Download presentation
Presentation is loading. Please wait.

1
Evolutionary Processes
25

2
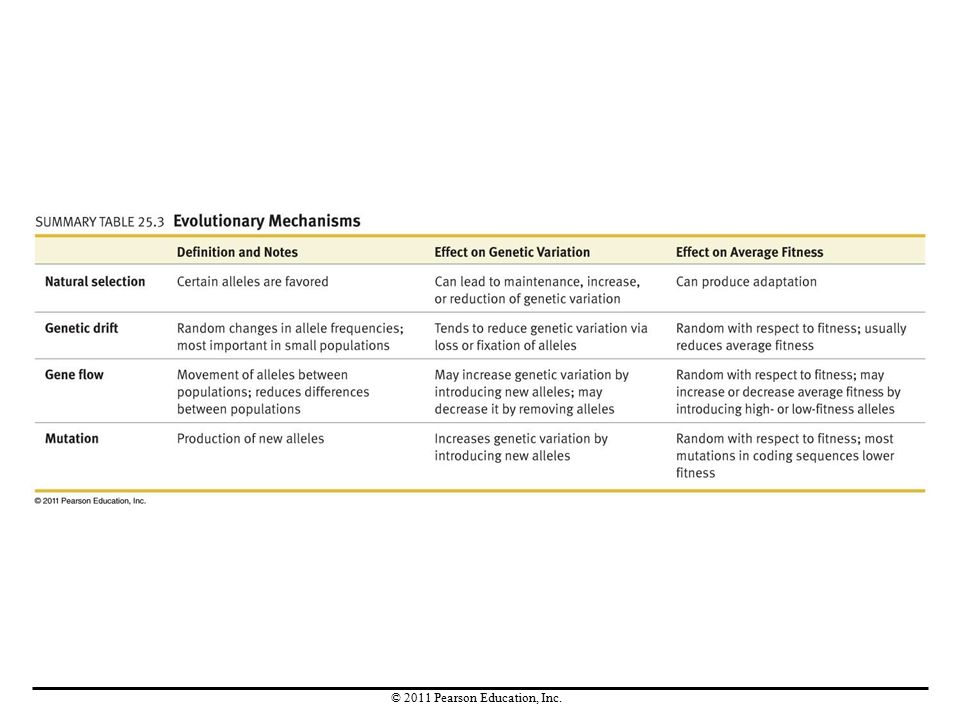
Key Concepts The Hardy-Weinberg principle acts as a null hypothesis when researchers want to test whether evolution or nonrandom mating is occurring at a particular gene. Each of the four evolutionary mechanisms has different consequences. Only natural selection produces adaptation. Genetic drift causes random fluctuations in allele frequencies. Gene flow equalizes allele frequencies between populations. Mutation introduces new alleles.

3
Key Concepts Inbreeding changes genotype frequencies but does not change allele frequencies. Sexual selection leads to the evolution of traits that help individuals attract mates. It is usually stronger on males than on females.

4
Introduction There are four mechanisms that shift allele frequencies in populations: Natural selection increases the frequency of those alleles that contribute to reproductive success in a particular environment. Genetic drift causes allele frequencies to change randomly. Gene flow occurs when individuals leave one population, join another, and breed. Mutation modifies allele frequencies by continually introducing new alleles.

5
Analyzing Change in Allele Frequencies
To study how the four evolutionary processes affect populations, biologists take a three-pronged approach: They create mathematical models that predict the fate of alleles over time under various conditions. They collect data to test predictions made by the models. They apply the results to solve problems in human genetics, conservation of endangered species, or other fields.

6
The Hardy-Weinberg Principle
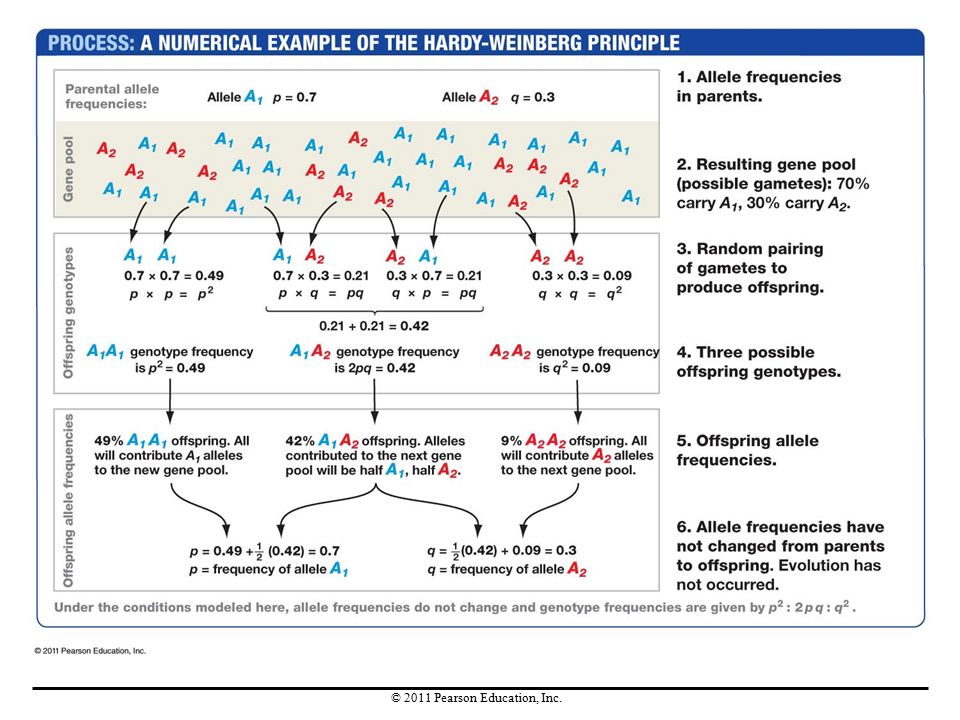
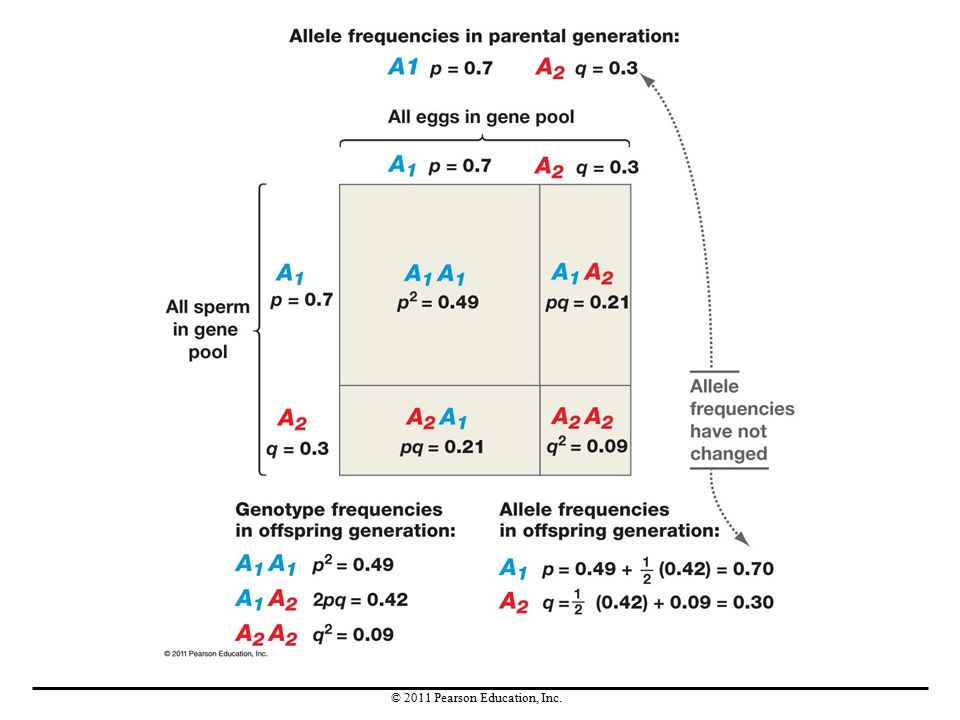
To study how the four evolutionary processes affect populations, in 1908 G. H. Hardy and Wilhelm Weinberg developed a mathematical model to analyze the consequences of matings among all of the individuals in a population. Hardy and Weinberg wanted to know what happened in an entire population, when all of the individuals—and thus all possible genotypes—bred.

7
The Gene Pool Concept Hardy and Weinberg imagined that all of the gametes produced in each generation go into a single group called a gene pool and then combine randomly. Their calculations predict the genotypes of the offspring that the population would produce, as well as the frequency of each genotype.

8
Deriving the Hardy-Weinberg Principle
They started with the simplest situation, a gene with two alleles, A1 and A2. The frequency of A1 is represented by p and the frequency of A2 is represented by q. Because there are only two alleles, p + q = 1. In this situation, three genotypes are possible: A1A1, A1A2, and A2A2. The model predicts the following genotype frequencies: The frequency of the A1A1 genotype is p2. The frequency of the A2A2 genotype is q2. The frequency of the A1A2 genotype is 2pq.

9
Deriving the Hardy-Weinberg Principle
Because all individuals in the new generation must have one of the three genotypes, the sum of the three genotype frequencies must equal 1 (100% of the population): p2 + 2pq + q2 = 1. This is the Hardy-Weinberg equation. When allele frequencies are calculated for this new generation, the frequency of A1 is still p and the frequency of A2 is still q.
: p2 + 2pq + q2 = 1. This is the Hardy-Weinberg equation. When allele frequencies are calculated for this new generation, the frequency of A1 is still p and the frequency of A2 is still q.")
11
The Hardy-Weinberg Principle
The Hardy-Weinberg principle makes two fundamental claims: If the frequencies of alleles A1 and A2 in a population are given by p and q, then the frequencies of genotypes A1A1, A1A2, and A2A2 will be given by p2, 2pq, and q2 for generation after generation. When alleles are transmitted via meiosis and random combination of gametes, their frequencies do not change over time. For evolution to occur, some other factor or factors must come into play.

13
Important Assumptions of the Hardy-Weinberg Model
For a population to conform to the Hardy-Weinberg principle, none of the four mechanisms of evolution can be acting on the population. In addition, the model assumes that mating is random with respect to the gene in question. Thus, here are the five assumptions that must be met: No natural selection. No genetic drift or random allele frequency changes. No gene flow. No mutation. Random mating.

14
Hardy-Weinberg Principle: A Null Hypothesis
Biologists often want to test whether natural selection is acting on a particular gene, nonrandom mating is occurring, or one of the other evolutionary mechanisms is at work. In addressing questions like these, the Hardy-Weinberg principle functions as a null hypothesis. When genotype frequencies do not conform to Hardy-Weinberg proportions, evolution or nonrandom mating is occurring in that population.

15
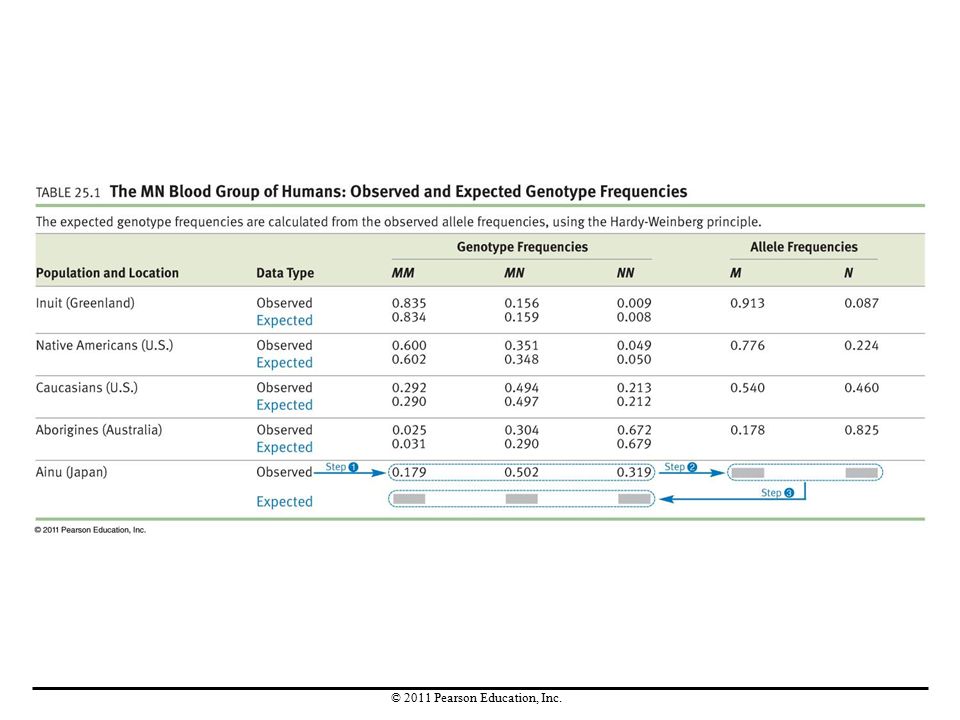
Case Study 1: MN Blood Types in Humans
Most human populations have two alleles for the MN blood group. The genotype of a person can be determined from blood samples. Analysis to determine if the Hardy-Weinberg principle holds requires four steps: Estimate genotype frequencies. Calculate observed allele frequencies from the observed genotype frequencies. Use the observed allele frequencies to calculate the genotypes expected according to the Hardy-Weinberg principle. Compare the observed and expected values.

17
Are MN Blood Types in Equilibrium?
The observed and expected MN genotype frequencies were almost identical. Since the genotypes at the MN locus are in Hardy-Weinberg proportions, evolutionary processes do not currently affect MN blood groups, and mating must be random with respect to this trait. The Hardy-Weinberg principle is used to test the hypothesis that currently no evolution is occurring at a particular gene and that in the previous generation, mating was random with respect to the gene in question.

18
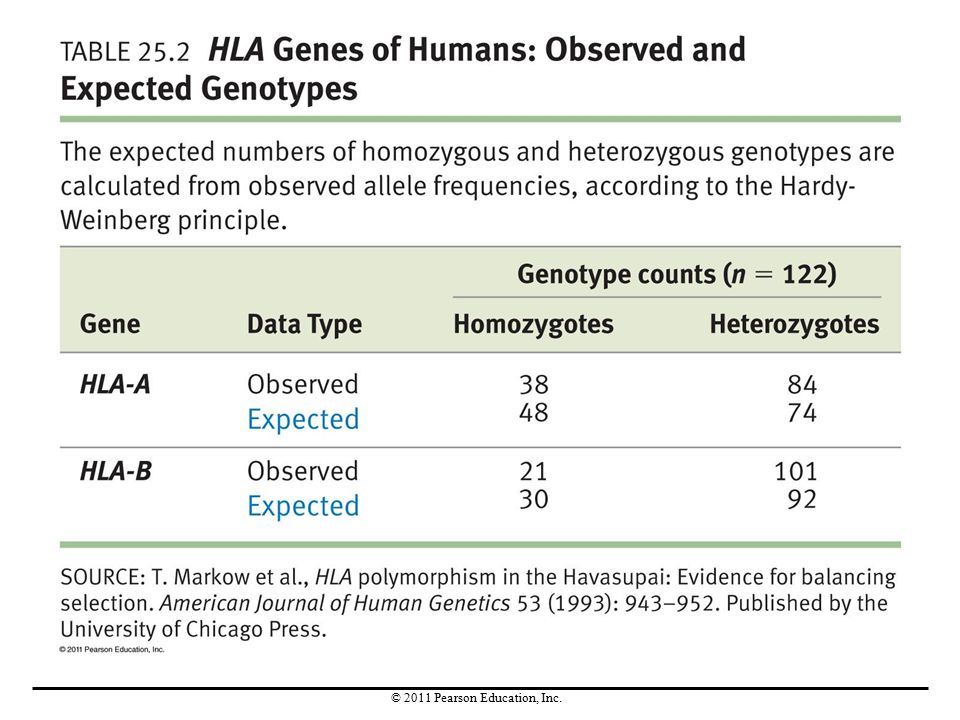
Case Study 2: HLA Genes in Humans
The HLA genes code for proteins that are important in the function of the immune system. To test the hypothesis that heterozygotes for the HLA-A and HLA-B genes might be more fit than homozygotes, researchers used the genotypes of 125 Havasupai tribe members to estimate population allele frequencies.

20
Are HLA Genes in Equilibrium?
The expected genotype frequencies did not match the observed frequencies. Therefore, at least one of the Hardy-Weinberg assumptions must be violated for these alleles in this population. Mutation, migration, and genetic drift are negligible in this case. There are two possible explanations for this result: Mating is not random with respect to the HLA genotype. Heterozygous individuals have higher fitness. Research continues on this, with no answer apparent yet.

21
The Hardy-Weinberg Principle

22
Types of Natural Selection
Natural selection occurs in a wide variety of patterns. Genetic variation refers to the number and relative frequency of alleles that are present in a particular population. Maintaining genetic variation is important because lack of variation can make populations less able to respond successfully to changes in the environment.

23
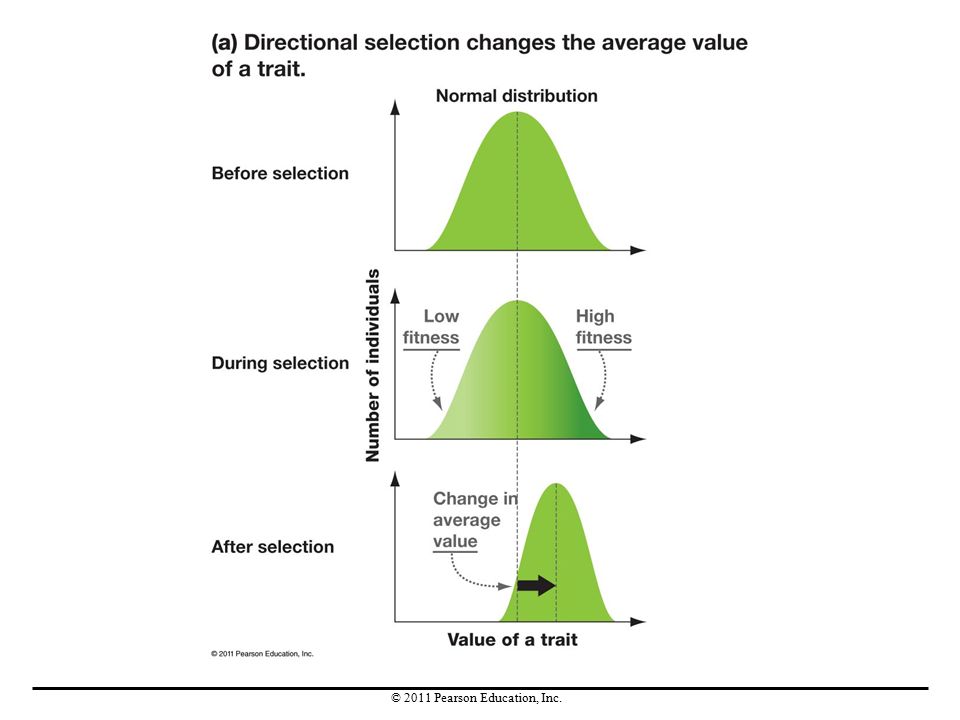
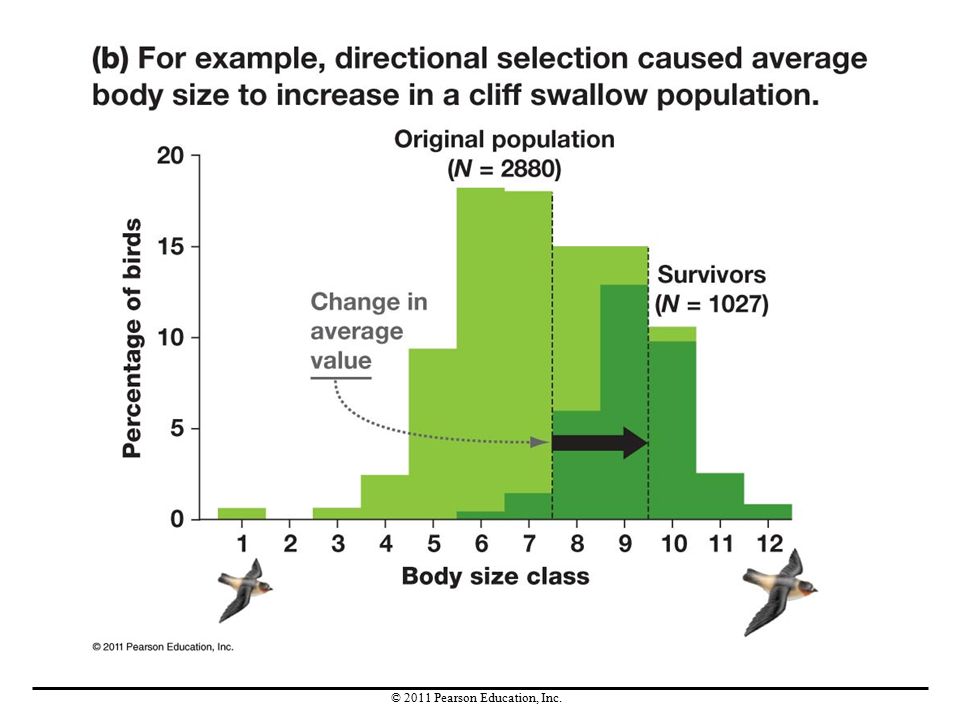
Directional Selection
Directional selection is a pattern of natural selection that increases the frequency of one allele. This type of selection reduces a population’s genetic diversity over time. If directional selection continues over time, the favored alleles eventually become fixed, reaching a frequency of 1.0, or 100%. Disadvantageous alleles will be lost, reaching a frequency of 0.0. When disadvantageous alleles decline in frequency, purifying selection is said to occur.

27
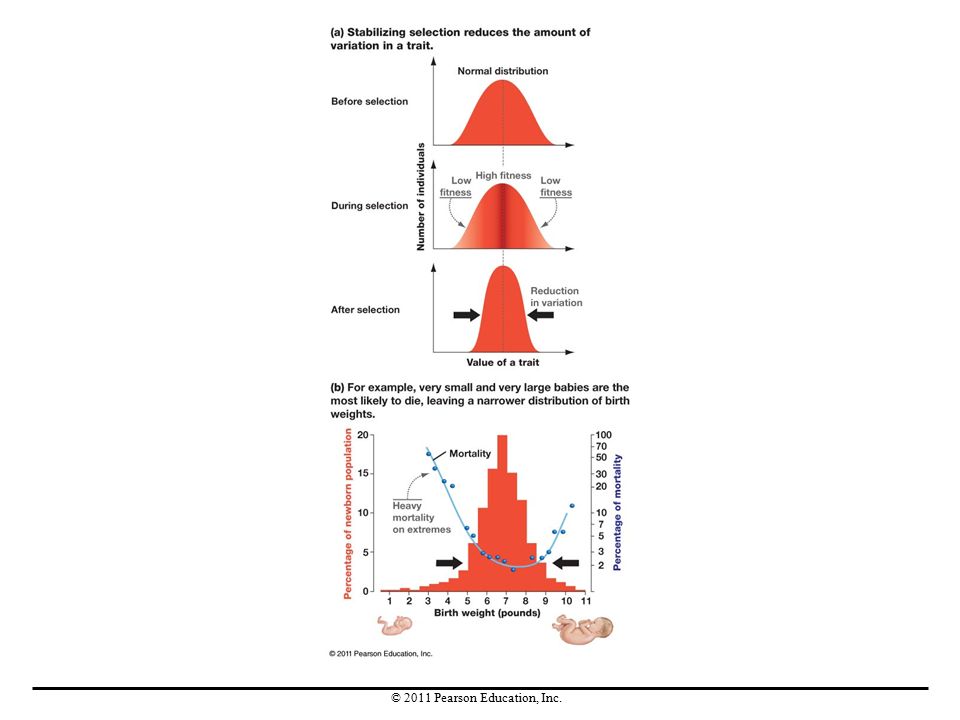
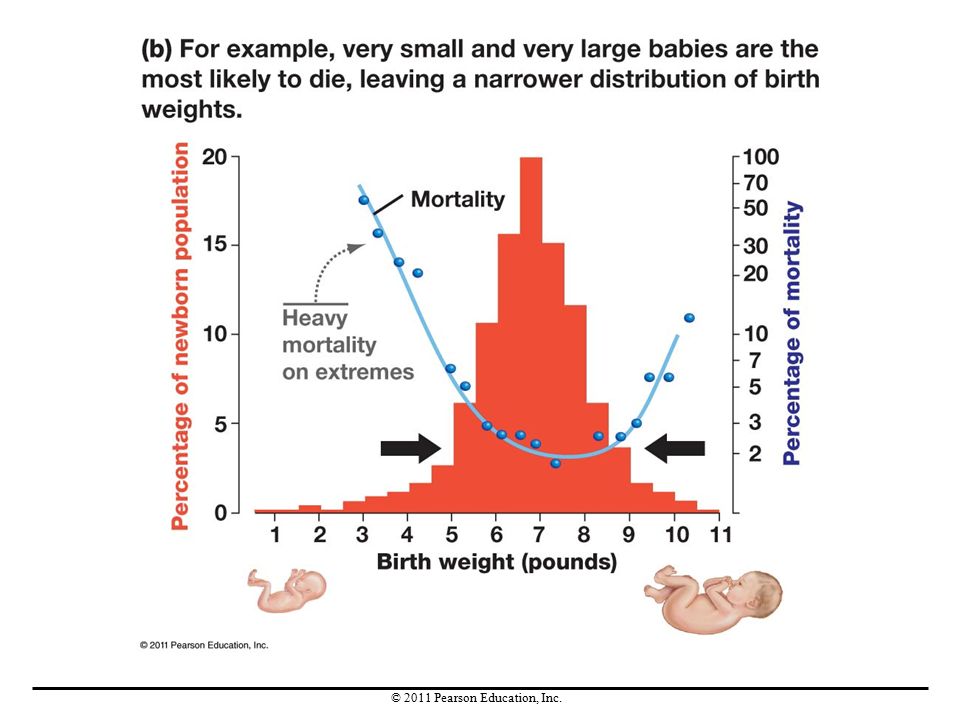
Stabilizing Selection
Stabilizing selection occurs when individuals with intermediate traits reproduce more than others, thereby maintaining intermediate phenotypes in a population. There is no change in the average value of a trait over time, and genetic variation in the population is reduced. An example is the percentage of newborn humans with various birth weights compared with their mortality rates. Those with birth weights in the middle of the range were most likely to survive.

31
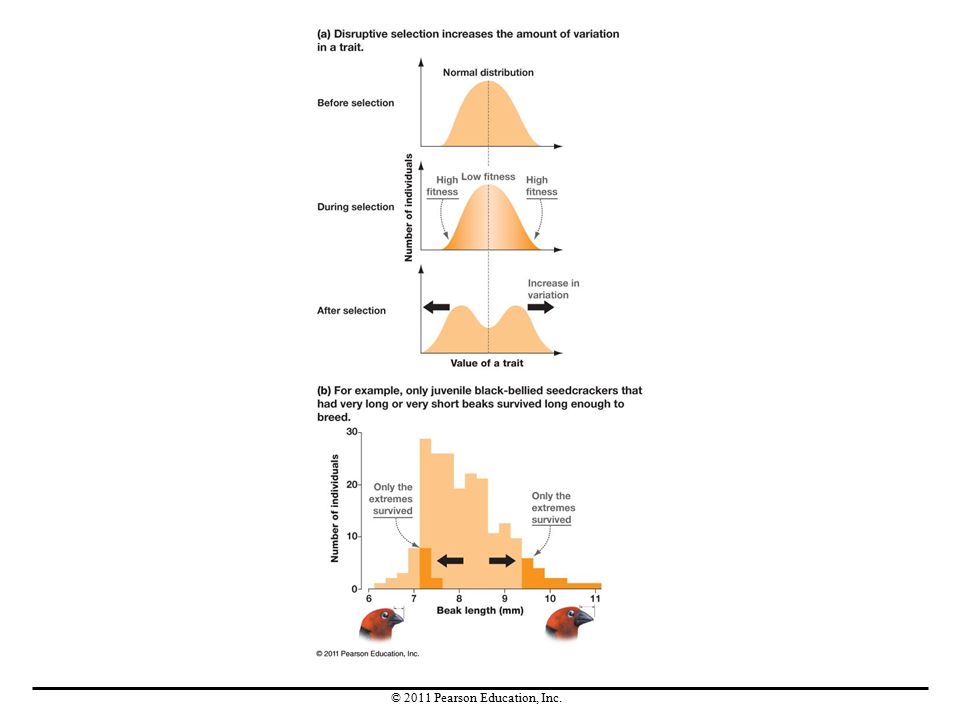
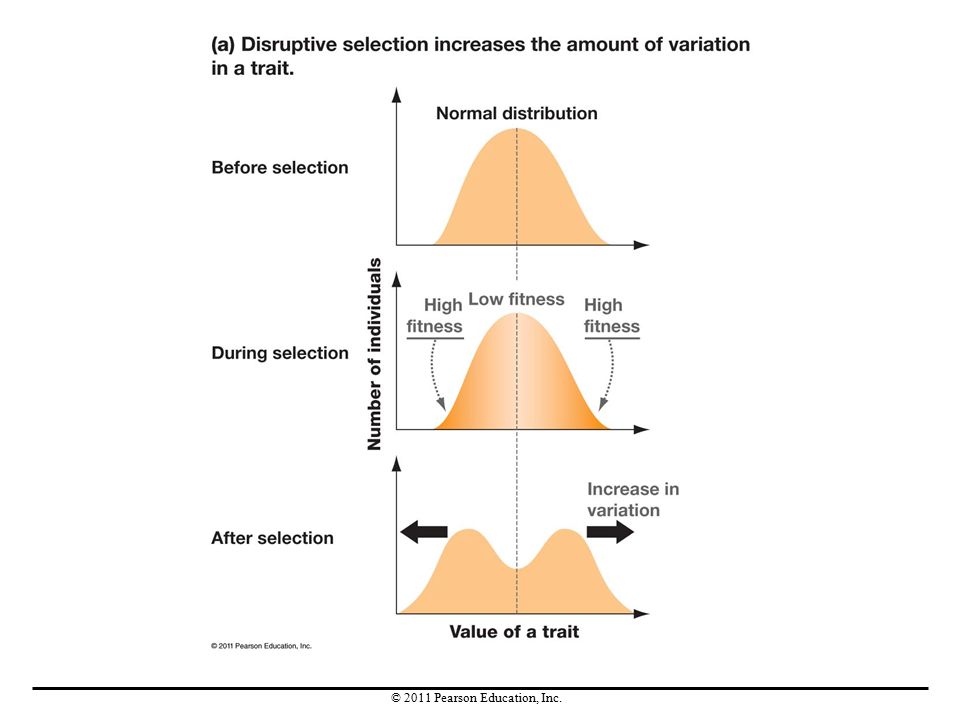
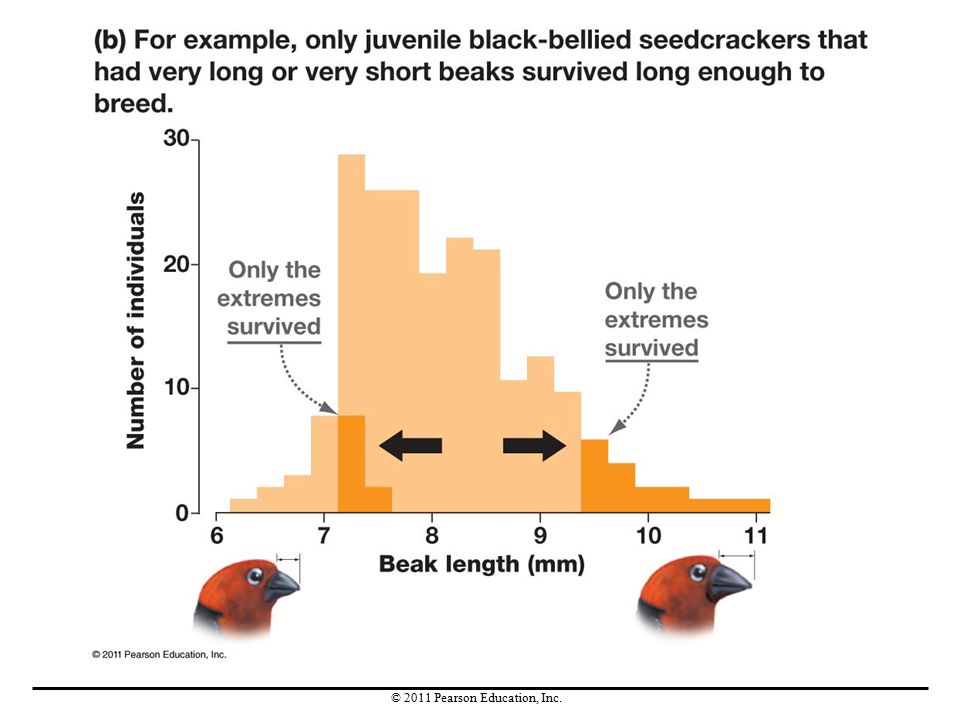
Disruptive Selection In contrast to stabilizing selection, the pattern of natural selection called disruptive selection occurs when intermediate phenotypes are selected against and extreme phenotypes are favored. Disruptive selection maintains genetic variation but does not change the mean value of a trait. Disruptive selection can cause speciation, the formation of new species, if individuals with one extreme of a trait start mating preferentially with individuals that have the same trait.

35
Heterozygote Advantage
Directional selection, stabilizing selection, and disruptive selection describe how natural selection can act on polygenic traits in a single generation or episode. However, they are not the only patterns of selection. In heterozygote advantage, heterozygous individuals have higher fitness than homozygous individuals do, thus maintaining genetic variation in the population. Heterozygote advantage is one mechanism responsible for a balancing selection, in which no single allele has a distinct advantage.

36
Balancing Selection Balancing selection also occurs when:
The environment varies over time or in different geographic areas such that certain alleles are favored by natural selection at different times or in different places. This results in the maintenance of or increase in overall genetic variation. Certain alleles are favored when they are rare, but not when they are common—a pattern known as frequency-dependent selection. As a result, overall genetic variation in the population is maintained or increased.

37
Three Modes of Natural Selection

38
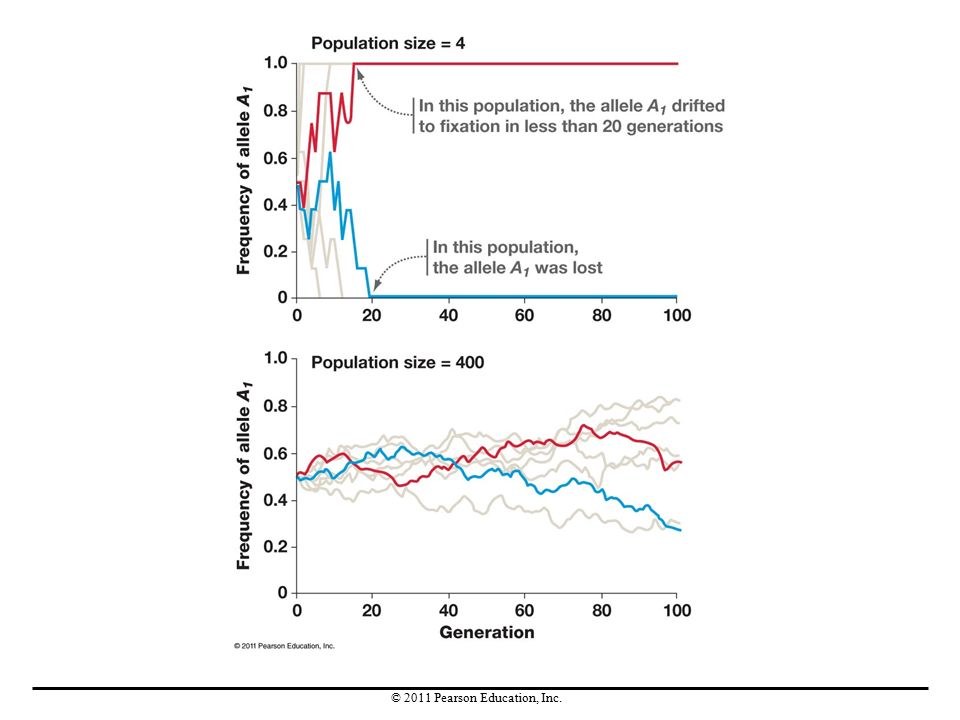
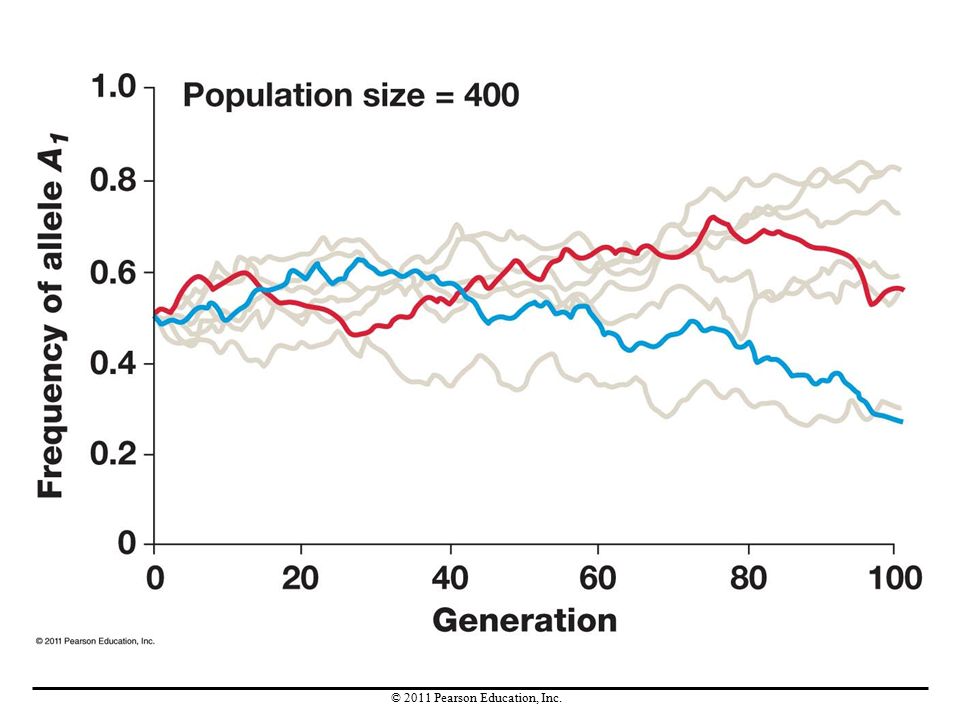
Genetic Drift Genetic drift is any change in allele frequencies in a population due to chance (sampling error). Drift occurs in every population, in every generation. It causes allele frequencies to drift up and down randomly over time.
. Drift occurs in every population, in every generation. It causes allele frequencies to drift up and down randomly over time.")
39
Key Points about Genetic Drift
Genetic drift is random with respect to fitness. These allele frequency changes are not adaptive. Over time, genetic drift can lead to the random loss or fixation of alleles. When random loss or fixation occurs, genetic variation in the population declines. Genetic drift is more pronounced in small populations than in large ones. Given enough time, however, genetic drift can be an important factor even in large populations.

43
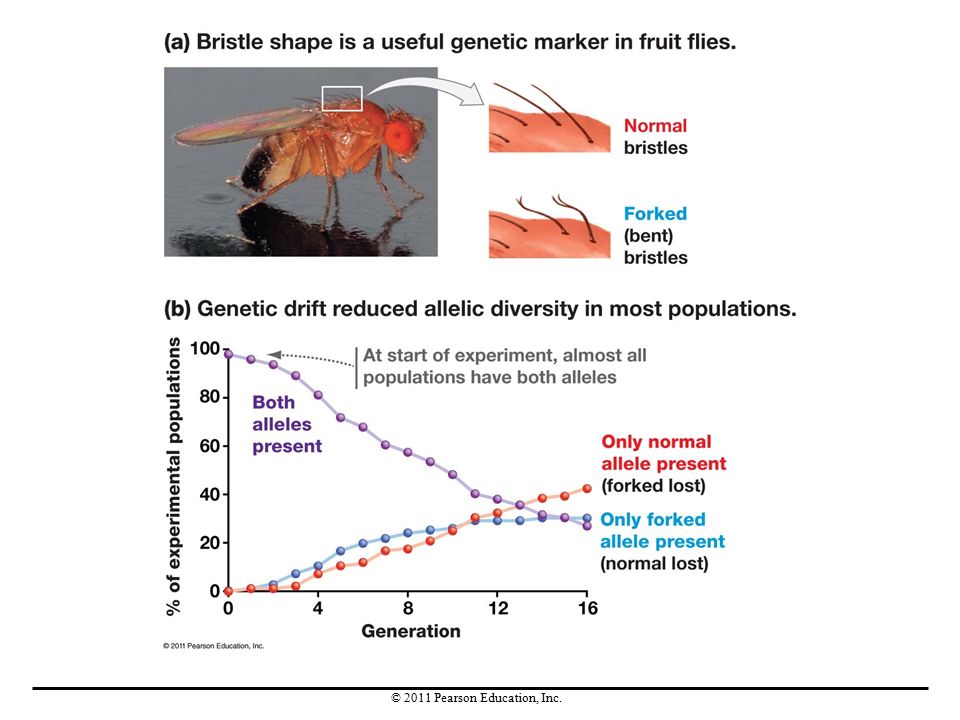
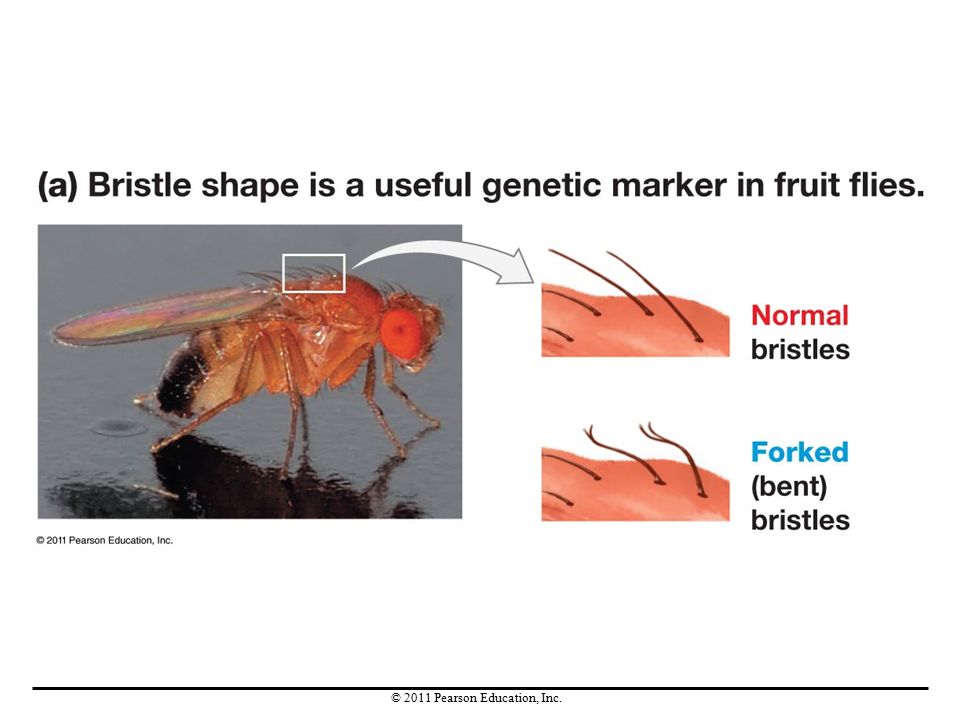
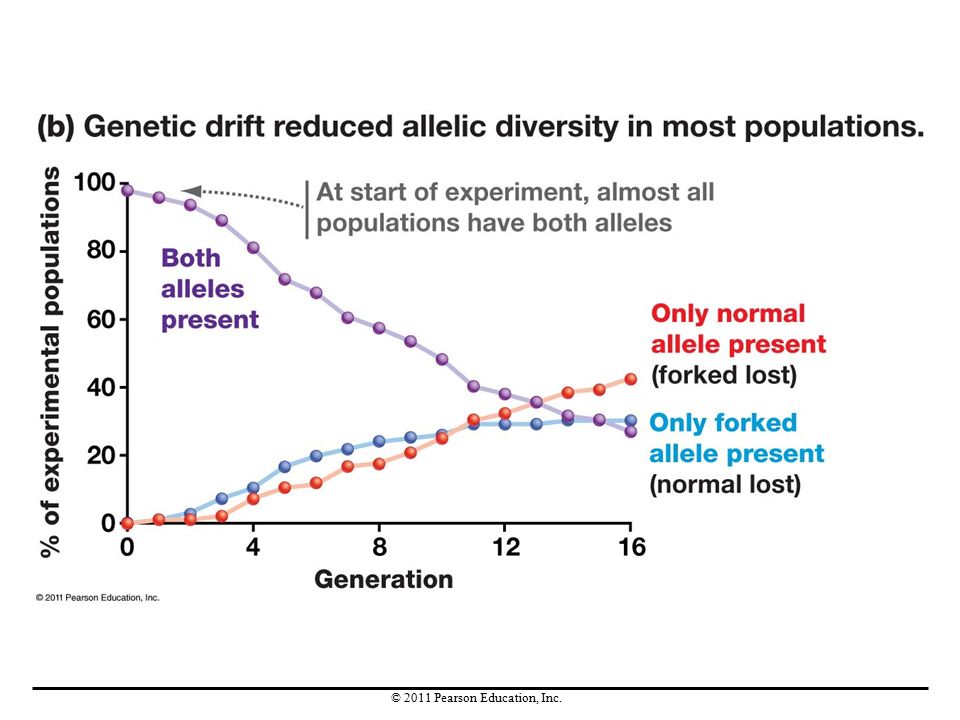
Experimental Studies of Genetic Drift
Research on genetic drift in small populations of fruit flies used a genetic marker—a specific allele that causes a distinctive phenotype—for leg-bristle morphology. This gene has two alleles, one resulting in straight (wild-type) leg bristles and the other in forked (bent) bristles. In 70 of the 96 populations studied, genetic drift caused one allele to be lost. In the laboratory, genetic drift was found to decrease genetic variation within populations and increase genetic differences between populations.
 leg bristles and the other in forked (bent) bristles. In 70 of the 96 populations studied, genetic drift caused one allele to be lost. In the laboratory, genetic drift was found to decrease genetic variation within populations and increase genetic differences between populations.")
47
Genetic Drift in Natural Populations
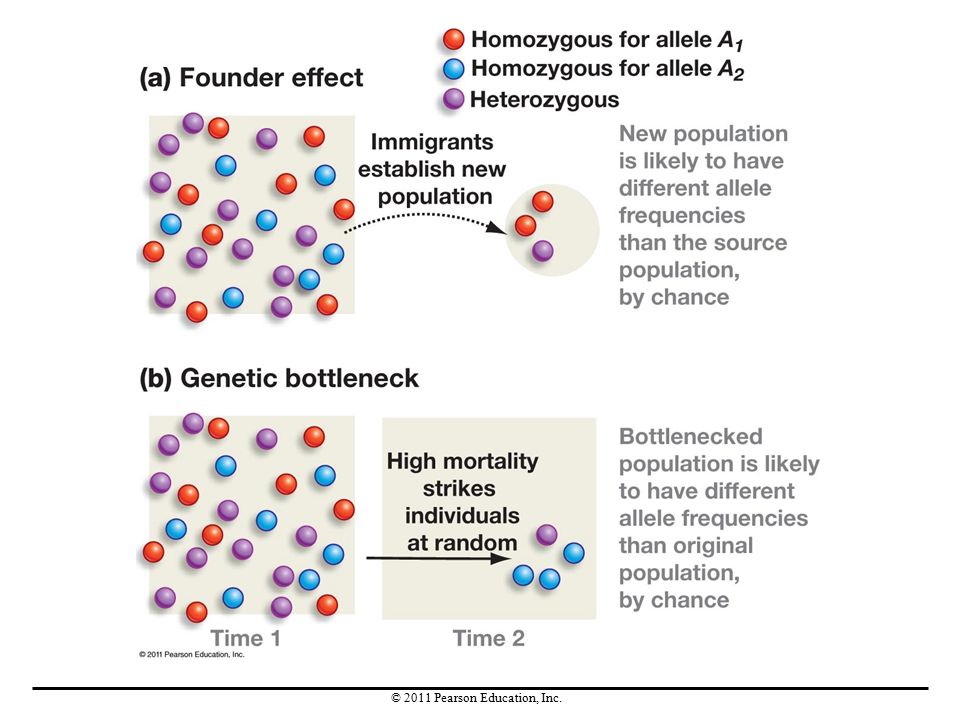
Genetic drift is of great concern to conservation biologists because the small populations found on nature reserves or in zoos are especially susceptible to it. Genetic drift can be caused by any event or process that involves sampling error, not just the sampling of gametes that occurs during fertilization. Two examples are the founder effect and bottlenecks.

48
Founder Effects Cause Genetic Drift
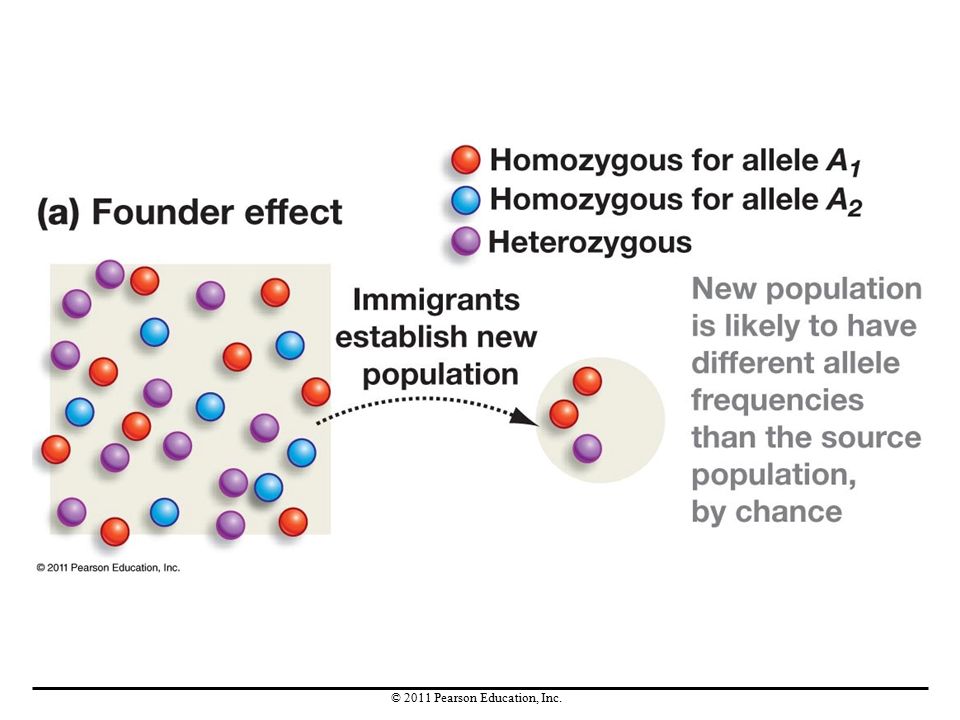
A founder event occurs when a group starts a new population in a new area. Especially in small groups, allele frequencies probably differ from the source population; this change in allele frequencies is called a founder effect. Founder effects are especially common in the colonization of isolated habitats. Each time a founder event occurs, a founder effect is likely to accompany it, changing allele frequencies through genetic drift.

49
Population Bottlenecks Cause Genetic Drift
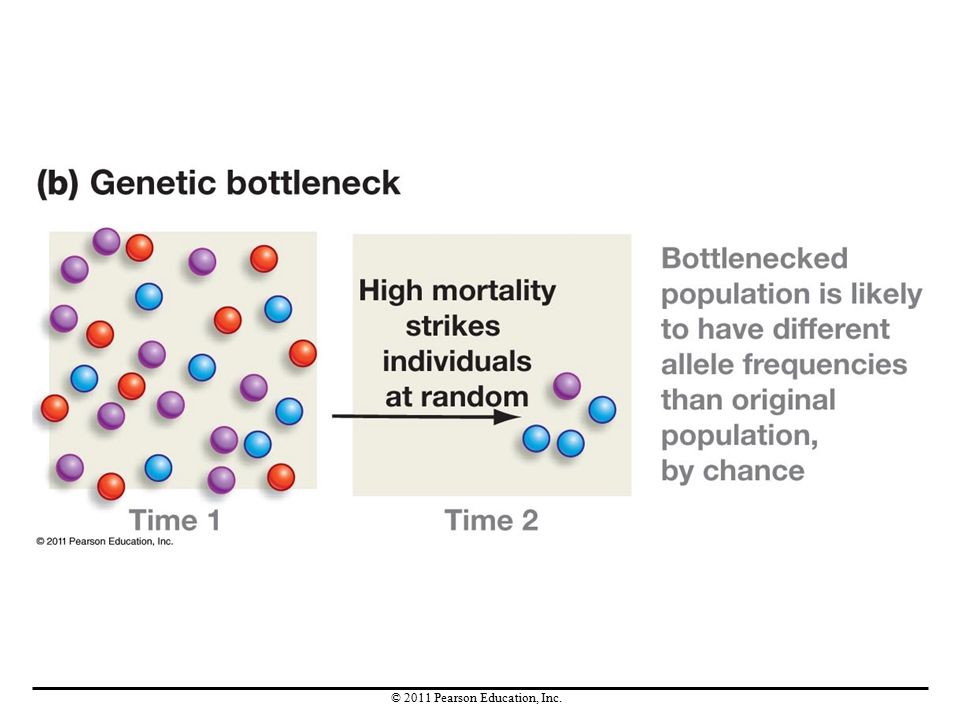
A sudden decrease in population size is called a population bottleneck. Population bottlenecks are commonly caused by disease outbreaks and natural catastrophes. Population bottlenecks lead to genetic bottlenecks—a sudden reduction in the number of alleles in a population. Drift occurs during genetic bottlenecks and causes a change in allele frequencies.

53
Gene Flow Gene flow, the movement of alleles from one population to another, occurs whenever individuals leave one population, join another, and breed. Gene flow equalizes gene frequencies between the source and recipient populations. In other words, gene flow homogenizes populations. Gene flow is random with respect to fitness, but movement of alleles between populations always tends to reduce genetic differences between them.

55
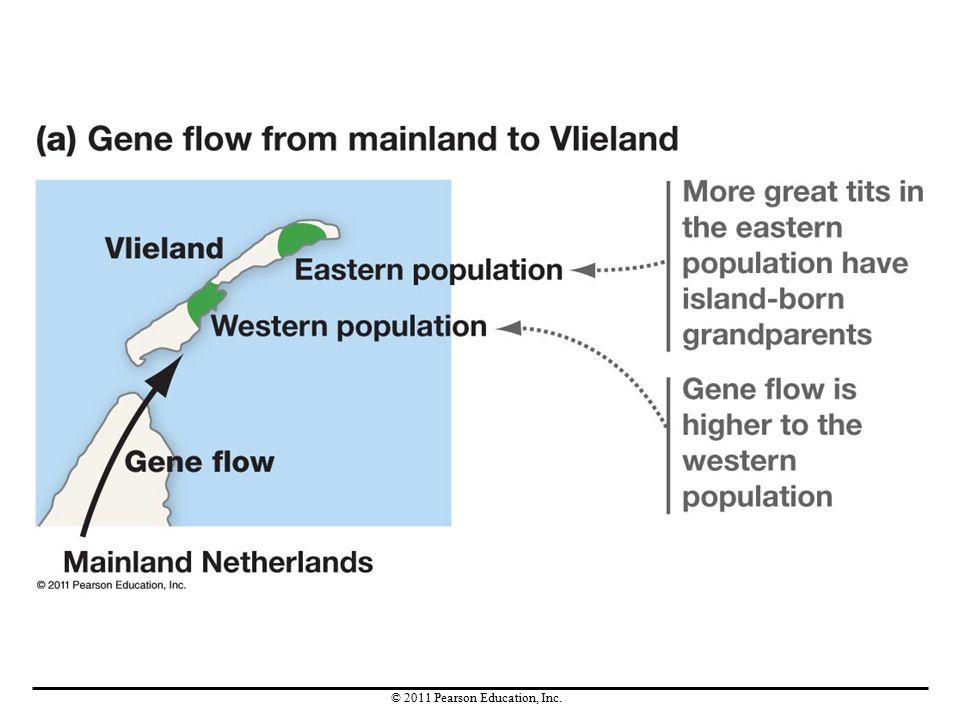
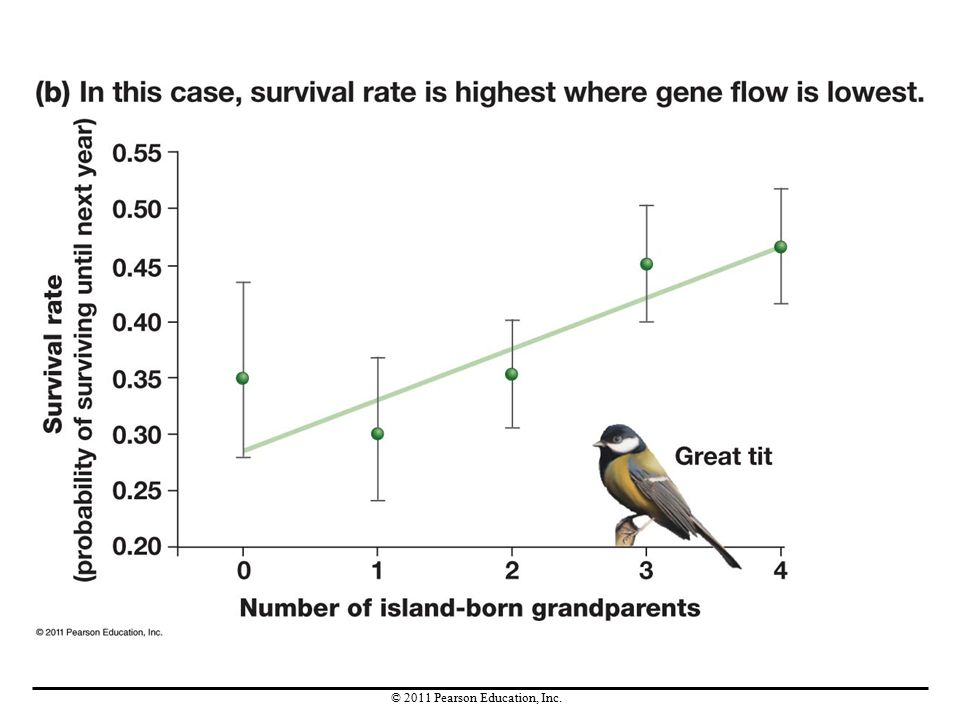
Gene Flow in Natural Populations
On the island of Vlieland off the coast of the Netherlands, birds called great tits breed in two sets of woodlands. Females hatched in the eastern woodland appear to be more well adapted. Gene flow is higher in the western woodland; thus birds coming from the mainland are introducing alleles at a higher rate into the western population. These mainland alleles result in individuals less well-adapted to the island environment.

59
Mutation Although most evolutionary mechanisms reduce genetic diversity, mutation restores genetic diversity by creating new alleles. Mutation is random with respect to the fitness of the affected allele. Because mutations cause random changes in genes, most of them result in deleterious alleles, alleles that lower fitness. These alleles tend to be eliminated by selection. Rarely, mutation produces a beneficial allele that should increase in frequency in a population due to natural selection.

60
Mutation as an Evolutionary Mechanism
Mutation can be a significant evolutionary force in bacteria and archaea, which have short generation times. However, mutation in eukaryotes rarely causes a change from the genotype frequencies expected under the Hardy-Weinberg principle. As an evolutionary mechanism, mutation is relatively slow compared with natural selection, genetic drift, and gene flow. Mutation introduces new alleles into every individual in every population in every generation.

61
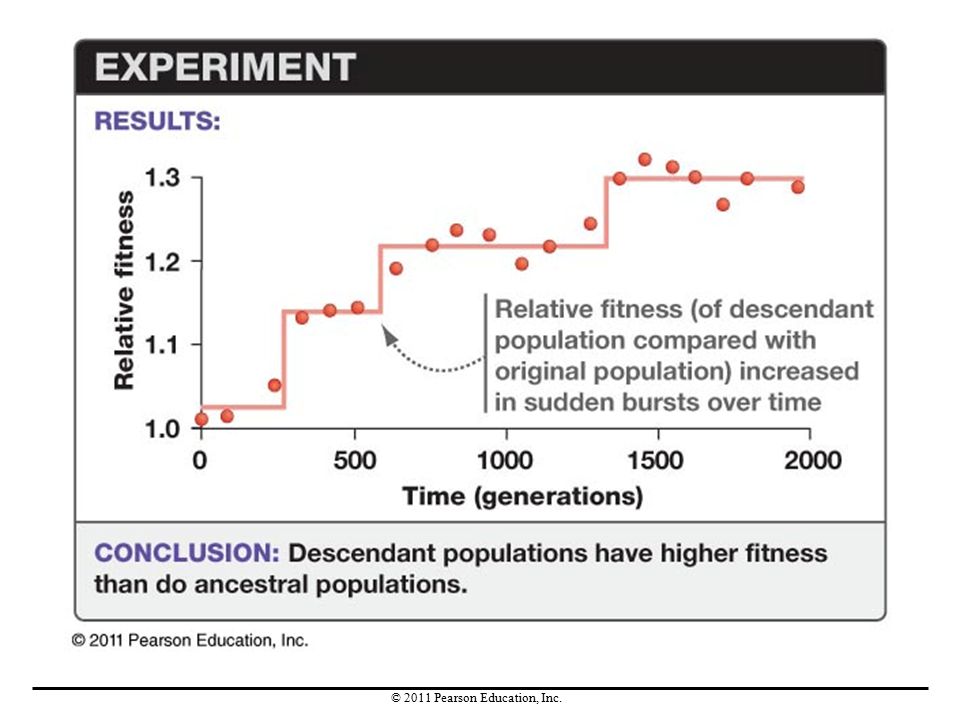
Experimental Studies of Mutation
The bacterium Escherichia coli has been used as a model to study how mutation affects evolution. In one experiment, Richard Lenski and colleagues set up populations of E. coli and followed them for 10,000 generations. E. coli is asexual, so mutation is its only source of genetic variation. Although no gene flow occurred, both selection and genetic drift operated in each population.

62
Experimental Studies of Mutation
The researchers found that the relative fitness of the populations increased over time in jumps. They suggested that this pattern resulted from novel mutations arising and conferring a fitness benefit.

67
Take-Home Messages Mutation plays an important role in evolution:
Mutation is the ultimate source of genetic variation. Crossing over and independent assortment shuffle existing alleles into new combinations, but only mutation creates new alleles. Without mutation, evolution would eventually stop. Mutation alone is usually inconsequential in changing allele frequencies at a particular gene. Each of the four evolutionary forces has different consequences for allele frequencies.

69
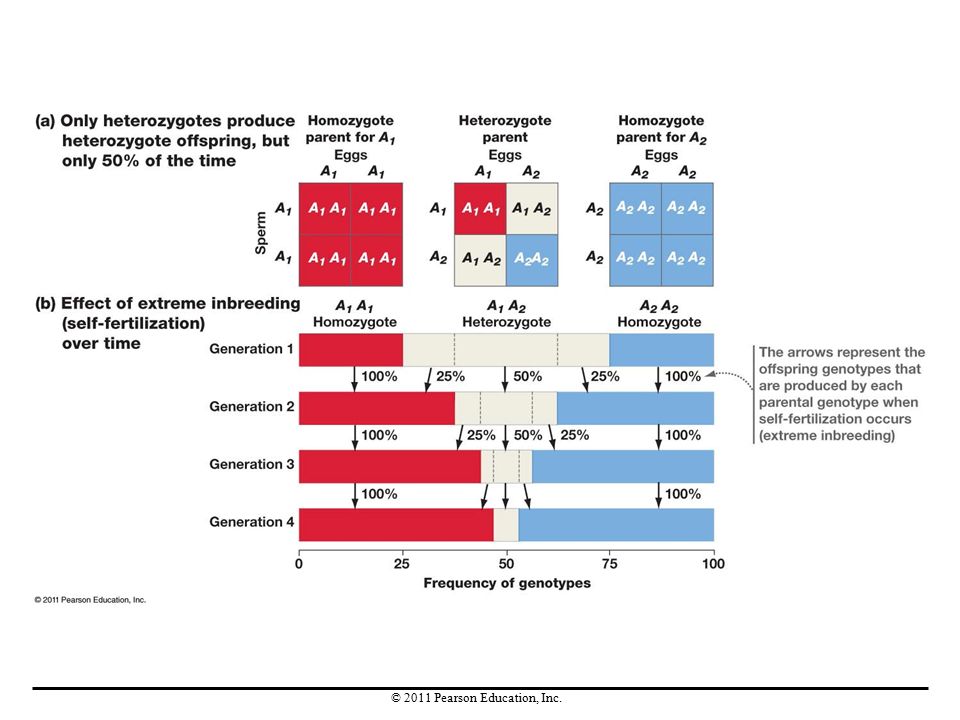
Nonrandom Mating In nature, mating may not be random with respect to any particular gene in question. Two mechanisms that violate the Hardy-Weinberg assumption of random mating are inbreeding and sexual selection.

70
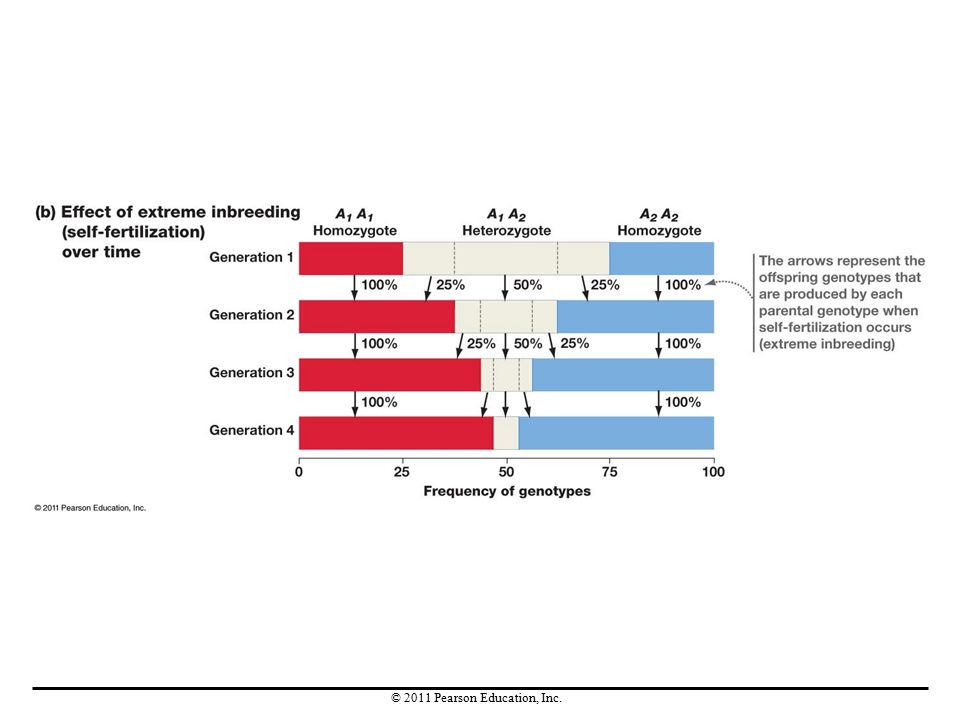
Inbreeding Inbreeding (mating between relatives) increases the frequency of homozygotes and reduces the frequency of heterozygotes in each generation. Inbreeding does not cause evolution, because allele frequencies do not change in the population as a whole. Inbreeding and other forms of nonrandom mating change genotype frequencies—not allele frequencies. Inbreeding depression is a decline in average fitness that takes place when homozygosity increases and heterozygosity decreases in a population.
 increases the frequency of homozygotes and reduces the frequency of heterozygotes in each generation. Inbreeding does not cause evolution, because allele frequencies do not change in the population as a whole. Inbreeding and other forms of nonrandom mating change genotype frequencies—not allele frequencies. Inbreeding depression is a decline in average fitness that takes place when homozygosity increases and heterozygosity decreases in a population.")
74
Inbreeding Depression
Inbreeding depression results from two processes: Many recessive alleles represent loss-of-function mutations. In heterozygotes these alleles have little or no effect; but inbreeding increases the frequency of homozygous recessive individuals and thus the frequency of individuals expressing the mutation. Many genes—especially those involved in fighting disease—are under intense selection for heterozygote advantage. If an individual is homozygous at these genes, then fitness declines.

76
Inbreeding Even though it does not cause evolution directly—because it does not change allele frequencies—inbreeding can speed the rate of evolutionary change. More specifically, it increases the rate at which purifying selection eliminates recessive deleterious alleles from a population.

77
Sexual Selection Sexual selection occurs when individuals within a population differ in their ability to attract mates. It favors individuals with heritable traits that enhance their ability to obtain mates. Sexual selection is a special form of natural selection.

78
The Fundamental Asymmetry of Sex
The fundamental asymmetry of sex results from the fact that, in most species, females usually invest more in their offspring than males do. There are two broad types of sexual selection: female choice and male-male competition. In other words, females should be choosy about their mates, while males will have to compete with each other for mates. Sexual selection should act more strongly on males than on females.

79
Female Choice for “Good Alleles”
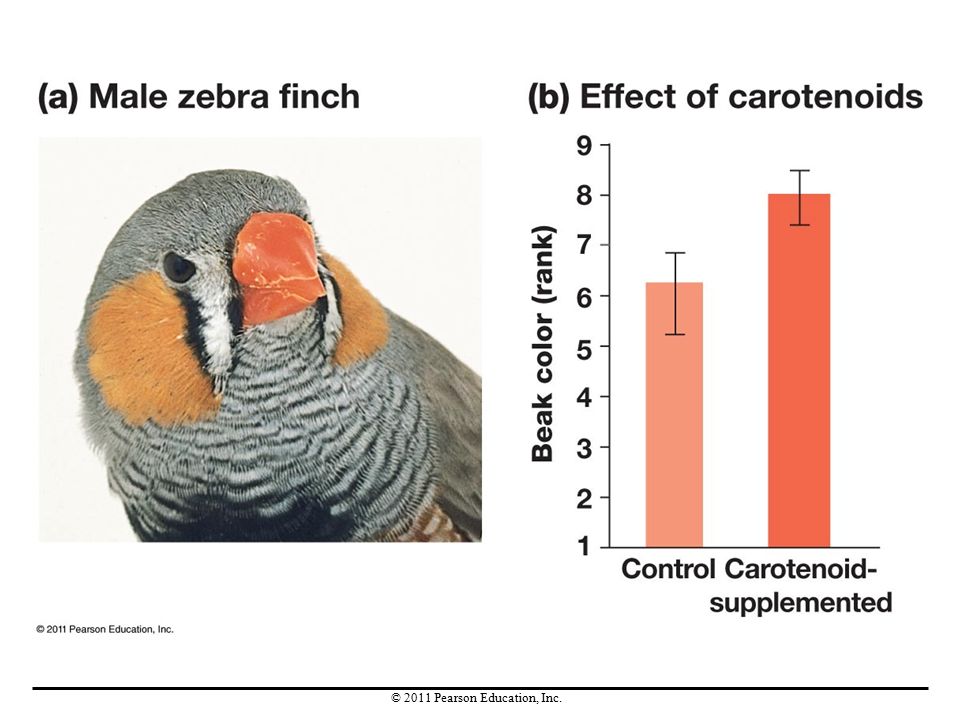
Females may choose mates on the basis of physical characteristics that signal male genetic quality. For example, research has shown that female zebra finches preferred males with more colorful beaks and feathers (indicating better health).
.")
83
Female Choice for Paternal Care
In many species, females prefer to mate with males that care for young or that provide the resources required to produce eggs. For example, brown kiwi females make an enormous initial investment in their offspring—their eggs routinely represent over 15 percent of the mother’s total body weight—but choose to mate with males that take over all of the incubation and other care of the offspring.

85
Male-Male Competition
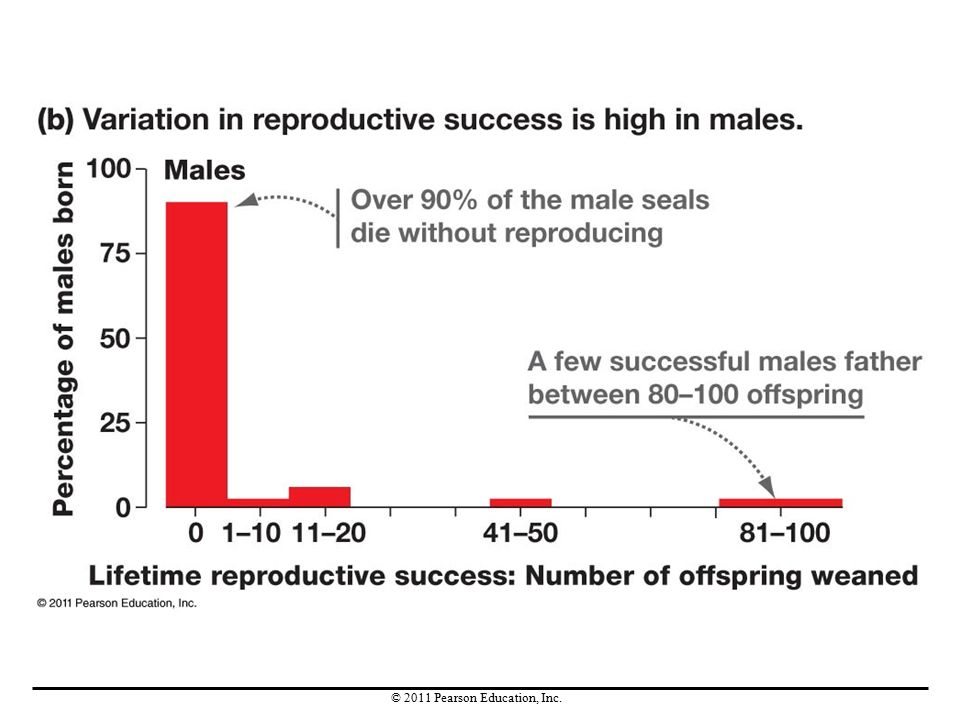
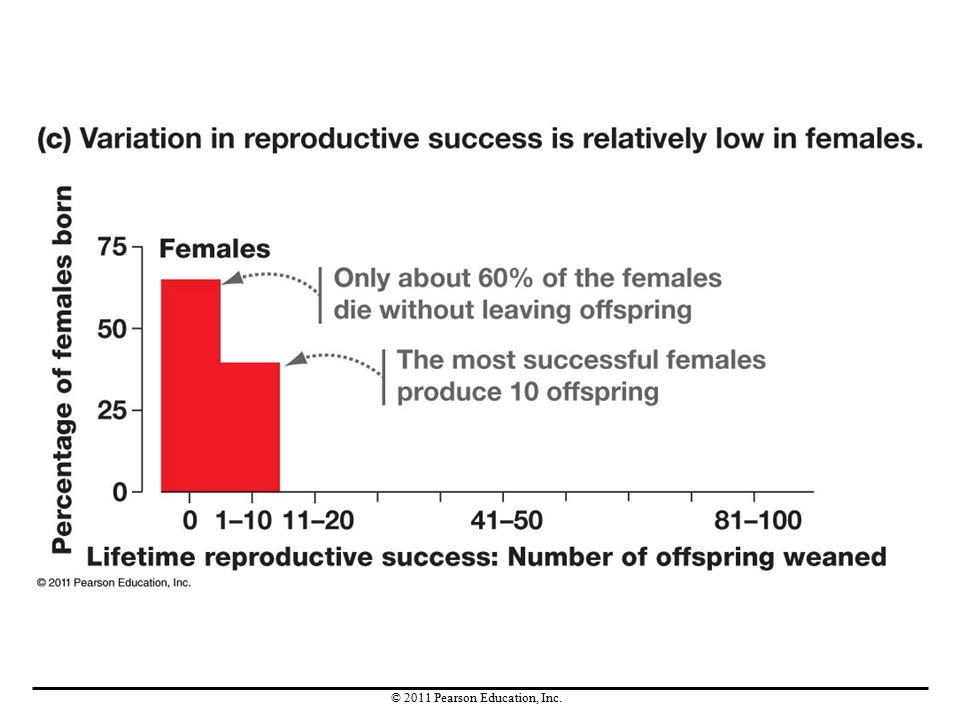
Sexual selection is intense in elephant seals and is driven by male-male competition. Male elephant seals establish territories, areas that they defend and can use exclusively. Evidence for intense sexual selection in males is indicated by the fact that variation in reproductive success is high in males, whereas females have lower variation in reproductive success. Males with larger territories father more offspring, and their alleles rapidly increase in the population.

90
What Are the Consequences of Sexual Selection?
Sexually selected traits often differ sharply between the sexes. Sexual dimorphism refers to any trait that differs between males and females of the same species. Sexual selection violates the assumptions of the Hardy-Weinberg principle by causing certain alleles to increase or decrease in frequency and resulting in evolutionary change.

Similar presentations


























 Genotype frequency.>")








