Download presentation
Presentation is loading. Please wait.

1
Food Biotechnology Dr. Tarek Elbashiti 5. Metabolic Engineering of Bacteria for Food Ingredients

2
Metabolic engineering has been defined as the directed improvement of product formation or cellular properties through the modification of specific biochemical reactions or the introduction of new ones with the use of recombinant DNA technology. ME can rationally improve the properties of the microorganisms and their efficiency for producing different products. This illustrates the tremendous potential of ME for improving the bacteria used by the food industry. Food additives produced by bacteria can be incorporated into foods as nutritional supplements, flavor enhancers, texturizers, acidulants, preservatives, emulsifiers, surfactants, thickeners, or functional food ingredients.

3
Applying ME principles and tools to the production of food ingredients by bacteria has resulted in the efficient production of both native and totally novel products by several cultures, including strains of lactic acid bacteria (LAB), Escherichia coli, Bacillus subtilis, and Corynebacterium glutamicum. The examples in this chapter (summarized in Table 5.2) will cover the analysis and modification of central metabolic pathways, biosynthetic pathways, and transport systems involved in producing amino acids, organic acids, vitamins, carbohydrates, bacteriocins, low calorie sugars, and aroma compounds.
 will cover the analysis and modification of central metabolic pathways, biosynthetic pathways, and transport systems involved in producing amino acids, organic acids, vitamins, carbohydrates, bacteriocins, low calorie sugars, and aroma compounds..")
4
AMINO ACIDS Amino acids have a great variety of current and potential uses as food, pharmaceuticals, and animal feed. Their main application field is food, where about 50% of the product is applied. They can be used as nutritional supplements, flavor enhancers, sweeteners, and in pre and postoperative nutrition therapy. This section will discuss using ME to produce some of these amino acids.

5
Recent progress in this field can be divided into three categories: (1) modification of central metabolic pathways, (2) modification of biosynthetic pathways, and (3) modification of transport systems. Figures 5.1, 5.2, and 5.3 summarize some of the examples discussed below.

6
Modification of Central Metabolic Pathways From reasoning based on metabolic pathways structure, rerouting a carbon source to produce a desired amino acid should start by increasing the availability of precursor metabolites, energy, and reducing equivalents used in its synthesis. Central metabolic pathways meet these criteria, and therefore engineering central metabolism is essential for the efficient production of amino acids.

9
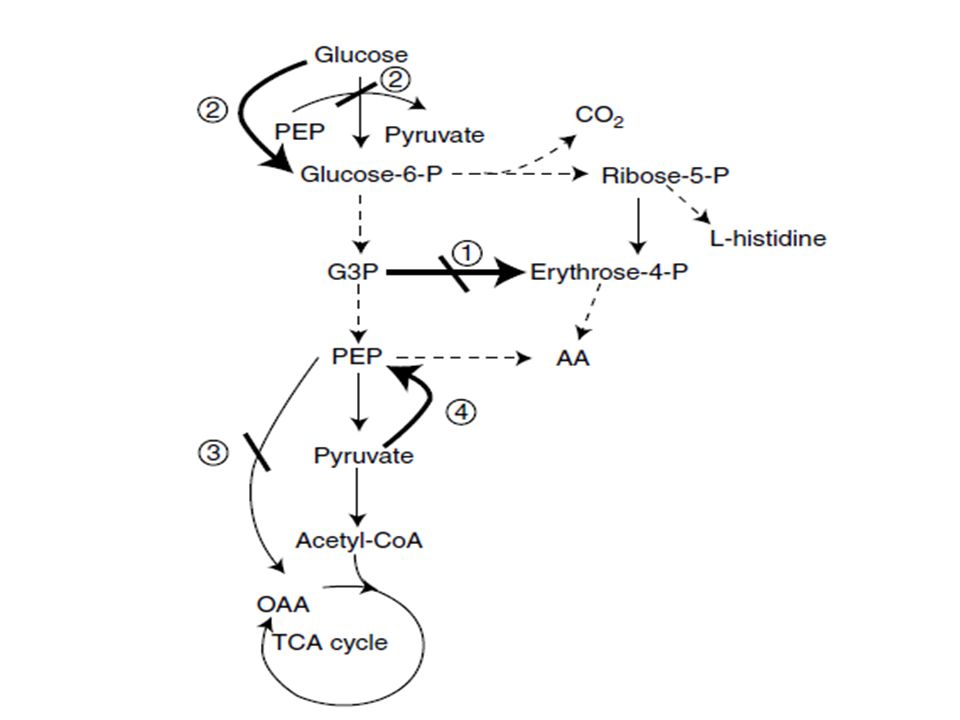
The analytical tools included in the ME toolbox have played an essential role in elucidating the function of different central pathways and suggesting useful strategies for redirecting carbon flow toward the biosynthesis of amino acids. For example, it has been shown that the pentose phosphate pathway (PPP) supports higher fluxes during the production of L-lysine compared to the production of L-glutamic acid in C. glutamicum. This was attributed to the higher requirements of reducing power (NADPH) in the production of L-lysine. Another example is improving aromatic amino acids and L-histidine production in C. glutamicum by increasing the availability of their precursor metabolites, erythrose 4- phosphate and ribose 5-phosphate, respectively, as well as NADPH.
 supports higher fluxes during the production of L-lysine compared to the production of L-glutamic acid in C. glutamicum. This was attributed to the higher requirements of reducing power (NADPH) in the production of L-lysine. Another example is improving aromatic amino acids and L-histidine production in C. glutamicum by increasing the availability of their precursor metabolites, erythrose 4- phosphate and ribose 5-phosphate, respectively, as well as NADPH..")
10
This can be done by modifying the flux through the PPP, either by increasing the activity of transketolase (and providing more erythrose 4- phosphate for aromatic amino acids biosynthesis) or by decreasing the activity of transketolase (and providing more ribose 5- phosphate for L-histidine biosynthesis) as shown in Figure 5.1, strategy 1. Both approaches have produced C. glutamicum strains with an increased capacity for making aromatic amino acids and L-histidine. Figure 5.1 shows additional examples of engineering central metabolic pathways to increase the availability of precursor metabolites used to synthesize aromatic amino acids in C. glutamicum.

12
In general, these strategies are based on increasing the availability of erythrose 4- phosphate (strategy 1, Figure 5.1), phosphoenolpyruvate (strategies 2, 3, and 4, Figure 5.1), and ribose-5-P (strategy 1 Figure 5.1) by inactivating the enzymes involved in their consumption and/or amplifying the enzymes involved in their production.
, phosphoenolpyruvate (strategies 2, 3, and 4, Figure 5.1), and ribose-5-P (strategy 1 Figure 5.1) by inactivating the enzymes involved in their consumption and/or amplifying the enzymes involved in their production.")
13
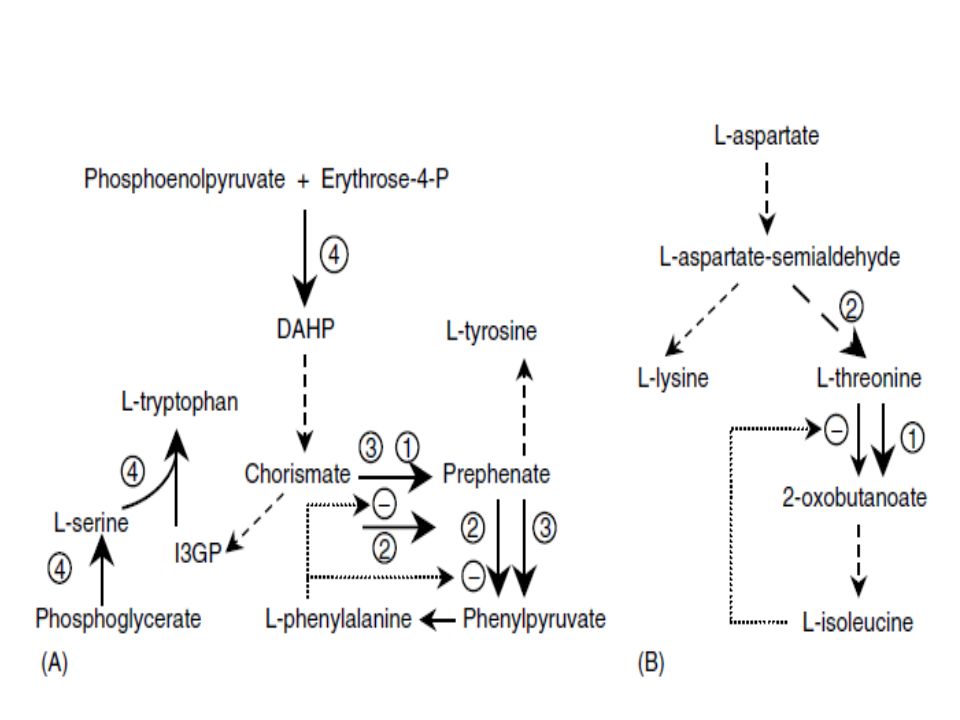
Modification of Biosynthetic Pathways After engineering central metabolic pathways, a sufficient supply of precursor metabolites, energy, and reducing power is ensured, and efforts then need to be focused on engineering biosynthetic pathways that convert precursor metabolites into amino acids. Several strategies have been used to achieve this goal, and some of them are illustrated in Figure 5.2. For example, the gene that encodes a rate-limiting enzyme can be amplified, resulting in the release of a bottleneck. Ozaki et al. and Ikeda et al. used this strategy to improve the production of L-phenylalanine in C. glutamicum. Overexpression of the gene that encodes chorismate mutase in C. glutamicum K38 resulted in a 50% increase in the yield of L-phenylalanine.

15
On the other hand, introducing heterologous enzymes subject to different regulatory mechanisms can also result in the release of a bottleneck. For example, overexpressing a mutated (insensitive to L- phenylalanine) E. coli gene that encoded the bifunctional enzyme chorismate mutase–prephenate dehydratase in C. glutamicum KY10694 led to a 35% increase in the production of L-phenylalanine. In addition, expressing the E. coli catabolic threonine dehydratase (insensitive to L-isoleucine) in C. glutamicum resulted in increased isoleucine production. A second strategy could be the redirection of metabolic flux in a branch point. Ikeda and Katsumata engineered a tryptophan- producing mutant of C. glutamicum to produce L-tyrosine or L-phenylalanine in abundance
 E. coli gene that encoded the bifunctional enzyme chorismate mutase–prephenate dehydratase in C. glutamicum KY10694 led to a 35% increase in the production of L-phenylalanine. In addition, expressing the E. coli catabolic threonine dehydratase (insensitive to L-isoleucine) in C. glutamicum resulted in increased isoleucine production. A second strategy could be the redirection of metabolic flux in a branch point. Ikeda and Katsumata engineered a tryptophan- producing mutant of C. glutamicum to produce L-tyrosine or L-phenylalanine in abundance.")
16
(26 and 28 g/L, respectively) by overexpressing the branch-point enzymes (chorismate mutase and prephenate dehydratase), catalyzing the conversion of the common intermediate chorismate into tyrosine and phenylalanine. Using a similar approach, Katsumata et al. produced threonine using a lysine-producing C. glutamicum strain, by amplifying a threonine biosynthetic operon. Other strategies could include introducing heterologous enzymes that use different cofactors than those used by the native enzyme as well as amplifying the enzyme that catalyzes the steps linking central metabolism and the biosynthetic pathway.

17
Ikeda et al. achieved a 61% increase in tryptophan yield (50 g/L of tryptophan) in a tryptophan-producing C. glutamicum KY10894 by coexpressing two enzymes catalyzing the initial steps in the biosynthesis of aromatic amino acids (3-deoxy-D-arabinoheptulosonate 7-phosphate synthase) and serine (3- phosphoglycerate dehydrogenase) together with tryptophan biosynthetic enzymes.
 in a tryptophan-producing C. glutamicum KY10894 by coexpressing two enzymes catalyzing the initial steps in the biosynthesis of aromatic amino acids (3-deoxy-D-arabinoheptulosonate 7-phosphate synthase) and serine (3- phosphoglycerate dehydrogenase) together with tryptophan biosynthetic enzymes..")
18
Modification of Transport Systems: By altering amino acid transport systems, one could expect to decrease their intracellular concentration and avoid feedback inhibition. The following two examples illustrate that show the significance of this strategy. During the production of L-tryptophan by C. glutamicum, the accumulation of this product in the extracellular medium resulted in a backflow into the cells, which produced severe feedback inhibition in the biosynthesis of tryptophan.

19
Ikeda and Katsumata solved this problem by creating mutants with lower levels of tryptophan uptake, which resulted in an accumulation of 10 to 20% more tryptophan than in their parent. Another example of transport engineering is the increase in the fermentation yield of cysteine by overexpressing multidrug efflux genes (mar genes) in a cysteine-producing strain of E. coli.
 in a cysteine-producing strain of E. coli..")
Similar presentations





 2-phosphoglycerate. B) oxaloacetate. C)>")


 1. Why gluconeogenesis?>")


>")








