Download presentation
Presentation is loading. Please wait.

1
Key Points in Constitution of Cytoskeleton Network 1.Polymerization of monomer 1.Regulation of assembly and disassembly 1.Formation of network by associated protein 1.Perform function by motor proteins

2
Cytoskeleton functions are difficult to assay due to 1.Formation of large protein complex 2.Force generation without chemical change

3
Components of cytoskeleton

4
Microtubule is made up of 13 protofilaments

5
Tubulin dimers constitute protofilament

6
Nucleation of microtubule is GTP and Mg++- dependent

7
A threshold concentration of tubulin (critical concentration, Cc) Is indispensibe for microtubule elongation
 Is indispensibe for microtubule elongation")
8
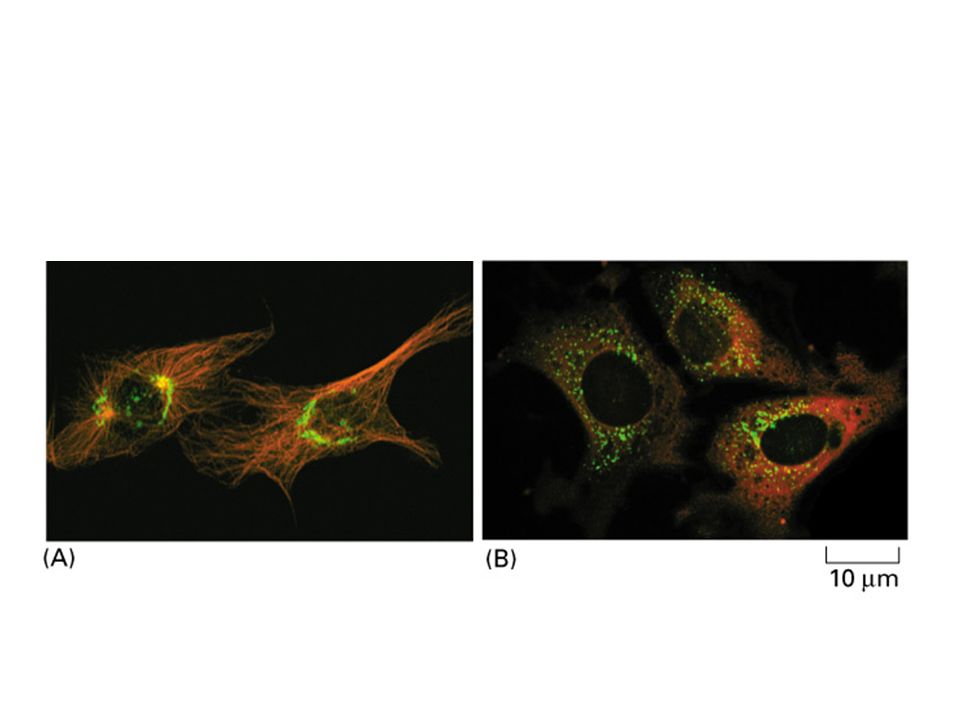
Flourescence– labeled microtubule demonstrates that microtubue is a dynamic structure

9
Dynamic instability of microtubule: consequence of delay hydrolysis of GTP after assembly

10
MTOC (microtubule organization center, centrosome)
")
11
Centrosome is the place within a cell where microtubule minus end is protected

12
Centrosome is located at peri-nuclear region

13
Centrosome is the center of a cell

14
Centrosome comprises centriole and pericentriole material

15
Structure of a centriole

16
Cilia: derivative of centrioles

17
-tubulin is the major component of pericentriole material

18
Microtubule dynamic can be manipulated experimentally

20
Microtubule associated proteins (MAP) enhance microtubule stability
 enhance microtubule stability")
21
Microtubules interact with other cytoskeleton via MAPs

22
Pigment granules move along the microtubule tracks

23
Dynein and kinesin are microtubule motors

24
Identification and characterization of microtubule motors

25
Dynein and kinesin move cargos along opposite direction

26
Molecular structure of kinesin

28
A model for moving microtubule cargo through motor proteins

29
Intracellular organelles distribute along microtubule networks Golgi ER MT

31
Intermediate filaments occur only in multicellular organisms

32
Intermediate filaments are the bona fide cellular skeleton

34
Intermdiate filament proteins are made up of helical rod domain and non-helical domain

35
Intermediate filament is a non-polar structure and its basic structural unit is the staggered tetramer of two coiled coil dimers

36
Assembly of Intermediate filaments (also called 10 nm filaments) is dependent on rod domain aggregation
 is dependent on rod domain aggregation")
37
Nuclear lamin disassembly is dependent on phosphorylation of non-helical domain

38
Intermediate filaments function as intercellular bridge to link neighboring cells via desmosomes

39
Keratins are divided into 2 groups based on their pI

40
Expression of keratin pairs are tissue specific

41
Keratins are more amenable to deforming force than other cytoskeletal proteins

42
Keratins are the first cytoskeletal protein found to be related to human diseases

43
Numerous hot spots are found in human keratin genes

44
Keratin mutation affects its assembly and mechanical property

45
Methods to generate transgenic mice

46
Transgenic mice overexpressing k14 mutant display Epidermolysis bullosa Simplex (EBS) phenotype
 phenotype")
47
Ultrastructure of epidermal cells displaying EBS

49
Actins occupy up to 5% total cellular protein

50
F actin is made up of G (globular) actins
 actins")
51
Polymerization of actin is ATP and K +, Mg 2+ - dependent

52
Amount of actin within a cell is kept dynamically equilibrium

53
S1 myosin fragment can be used to detect actin threadmilling

54
Barbed end (plus end) of actin contains newly decorated actin
 of actin contains newly decorated actin")
55
Profilin and thymosin compete with each other for actin monomer binding

57
Cytochalasin inhibits growth of actin fiber

58
Cellular structures made up of actin fibers

59
Cell cortex contains actin fibers arranged in different array and density

60
Stress fibers are not culture artifact

61
Actin binding proteins

62
-actinin and fimbrin are bundling proteins

63
Filamin is an actin-binding gel forming protein

64
Myosins are actin motor proteins Figure 16 – 51 Myosin II. (A) A myosin II molecule is composed of two heavy chains (each about 2000 amino acids long (green) and four light chains (blue). The light chains are of two distinct types, and one copy of each type is present on each myosin head. Dimerization occurs when the two a helices of the heavy chains wrap around each other to form a coiled- coil, driven by the association of regularly spaced hydrophobic amino acids (see Figure 3 – 11). The coiled-coil arrangement makes an extended rod in solution, and this part of the molecule is called the tail. (B) The two globular heads and the tail can be clearly seen in electron micrographs of myosin molecules shadowed with platinum. (B, courtesy of David Shotton.)
 A myosin II molecule is composed of two heavy chains (each about 2000 amino acids long (green) and four light chains (blue). The light chains are of two distinct types, and one copy of each type is present on each myosin head. Dimerization occurs when the two a helices of the heavy chains wrap around each other to form a coiled- coil, driven by the association of regularly spaced hydrophobic amino acids (see Figure 3 – 11). The coiled-coil arrangement makes an extended rod in solution, and this part of the molecule is called the tail. (B) The two globular heads and the tail can be clearly seen in electron micrographs of myosin molecules shadowed with platinum. (B, courtesy of David Shotton.).")
65
Mysoin derivatives

66
Actin moves along myosin

67
Myosin mutant mice displays defective heart Figure 16 – 75 Effect on the heart of a subtle mutation in cardiac myosin. Left, normal heart from a 6-day old mouse pup. Right, heart from a pup with a point mutation in both copies of its cardiac myosin gene, changing Arg 403 to Gln. Both atria are greatly enlarged (hypertrophic), and the mice die within a few weeks of birth. (From D. Fatkin et al., J. Clin. Invest. 103:147, 1999.)
, and the mice die within a few weeks of birth. (From D. Fatkin et al., J. Clin. Invest. 103:147, 1999.).")
68
Summary of actin binding proteins

72
Figure 16 – 50 The dramatic effects of Rac, Rho, and Cdc42 on actin organization in fibroblasts. In each case, the actin filaments have been labeled with fluorescent phalloidin, and focal contacts have been located with an antibody against vinculin. (A) Serum-starved fibroblasts have actin filaments primarily in the cortex, and relatively few focal contacts. (B) Microinjection of a constitutively activated form of Rho causes the rapid assembly of many prominent stress fibers and focal contacts. (C) Microinjection of a constitutively activated form of Rac, a closely related monomeric GTPase, causes the formation of an enormous lamellipodium that extends from the entire circumference of the cell. (D) Microinjection of a constitutively activated form of Cdc42, another Rho family member, causes the protrusion of many long filopodia at the cell periphery that form adhesive contacts with the substratum. The distinct global effects of these three GTPases on the organization of the actin cytoskeleton are mediated by the actions of dozens of other protein molecules that are regulated by the GTPases. Many of these target proteins resemble the various actin-associated proteins that we have discussed in this chapter. (From A. Hall, Science 279:509 – 514, 1998. © AAAS.)
 Serum-starved fibroblasts have actin filaments primarily in the cortex, and relatively few focal contacts. (B) Microinjection of a constitutively activated form of Rho causes the rapid assembly of many prominent stress fibers and focal contacts. (C) Microinjection of a constitutively activated form of Rac, a closely related monomeric GTPase, causes the formation of an enormous lamellipodium that extends from the entire circumference of the cell. (D) Microinjection of a constitutively activated form of Cdc42, another Rho family member, causes the protrusion of many long filopodia at the cell periphery that form adhesive contacts with the substratum. The distinct global effects of these three GTPases on the organization of the actin cytoskeleton are mediated by the actions of dozens of other protein molecules that are regulated by the GTPases. Many of these target proteins resemble the various actin-associated proteins that we have discussed in this chapter. (From A. Hall, Science 279:509 – 514, © AAAS.).")
75
Figure 16 – 90 A model for protrusion of the actin meshwork at the leading edge. Two time points during advance of the lamellipodium are illustrated, with newly assembled structures at the later time point shown in a lighter color. Nucleation is mediated by the ARP complex at the front. Newly nucleated actin filaments are attached to the sides of preexisting filaments, primarily at a 70° angle. Filaments elongate, pushing the plasma membrane forward because of some sort of anchorage of the array behind. At a steady rate, actin filament plus ends become capped. After newly polymerized actin subunits hydrolyze their bound ATP in the filament lattice, the filaments become susceptible to depolymerization by cofilin. This cycle causes a spatial separation between net filament assembly at the front and net filament disassembly at the rear, so that the actin filament network as a whole can move forward, even though the individual filaments within it remain stationary with respect to the substratum.

Similar presentations
















, intermediate.>")






 Actin-binding proteins affect the localized assembly or disassembly of the actin.>")



 * Cancer diseases therapy: taxanes.>")

