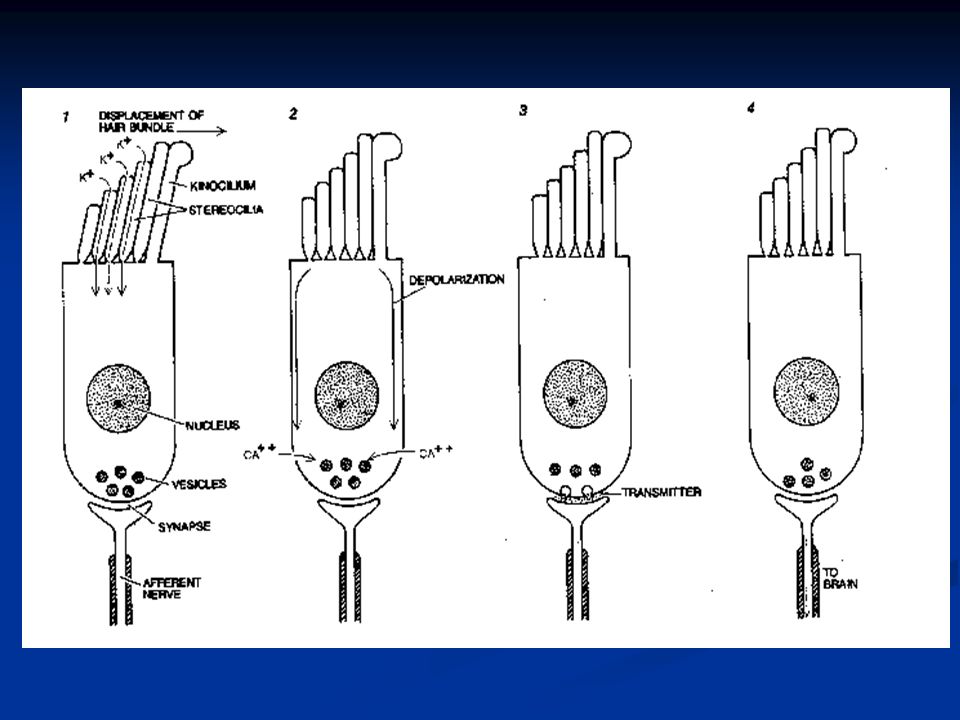
Per la posizione particolare delle cellule cigliate ecco quello che succede. Durante la fase ascendente dell’onda le stereociglia deflettono dalla più bassa verso la più alta e in questo movimento stirano meccanicamente canali del K + localizzati sulla superficie laterale delle ciglia più alte. Siccome il potassio extracellulare nella rampa media è superiore a quello intracellulare, il K + ENTRA nella cellula depolarizzando la membrana.

35
K+K+K+K+ K+K+K+K+ K+K+K+K+ K+K+K+K+ Condizione di riposo: media quantità di canali aperti Semionda positiva: depolarizzazione per aumento dei canali aperti Semionda negativa: chiusura totale dei canali e iperpolarizzazione

37
Schema delle stereociglia e apertura dei canali potassio

39
La depolarizzazione determina l’apertura di canali del Ca 2+ potenziale-dipendenti rivolti verso la rampa timpanica. Il calcio innesca l’esocitosi di neurotrasmettitore verso il neurone del I ordine. L’aumento del calcio a sua volta determina l’attivazione di canali K + Ca 2+ -dipendenti rivolti verso la perilinfa della rampa timpanica, dove la concentrazione di ioni K + è quella normale. Il K + esce dalla cellula e la membrana può ripolarizzare.

40
Durante la fase discendente dell’onda il processo è inverso, nel senso che i canali K + verso l’endolinfa si chiudono iperpolarizzando la cellula. Va notato che questi canali del K + sono aperti in una certa percentuale, nella cellula non stimolata, in modo tale che questo numero sia suscettibile di aumentare o diminuire a seconda della deflessione delle stereociglia in un senso o nell’altro.

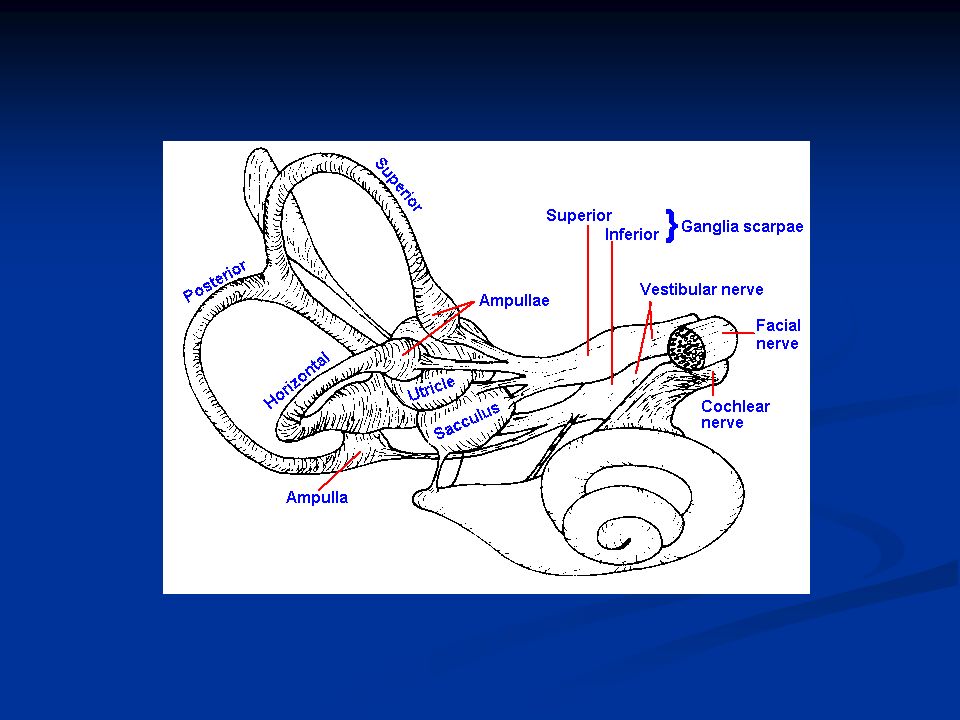
41
Vediamo adesso come viene veicolata l’informazione sonora. L’onda sonora mette in vibrazione la membrana timpanica e questa vibrazione, opportunamente amplificata, trasmette la vibrazione alla catena degli ossicini, sempre propagandosi nel mezzo aereo. Quando la vibrazione arriva al piede della staffa, questa si ripercuote sulla membrana della finestra ovale, da dove inizia l’orecchio interno. Qui si passa ad un mezzo liquido, in quanto, come detto troviamo perilinfa nella rampa vestibolare e timpanica ed endolinfa nella rampa media.

42
La perilinfa è un liquido e pertanto incomprimibile. A questo punto il liquido viene messo in movimento dal propagarsi dell’onda e così facendo determina un’oscillazione della membrana basilare. Le cellule cigliate si spostano uin maniera sincrona con la membrana basilare con conseguente dislocazione delle stereociglia e variazione del potenziale di recettore.

44
Proprietà di risonanza della membrana basilare La percezione e codificazione dell’intensità di un suono è quindi possibile grazie alla maggiore o minore dislocazione delle ciglia e quindi all’ampiezza maggiore o minore del potenziale di recettore. Ma come viene codificata la frequenza di un suono?

45
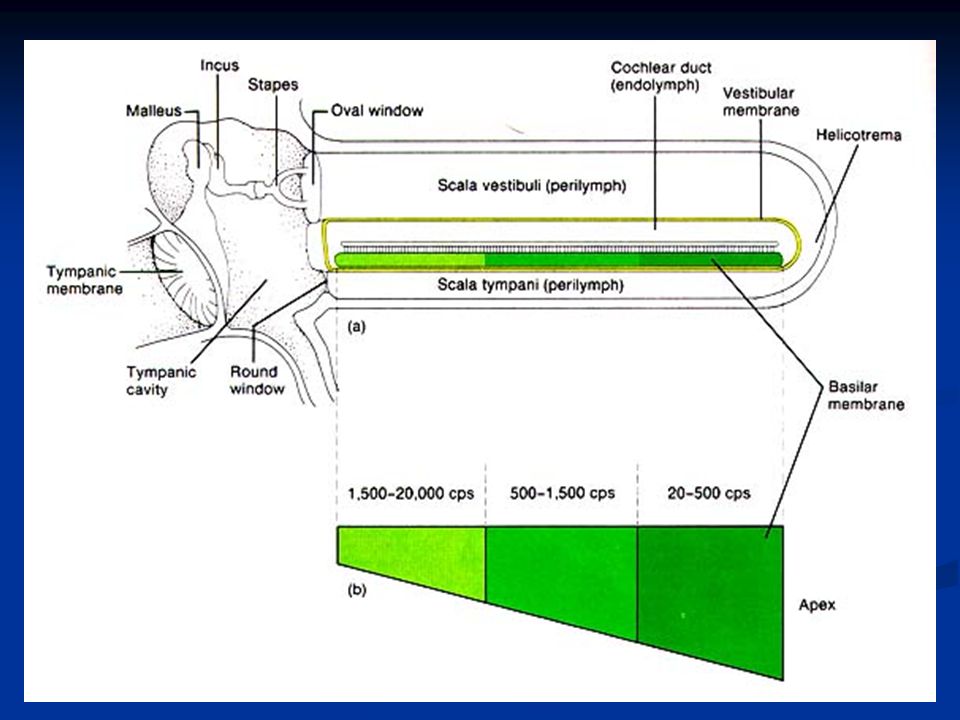
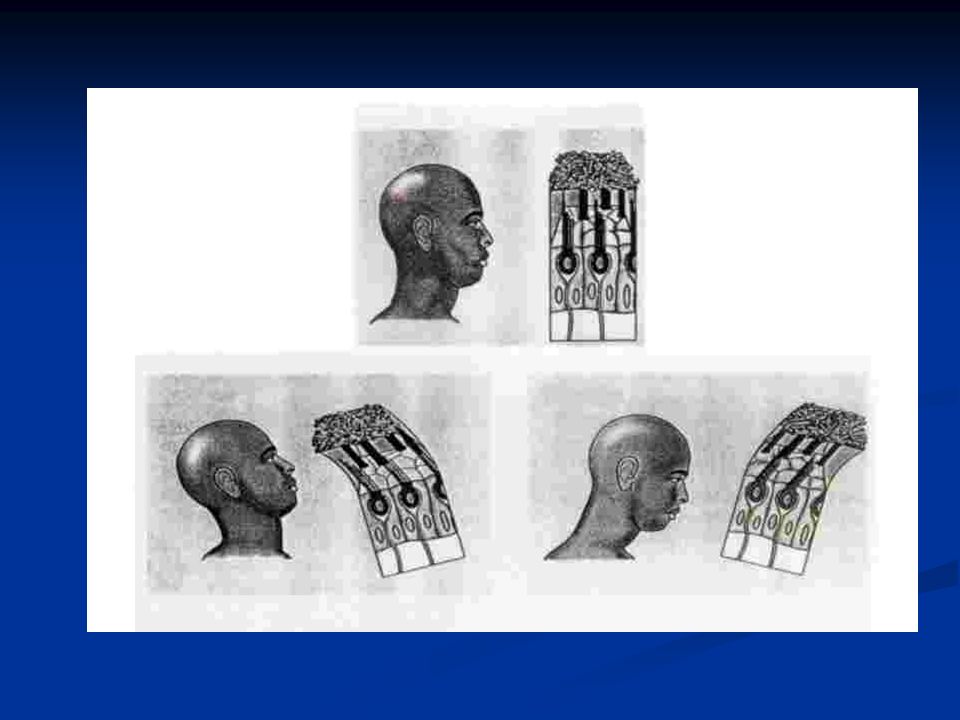
Le cellule cigliate sono distribuite lungo la coclea con una precisa organizzazione spaziale (organizzazione tonotopica), per cui le cellule con ciglia più corte e rigide sono localizzate verso la base della coclea e sono più sensibili a frequenze acute. Le cellule con ciglia più lunghe e sottili sono invece verso l’apice della coclea (lunghezza coclea circa 13mm) e sono sintonizzate su frequenze più basse. Inoltre la coclea è più stretta verso la staffa e verso l’elicotrema si allarga. Poiché la rigidità di una struttura dipende, fra l’altro, dalla sua larghezza, la membrana basilare alla base è più rigida mentre verso l’apice, dove è più larga, è più flessibile.
 e sono sintonizzate su frequenze più basse. Inoltre la coclea è più stretta verso la staffa e verso l’elicotrema si allarga. Poiché la rigidità di una struttura dipende, fra l’altro, dalla sua larghezza, la membrana basilare alla base è più rigida mentre verso l’apice, dove è più larga, è più flessibile..")
47
Quando l’onda raggiunge la zona di membrana basilare che ha caratteristiche più adatte per quella determinata frequenza, questa fa vibrare tanto facilmente la membrana basilare che l’energia dell’onda si estingue completamente. La figura mostra i diversi modelli di onda viaggiante per suoni di differenti frequenze.

48
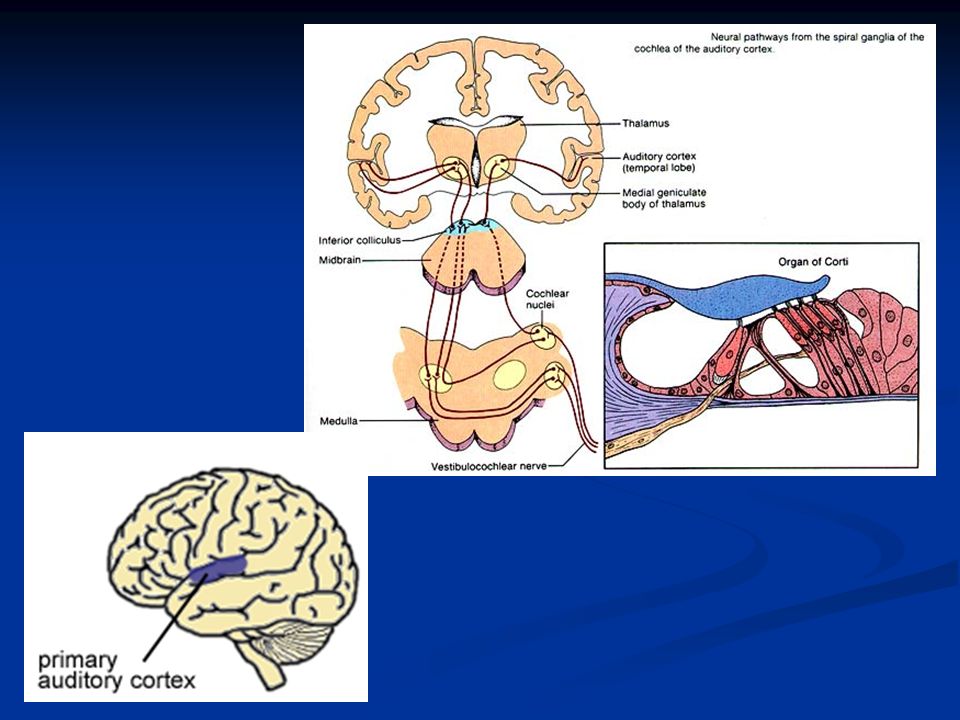
Vie uditive

49
Le fibre sensoriali sono rappresentate dalle fibre afferenti dell’VIII nervo cranico (nervo uditivo) che comprende una parte cocleare e una parte vestibolare. Il nervo cocleare deriva da neuroni che hanno il soma localizzato a livello del ganglio spiralato della coclea e i cui dendriti contraggono sinapsi con le cellule cigliate.

50
Selezione delle frequenze Le fibre cocleari hanno una selettività verso certe frequenze poiché rispondono a stimoli provenienti da zone ristrette della membrana basilare Le fibre cocleari hanno una selettività verso certe frequenze poiché rispondono a stimoli provenienti da zone ristrette della membrana basilare Hanno una frequenza caratteristica, cioé una frequenza a cui la loro soglia di attivazione è minore Hanno una frequenza caratteristica, cioé una frequenza a cui la loro soglia di attivazione è minore

51
Tronco encefalico e corteccia uditiva La capacità di localizzare i suoni dipende dall’interazione dei segnali provenienti dalle due orecchie L’informazione proveniente da entrambi i nervi uditivi è integrata a livello di tronco encefalico. I nuclei del tronco encefalico coinvolti sono: nucleo cocleare, nucleo olivare superiore, collicolo inferiore, corpo genicolato mediale

52
1. Nucleo cocleare del tronco: l’input proviene dal solo orecchio omolaterale La maggior parte degli assoni dei neuroni del II ordine a questo livello decussa 2. Complesso olivare superiore: convergenza dei segnali provenienti dalle due orecchie Importante per la localizzazione del suono

53
3. Collicolo inferiore: Riceve inputs dal complesso olivare e dal nucleo cocleare Alcune, ma non tutte, le informazioni che arrivano al collicolo derivano dall’orecchio controlaterale Interviene nella localizzazione del suono e combina le informazioni provenienti da aree inferiori del tronco

54
4. Nucleo genicolato mediale: Ultima tappa prima di arrivare in corteccia Flusso bidirezionale di informazioni fra il n.g.m. e la corteccia uditiva Stretta integrazione a feed-back fra cervello e informazioni sensoriali dirette al cervello

55
Corteccia uditiva Localizzata a livello di Localizzata a livello di lobi temporali con notevole organizzazione tonotopica delle frequenze. Aree 41 e 42 di Brodmann Informazione bidirezionale fra corteccia e tronco Informazione bidirezionale fra corteccia e tronco

57
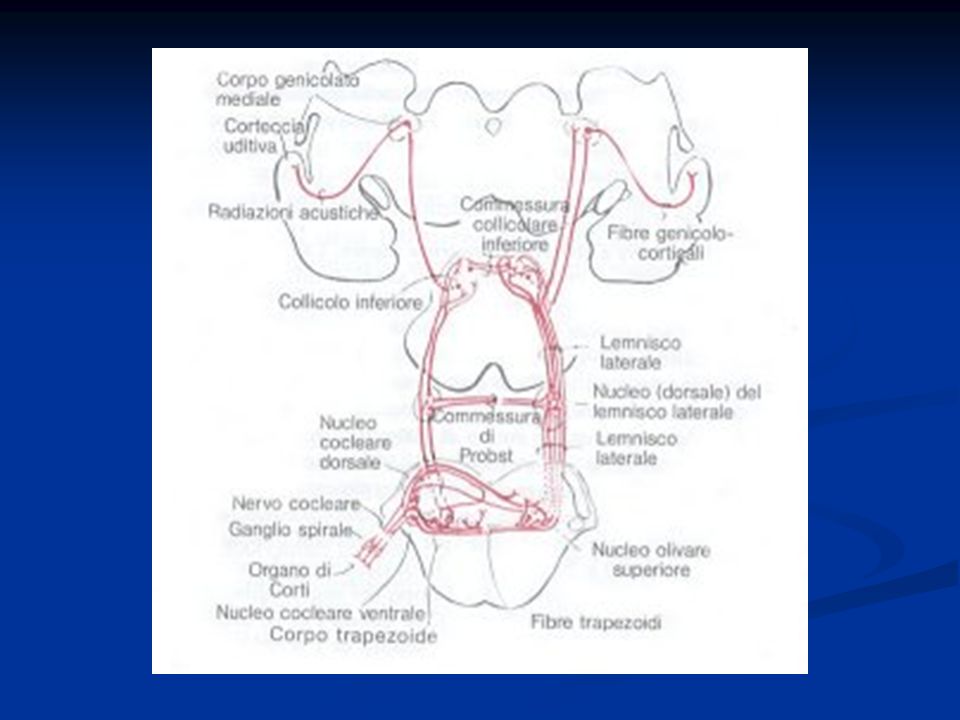
Le cellule cigliate sono innervate da fibre nervose il cui soma è localizzato nel ganglio spirale del Corti (neurone del I ordine). Gli assoni di questi neuroni formano il nervo acustico. Le fibre entrano nei nuclei cocleari dorsali e ventrali del bulbo dove contraggono sinapsi con i neuroni del II ordine. La maggior parte di questi neuroni invia assoni al nucleo olivare superiore controlaterale decussando a livello del corpo trapezoide. Altre fibre proiettano al nucleo olivare ipsilaterale. Da qui attraverso il lemnisco laterale e passando per il collicolo mesencefalico inferiore, le fibre proiettano al nucleo genicolato mediale del talamo da dove i neuroni del III ordine proiettano in corteccia uditiva.

58
Il ganglio del Corti si differenzia molto presto, prima dell’organo del Corti stesso. Esso è formato, nell’uomo da 30 a 35.000 neuroni bipolari di due tipi differenti: Neuroni di tipo I (che sono più del 90%): neuroni mielinici, piuttosto grandi, sono connessi alle cellule cigliate interne. Neuroni di tipo II: amielinici, picccoli connessi alle cellule cigliate esterne Entrambi i tipi inviano i loro assoni ai nuclei cocleari.
: neuroni mielinici, piuttosto grandi, sono connessi alle cellule cigliate interne. Neuroni di tipo II: amielinici, picccoli connessi alle cellule cigliate esterne Entrambi i tipi inviano i loro assoni ai nuclei cocleari..")
60
Il campo uditivo umano e i suoi limiti (diagramma di Wegel)
")
61
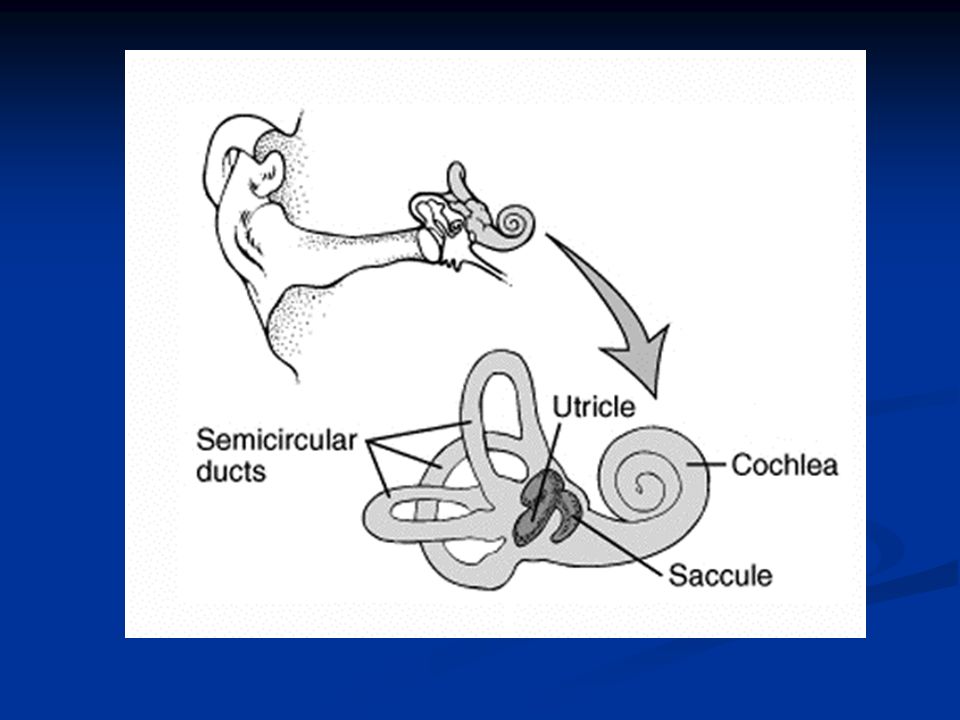
Sistema vestibolare

62
Il sistema vestibolare: struttura e funzione Il sistema vestibolare è un sofisticato apparato sensoriale che rileva informazioni relative alla posizione ed al movimento della testa e del corpo nello spazio. A differenza di altri sistemi sensoriali, come il sistema visivo o uditivo, non siamo generalmente coscienti delle informazioni trasmesse dall’apparato vestibolare al sistema nervoso centrale. Tuttavia, esse sono fondamentali per il mantenimento della postura e della stazione eretta, e per la coordinazione del movimento degli occhi con quello della testa.

63
Una disfunzione dell’apparato vestibolare induce, infatti, notevoli alterazioni dell’equilibrio, del senso dell’orientamento nello spazio e del controllo della motilità oculare, e si accompagna ad una serie di intense e fastidiose sensazioni, quali la nausea o le vertigini. Non comporta invece problemi di sordità perché i due sistemi veicolano informazioni del tutto differenti

64
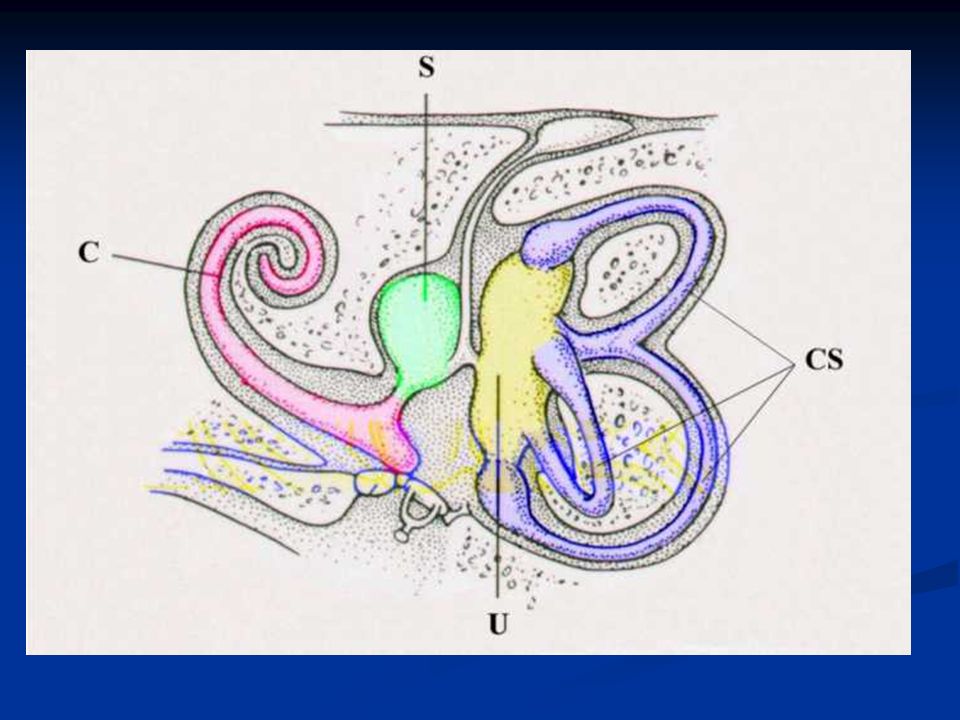
Le strutture sensoriali periferiche dell’apparato vestibolare sono strettamente imparentate con quelle dell’apparato uditivo. Gli organi recettoriali dei due apparati condividono, infatti, l’origine embrionale dal placode otico ectodermico. I recettori vestibolari sono situati all’interno di una complicata rete di canalicoli ossei, che costituisce il cosiddetto labirinto, scavato all’interno della rocca petrosa dell’osso temporale. All’interno di queste cavità ossee è contenuto un complicato sistema di canalicoli delimitati da membrane connettivali, chiamato labirinto membranoso.

65
I dotti del labirinto membranoso sono pieni di un particolare liquido detto endolinfa, mentre gli spazi esterni sono pieni di perilinfa. Al loro interno si trovano i recettori vestibolari, le cellule cigliate. Per registrare le informazioni relative al movimento della testa l’apparato vestibolare è dotato di due diverse strutture sensoriali: gli organi otolitici, che misurano i movimenti lineari, ed i canali semicircolari, che registrano quelli rotatori.

66
Più in particolare, i recettori degli organi otolitici rispondono ad accelerazioni lineari del capo, mentre quelli dei canali semicircolari sono sensibili ad accelerazioni angolari. Entrambi gli organi sensoriali contengono lo stesso tipo di cellula recettrice, ma sono caratterizzati da una particolare struttura anatomica che ne determina la specificità nel rilevare i movimenti lineari o rotatori.

68
Ricordiamo che il ciuffo delle cellule cigliate vestibolari presenta anche un lungo chinociglio, assente in quelle cocleari, dotato delle caratteristiche citologiche di un flagello. La deflessione verso il chinociglio induce una depolarizzazione del recettore e un conseguente aumento della frequenza di scarica nel nervo vestibolare. Al contrario, la deflessione in senso opposto, induce una iperpolarizzazione del recettore, il quale libera una minor quantità di neurotrasmettitore riducendo così l’attività del nervo vestibolare.

70
Gli organi otolitici Gli organi otolitici, contenuti nelle cavità labirintiche del sacculo e dell’utricolo sono sensibili alle accelerazioni lineari cui è sottoposto il capo. Questi stimoli comprendono le accelerazioni generate nel corso di movimenti di flessione o di traslazione lineare della testa e l’accelerazione di gravità, che esercita costantemente la sua azione sul corpo. All’interno delle cavità dell’utricolo e del sacculo è presente un ispessimento dell’epitelio, detto macula, che rappresenta la struttura recettoriale vera e propria.

72
I ciuffi di ciglia dei recettori sporgono dalla superficie della macula e sono immersi in una sostanza gelatinosa al di sopra della quale è posta un membrana fibrosa, la membrana otolitica, che contiene numerosi cristalli di carbonato di calcio detti otoconi. Gli otoconi rendono la membrana otolitica notevolmente più densa e pesante rispetto all’endolinfa, di modo che quando la testa si flette la forza di gravità determina uno spostamento relativo della membrana otolitica rispetto alla macula che si traduce in una deflessione dei ciuffi di ciglia

74
L’utricolo è sensibile a movimenti della testa sul piano orizzontale (la macula dell’utricolo è infatti posta sul piano orizzontale ), come flessioni laterali della testa o movimenti traslatori in senso laterale, mentre il sacculo (la macula del sacculo è posta sul piano verticale) risponde a movimenti sul piano verticale, come movimenti dal basso verso l’alto. Inoltre, le macule dell’utricolo e del sacculo dei due lati hanno una disposizione simmetrica e perciò all’eccitazione delle cellule cigliate di un lato corrisponde l’inibizione dei recettori controlaterali corrispondenti.

75
In questo modo, le informazioni relative ai movimenti del capo nelle diverse direzioni che arrivano al sistema nervoso centrale sono il risultato dall’attività combinata degli organi recettoriali dei due lati.

76
Posizione statica A riposo le stereociglia rimangono erette e la cellule cigliata è solo parzialmente depolarizzata. Il neurone afferente scarica potenziali d’azione ad una frequenza tonica.

77
Depolarizzazione cellula ciliata, aumento della frequenza dei potenziali d’azione. Iperpolarizzazione cellula ciliata, riduzione della frequenza dei potenziali d’azione. Accelerazione del capo in avanti Accelerazione del capo all’indietro

78
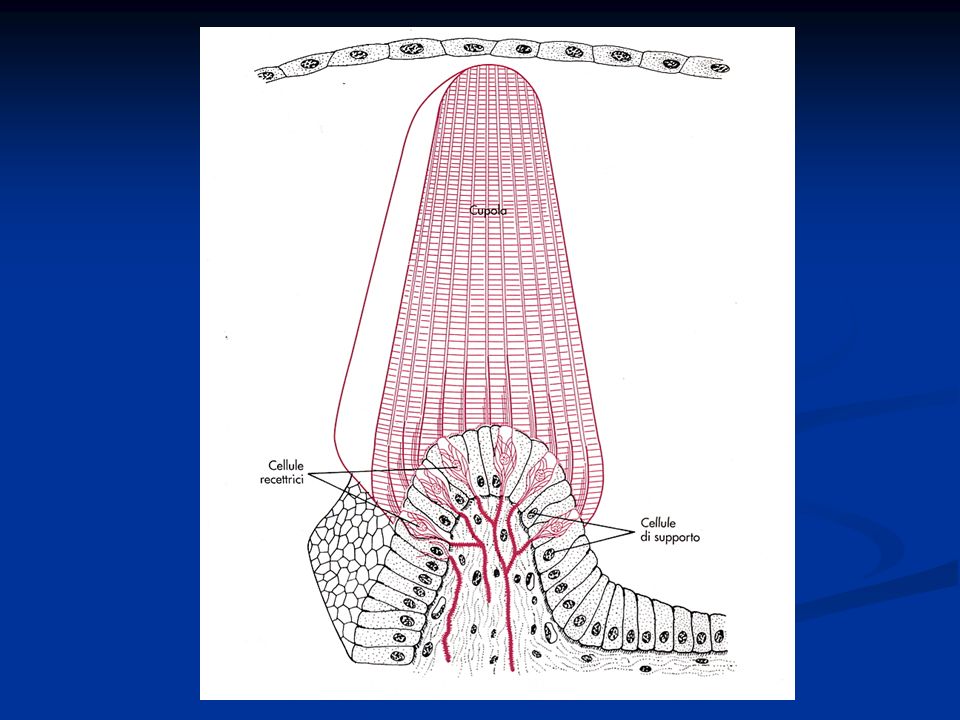
I canali semicircolari I canali semicircolari (laterale, superiore e posteriore) sono tre sottili condotti di forma circolare che si aprono nella cavità dell’utricolo. In prossimità dello sbocco nell’utricolo ogni canale presenta una dilatazione detta ampolla, nella quale è contenuto l’epitelio sensoriale. All’interno dell’ampolla l’epitelio forma un rilievo detto cresta ampollare, sulla quale sono situati i recettori. I ciuffi di ciglia delle cellule recettrici sono inglobati in una struttura gelatinosa, detta cupola, che oblitera completamente il lume del condotto in corrispondenza della cresta

80
N.B.: La cupola, a differenza della membrana otolitica, ha la stessa densità dell’endolinfa. Di conseguenza, accelerazioni lineari non determinano alcuno spostamento relativo della cupola rispetto all’endolinfa e quindi non stimolano i recettori. Per contro, la stimolazione di questi recettori sarà prodotta da accelerazioni angolari, alle quali è sottoposto il capo nel corso di movimenti rotatori

81
La stimolazione di questi recettori è prodotta da accelerazioni angolari, alle quali è sottoposto il capo nel corso di movimenti rotatori. Infatti, quando il capo ruota sul piano di uno dei canali semicircolari, l’endolinfa, a causa della sua inerzia, segue con ritardo il movimento del dotto membranoso in cui è contenuta. Si genera così una corrente endolinfatica, diretta lungo l’asse del canale in direzione opposta a quella del movimento del capo, la quale induce una distorsione della cupola che si traduce nella deflessione dei ciuffi di ciglia in essa inglobati

83
I canali semicircolari sono organizzati in modo che ogni canale su lato sx ha la sua controparte sul lato dx. Ciascuna di queste tre coppie lavora con un metodo push-pull: quando un canale è stimolato la sua parte corrispondente sull’altro lato è inibita e viceversa I canali semicircolari sono organizzati in modo che ogni canale su lato sx ha la sua controparte sul lato dx. Ciascuna di queste tre coppie lavora con un metodo push-pull: quando un canale è stimolato la sua parte corrispondente sull’altro lato è inibita e viceversa

84
Movimento relativo dell’endolinfa

85
Riflesso vestibolo-oculare Trattasi di un movimento riflesso degli occhi volto a stabilizzare l’immagine sulla retina durante i movimenti del capo producendo un movimento degli occhi nella direzione opposta al movimento del capo. In questo modo si preserva l’immagine al centro del campo visivo. Il riflesso subentra entro 10ms, uno dei riflessi più veloci nel corpo umano. Trattasi di un movimento riflesso degli occhi volto a stabilizzare l’immagine sulla retina durante i movimenti del capo producendo un movimento degli occhi nella direzione opposta al movimento del capo. In questo modo si preserva l’immagine al centro del campo visivo. Il riflesso subentra entro 10ms, uno dei riflessi più veloci nel corpo umano.

86
Rotazione del capo

Similar presentations






 Convenzioni di uso dei registri Utilizzo.>")

 Corso di formazione Progetto Low Noise PON 01_01878 Dott.ssa La Rosa Daniela Dipartimento.>")










