Download presentation
Presentation is loading. Please wait.

1
The Seed- God’s wonder and tiny mystery YOSHINOBU NAMIHIRA MD,FACG 3000 HALLS FERRY ROAD VICKSBURG, MS 39180 PH 601-638-9800,FAX 601-638-9808 E MAIL: NAMIHIRA @VICKSBURG.COM

3
Germination is the process whereby growth emerges from a period of dormancy. The most common example of germination is the sprouting of a seedling from a seed of an angiosperm or gymnosperm. However, the growth of a sporeling from a spore, for example the growth of hyphae from fungal spores, is also germination. In a more general sense, germination can imply anything expanding into greater being from a small existence or germ.sproutingseedlingseedangiosperm gymnospermsporelingsporehyphaefungalgerm

4
Seed germination Brassica campestris germinating seeds Brassica campestris A germinated seedling (Eranthis hyemalis) emerges from the groundEranthis hyemalis Germination is the growth of an embryonic plant contained within a seed, it results in the formation of the seedling. The seed of a higher plant is a small package produced in a fruit or cone after the union of male and female sex cells. Most seeds go through a period of quiescences where there is no active growth, during this time the seed can be safely transported to a new location and/or survive adverse climate conditions until it is favorable for growth. The seed contains an embryo and in most plants stored food reserves wrapped in a seed coat. Under favorable conditions, the seed begins to germinate, and the embryonic tissues resume growth, developing towards a seedlingembryonic higher plantfruitconequiescencesclimateembryo

5
Requirements for seed germination The germination of seeds is dependent on both internal and external conditions. The most important external factors include: temperature, water, oxygen and sometimes light or darkness. [1] Often different varieties of seeds require distinctive variables for successful germination; some seeds germinate while the soil is cold, while most germinate while the soil is warm. This depends on the individual seed variety and is closely linked to the ecological conditions of the plants' natural habitat.temperaturewateroxygenlight [1]ecological conditionsnatural habitat

6
Water (1) Water - is required for germination. Mature seeds are often extremely dry and need to take in significant amounts of water, relative to the seeds dry weight, before cellular metabolism and growth can resume. Most seeds respond best when there is enough water to moisten the seeds but not soak them. The uptake of water by seeds is called imbibition which leads to the swelling and the breaking of the seed coat. metabolism imbibition

7
Water (2) When seeds are formed, most plants store food, such as starch, proteins, or oils, to provide nourishment to the growing embryo inside the seed. When the seed imbibes water, hydrolytic enzymes are activated that break down these stored food resources in to metabolically useful chemicals, allowing the cells of the embryo to divide and grow, so the seedling can emerge from the seed. [1]starchproteinsoils hydrolytic enzymeschemicals [1]

8
Water (3) Once the seedling starts growing and the food reserves are exhausted, it requires a continuous supply of water, nutrients and light for photosynthesis, which now provides the energy needed for continued growth.
 Once the seedling starts growing and the food reserves are exhausted, it requires a continuous supply of water, nutrients and light for photosynthesis, which now provides the energy needed for continued growth.")
9
oxygen Oxygen - is required by the germinating seed for metabolism: [2] If the soil is waterlogged or the seed is buried within the soil, it might be cut off from the necessary oxygen it needs. Oxygen is used in aerobic respiration, the main source of the seedling's energy until it has leaves, which can photosynthesize its energy requirements. [1] Some seeds have impermeable seed coats that prevent oxygen from entering the seeds, causing seed dormancy. Impermeable seed coats to oxygen or water, are types of physical dormancy which is broken when the seed coat is worn away enough to allow gas exchange or water uptake between the seed and its environment. metabolism [2]soilaerobic respiration [1]
![oxygen Oxygen - is required by the germinating seed for metabolism: [2] If the soil is waterlogged or the seed is buried within the soil, it might be cut off from the necessary oxygen it needs.](http://images.slideplayer.com/6/5660178/slides/slide_9.jpg "Oxygen is used in aerobic respiration, the main source of the seedling s energy until it has leaves, which can photosynthesize its energy requirements. [1] Some seeds have impermeable seed coats that prevent oxygen from entering the seeds, causing seed dormancy. Impermeable seed coats to oxygen or water, are types of physical dormancy which is broken when the seed coat is worn away enough to allow gas exchange or water uptake between the seed and its environment. metabolism [2]soilaerobic respiration [1].")
10
Temperature (1) Temperature - affects cellular metabolic and growth rates. Different seeds germinate over a wide range of temperatures, with many preferring temperatures slightly higher than room-temperature while others germinate just above freezing and others responding to alternation in temperature between warm to cool. Often, seeds have a set of temperature ranges where they will germinate and will not do so above or below this range. In addition, some seeds may require exposure to cold temperature (vernalization) to break dormancy before they can germinate.vernalization
 to break dormancy before they can germinate.vernalization.")
11
Temperature (2) As long as the seed is in its dormant state, it will not germinate even if conditions are favorable. Seeds that are dependent on temperature to end dormancy, have a type of physiological dormancy. For example, seeds requiring the cold of winter are inhibited from germinating until they experience cooler temperatures. For most seeds that require cold for germination 4C is cool enough to end dormancy, but some groups especially with in the family Ranunculaceae and others, need less than -5C. Some seeds will only germinate when temperatures reach hundreds of degrees, as during a forest fire. Without fire, they are unable to crack their seed coats, this is a type of physical dormancy. Ranunculaceaeforest fire

12
Light or darkness (1) Light or darkness - can be a type of environmental trigger for germination in seeds and is a type of physiological dormancy. Most seeds are not affected by light or darkness, but many seeds, including species found in forest settings will not germinate until an opening in the canopy allows them to receive sufficient light for the growing seedling. [1] [1]

13
Light or darkness (2) Stratification mimics natural processes that weaken the seed coat before germination. In nature, some seeds require particular conditions to germinate, such as the heat of a fire (e.g., many Australian native plants), or soaking in a body of water for a long period of time. Others have to be passed through an animal's digestive tract to weaken the seed coat and enable germination. [1]digestive tract [1]
, or soaking in a body of water for a long period of time. Others have to be passed through an animal s digestive tract to weaken the seed coat and enable germination. [1]digestive tract [1].")
14
Dormancy (1) Dormancy Many live seeds have dormancy, meaning they will not germinate even if they have water and it is warm enough for the seedling to grow. Dormancy factors include conditions affecting many different parts of the seed, from the embryo to the seed coat. Dormancy is broken or ended by a number of different conditions and cues both internal and external to the seed. Environmental factors like light, temperature, fire, ingestion by animals and others are conditions that can end seed dormancy.dormancy

15
Dormancy (2) Internally seeds can be dormant because of plant hormones such as absciscic acid, which affects seed dormancy and prevents germination, while the production and application of the hormone gibberellin can break dormancy and induces seed germination. This effect is used in brewing where barley is treated with gibberellin to ensure uniform seed germination to produce barley malt. [1]absciscic acidgibberellinmalt [1]

16
Establishment (1) Seedling establishment In some definitions, the appearance of the radicle marks the end of germination and the beginning of "establishment", a period that ends when the seedling has exhausted the food reserves stored in the seed. Germination and establishment as an independent organism are critical phases in the life of a plant when they are the most vulnerable to injury, disease, and water stress. [1] [1]

17
Establishment (2) The germination index can be used as an indicator of phytotoxicity in soils. The mortality between dispersal of seeds and completion of establishment can be so high, that many species survive only by producing huge numbers of seeds.phytotoxicity

18
Germination rate In agriculture and gardening, germination rate is the number of seeds of a particular plant species, variety or particular seedlot that are likely to germinate. This is usually expressed as a percentage, e.g. an 85% germination rate indicates that about 85 out of 100 seeds will probably germinate under proper conditions. Germination rate is useful in calculating seed requirements for a given area or desired number of plants.agriculturegardeningplantspecies

19
Pollen germination Another germination event during the life cycle of gymnosperms and flowering plants is the germination of a pollen grain after pollination. Like seeds, pollen grains are severely dehydrated before being released to facilitate their dispersal from one plant to another. They consist of a protective coat containing several cells (up to 8 in gymnosperms, 2-3 in flowering plants). One of these cells is a tube cell. Once the pollen grain lands on the stigma of a receptive flower (or a female cone in gymnosperms), it takes up water and germinates. Pollen germination is facilitated by hydration on the stigma, as well as the structure and physiology of the stigma and style. [1] Pollen can also be induced to germinate in vitro (in a petri dish or test tube). [3][4]gymnosperms flowering plantspollinationpollentube cellstigmaflowerconehydration physiology [1] [3][4] During germination, the tube cell elongates into a pollen tube. In the flower, the pollen tube then grows towards the ovule where it discharges the sperm produced in the pollen grain for fertilization. The germinated pollen grain with its two sperm cells is the mature male microgametophyte of these plants. [1pollen tubeovulesperm microgametophyte [1
. One of these cells is a tube cell. Once the pollen grain lands on the stigma of a receptive flower (or a female cone in gymnosperms), it takes up water and germinates. Pollen germination is facilitated by hydration on the stigma, as well as the structure and physiology of the stigma and style. [1] Pollen can also be induced to germinate in vitro (in a petri dish or test tube). [3][4]gymnosperms flowering plantspollinationpollentube cellstigmaflowerconehydration physiology [1] [3][4] During germination, the tube cell elongates into a pollen tube. In the flower, the pollen tube then grows towards the ovule where it discharges the sperm produced in the pollen grain for fertilization. The germinated pollen grain with its two sperm cells is the mature male microgametophyte of these plants. [1pollen tubeovulesperm microgametophyte [1.")
20
root Root From Wikipedia, the free encyclopedia (Redirected from Root (botany))Root (botany) Jump to: navigation, searchnavigationsearch For other uses, see Root (disambiguation).Root (disambiguation) Primary and secondary roots in a cotton plant In vascular plants, the root is the organ of a plant body that typically lies below the surface of the soil. This is not always the case, however, since a root can also be aerial (that is, growing above the ground) or aerating (that is, growing up above the ground or especially above water). Furthermore, a stem normally occurring below ground is not exceptional either (see rhizome). So, it is better to define root as a part of a plant body that bears no leaves, and therefore also lacks nodes. There are also important internal structural differences between stems and roots. The two major functions of roots are 1.) absorption of water and inorganic nutrients and 2.) anchoring the plant body to the ground. Roots also function in cytokinin synthesis, which supplies some of the shoot's needs. They often function in storage of food. The roots of most vascular plant species enter into symbiosis with certain fungi to form mycorrhizas, and a large range of other organisms including bacteria also closely associate with roots.vascular plantsplantaerialrhizomenodescytokininfungimycorrhizasbacteria
 or aerating (that is, growing up above the ground or especially above water). Furthermore, a stem normally occurring below ground is not exceptional either (see rhizome). So, it is better to define root as a part of a plant body that bears no leaves, and therefore also lacks nodes. There are also important internal structural differences between stems and roots. The two major functions of roots are 1.) absorption of water and inorganic nutrients and 2.) anchoring the plant body to the ground. Roots also function in cytokinin synthesis, which supplies some of the shoot s needs. They often function in storage of food. The roots of most vascular plant species enter into symbiosis with certain fungi to form mycorrhizas, and a large range of other organisms including bacteria also closely associate with roots.vascular plantsplantaerialrhizomenodescytokininfungimycorrhizasbacteria.")
21
Root growth Root systems of prairie plantsprairie Cross Section of a mango treemango Root system of a prairie grass Early root growth is one of the functions of the apical meristem located near the tip of the root. The meristem cells more or less continuously divide, producing more meristem, root cap cells (these are sacrificed to protect the meristem), and undifferentiated root cells. The latter will become the primary tissues of the root, first undergoing elongation, a process that pushes the root tip forward in the growing medium. Gradually these cells differentiate and mature into specialized cells of the root tissues. Roots will generally grow in any direction where the correct environment of air, mineral nutrients and water exists to meet the plant's needs. Roots will not grow in dry soil. Over time, given the right conditions, roots can crack foundations, snap water lines, and lift sidewalks. At germination, roots grow downward due to gravitropism, the growth mechanism of plants that also causes the shoot to grow upward. In some plants (such as ivy), the "root" actually clings to walls and structures.airnutrientswatergerminationgravitropismivy Growth from apical meristems is known as primary growth, which encompasses all elongation. Secondary growth encompasses all growth in diameter, a major component of woody plant tissues and many nonwoody plants. For example, storage roots of sweet potato have secondary growth but are not woody. Secondary growth occurs at the lateral meristems, namely the vascular cambium and cork cambium. The former forms secondary xylem and secondary phloem, while the latter forms the periderm.woody plantsweet potatolateral meristemsvascular cambiumcork cambium secondary xylemsecondary phloemperiderm In plants with secondary growth, the vascular cambium, originating between the xylem and the phloem, forms a cylinder of tissue along the stem and root. The cambium layer forms new cells on both the inside and outside of the cambium cylinder, with those on the inside forming secondary xylem cells, and those on the outside forming secondary phloem cells. As secondary xylem accumulates, the "girth" (lateral dimensions) of the stem and root increases. As a result, tissues beyond the secondary phloem (including the epidermis and cortex, in many cases) tend to be pushed outward and are eventually "sloughed off" (shed).cylinderstem At this point, the cork cambium begins to form the periderm, consisting of protective cork cells containing suberin. In roots, the cork cambium originates in the pericycle, a component of the vascular cylinder.cork Stilt roots in the Amazon Rainforest support a tree in very soft, wet soil conditionsAmazon Rainforest The vascular cambium produces new layers of secondary xylem annually. The xylem vessels are dead at maturity but are responsible for most water transport through the vascular tissue in stems and roots.
, and undifferentiated root cells. The latter will become the primary tissues of the root, first undergoing elongation, a process that pushes the root tip forward in the growing medium. Gradually these cells differentiate and mature into specialized cells of the root tissues. Roots will generally grow in any direction where the correct environment of air, mineral nutrients and water exists to meet the plant s needs. Roots will not grow in dry soil. Over time, given the right conditions, roots can crack foundations, snap water lines, and lift sidewalks. At germination, roots grow downward due to gravitropism, the growth mechanism of plants that also causes the shoot to grow upward. In some plants (such as ivy), the root actually clings to walls and structures.airnutrientswatergerminationgravitropismivy Growth from apical meristems is known as primary growth, which encompasses all elongation. Secondary growth encompasses all growth in diameter, a major component of woody plant tissues and many nonwoody plants. For example, storage roots of sweet potato have secondary growth but are not woody. Secondary growth occurs at the lateral meristems, namely the vascular cambium and cork cambium. The former forms secondary xylem and secondary phloem, while the latter forms the periderm.woody plantsweet potatolateral meristemsvascular cambiumcork cambium secondary xylemsecondary phloemperiderm In plants with secondary growth, the vascular cambium, originating between the xylem and the phloem, forms a cylinder of tissue along the stem and root. The cambium layer forms new cells on both the inside and outside of the cambium cylinder, with those on the inside forming secondary xylem cells, and those on the outside forming secondary phloem cells. As secondary xylem accumulates, the girth (lateral dimensions) of the stem and root increases. As a result, tissues beyond the secondary phloem (including the epidermis and cortex, in many cases) tend to be pushed outward and are eventually sloughed off (shed).cylinderstem At this point, the cork cambium begins to form the periderm, consisting of protective cork cells containing suberin. In roots, the cork cambium originates in the pericycle, a component of the vascular cylinder.cork Stilt roots in the Amazon Rainforest support a tree in very soft, wet soil conditionsAmazon Rainforest The vascular cambium produces new layers of secondary xylem annually. The xylem vessels are dead at maturity but are responsible for most water transport through the vascular tissue in stems and roots..")
22
Types of roots A true root system consists of a primary root and secondary roots (or lateral roots). lateral roots The primary root originates in the radicle of the seedling. It is the first part of the root to be originated. During its growth it rebranches to form the lateral roots. It usually grows downwards. Generally, two categories are recognized:radicle the taproot system: the primary root is prominent and has a single, dominant axis; there are fibrous secondary roots running outward. Usually allows for deeper roots capable of reaching low water tables. Most common in dicots. The main function of the taproot is to store food.taprootdicotstaproot the diffuse root system: the primary root is not dominant; the whole root system is fibrous and branches in all directions. Most common in monocots. The main function of the fibrous root is to anchor the plant. monocots

23
Specialized roots Aerating roots of a mangrovemangrove Buttress roots of Ceiba pentandraCeiba pentandra The roots, or parts of roots, of many plant species have become specialized to serve adaptive purposes besides the two primary functions described in the introduction. Adventitious roots arise out-of-sequence from the more usual root formation of branches of a primary root, and instead originate from the stem, branches, leaves, or old woody roots. They commonly occur in monocots and pteridophytes, but also in many dicots, such as clover (Trifolium), ivy (Hedera), strawberry (Fragaria) and willow (Salix). Most aerial roots and stilt roots are adventitious. In some conifers adventitious roots can form the largest part of the root system.monocotsdicotscloverivystrawberrywillow Aerating roots (or pneumatophores): roots rising above the ground, especially above water such as in some mangrove genera (Avicennia, Sonneratia). In some plants like Avicennia the erect roots have a large number of breathing pores for exchange of gases.mangroveAvicennia Sonneratia Aerial roots: roots entirely above the ground, such as in ivy (Hedera) or in epiphytic orchids. They function as prop roots, as in maize or anchor roots or as the trunk in strangler fig.epiphyticorchidsmaizestrangler fig Contractile roots: they pull bulbs or corms of monocots, such as hyacinth and lily, and some taproots, such as dandelion, deeper in the soil through expanding radially and contracting longitudinally. They have a wrinkled surface.monocotshyacinthlilydandelion Coarse roots: Roots that have undergone secondary thickening and have a woody structure. These roots have some ability to absorb water and nutrients, but their main function is transport and to provide a structure to connect the smaller diameter, fine roots to the rest of the plant. Fine roots: Primary roots usually <2 mm diameter that have the function of water and nutrient uptake. They are often heavily branched and support mycorrhizas. These roots may be short lived, but are replaced by the plant in an ongoing process of root 'turnover'. Haustorial roots: roots of parasitic plants that can absorb water and nutrients from another plant, such as in mistletoe (Viscum album) and dodder.mistletoedodder Propagative roots: roots that form adventitious buds that develop into aboveground shoots, termed suckers, which form new plants, as in Canada thistle, cherry and many others.suckersCanada thistlecherry Proteoid roots or cluster roots: dense clusters of rootlets of limited growth that develop under low phosphate or low iron conditions in Proteaceae and some plants from the following families Betulaceae, Casuarinaceae, Eleagnaceae, Moraceae, Fabaceae and Myricaceae. Proteoid rootsphosphateironProteaceaeBetulaceaeCasuarinaceaeEleagnaceaeMoraceaeFabaceaeMyricaceae Stilt roots: these are adventitious support roots, common among mangroves. They grow down from lateral branches, branching in the soil.mangroves Storage roots: these roots are modified for storage of food or water, such as carrots and beets. They include some taproots and tuberous roots.carrotsbeets Structural roots: large roots that have undergone considerable secondary thickening and provide mechanical support to woody plants and trees. Surface roots: These proliferate close below the soil surface, exploiting water and easily available nutrients. Where conditions are close to optimum in the surface layers of soil, the growth of surface roots is encouraged and they commonly become the dominant roots. Tuberous roots: A portion of a root swells for food or water storage, e.g. sweet potato. A type of storage root distinct from taproot.sweet potato
, ivy (Hedera), strawberry (Fragaria) and willow (Salix). Most aerial roots and stilt roots are adventitious. In some conifers adventitious roots can form the largest part of the root system.monocotsdicotscloverivystrawberrywillow Aerating roots (or pneumatophores): roots rising above the ground, especially above water such as in some mangrove genera (Avicennia, Sonneratia). In some plants like Avicennia the erect roots have a large number of breathing pores for exchange of gases.mangroveAvicennia Sonneratia Aerial roots: roots entirely above the ground, such as in ivy (Hedera) or in epiphytic orchids. They function as prop roots, as in maize or anchor roots or as the trunk in strangler fig.epiphyticorchidsmaizestrangler fig Contractile roots: they pull bulbs or corms of monocots, such as hyacinth and lily, and some taproots, such as dandelion, deeper in the soil through expanding radially and contracting longitudinally. They have a wrinkled surface.monocotshyacinthlilydandelion Coarse roots: Roots that have undergone secondary thickening and have a woody structure. These roots have some ability to absorb water and nutrients, but their main function is transport and to provide a structure to connect the smaller diameter, fine roots to the rest of the plant. Fine roots: Primary roots usually <2 mm diameter that have the function of water and nutrient uptake. They are often heavily branched and support mycorrhizas. These roots may be short lived, but are replaced by the plant in an ongoing process of root turnover . Haustorial roots: roots of parasitic plants that can absorb water and nutrients from another plant, such as in mistletoe (Viscum album) and dodder.mistletoedodder Propagative roots: roots that form adventitious buds that develop into aboveground shoots, termed suckers, which form new plants, as in Canada thistle, cherry and many others.suckersCanada thistlecherry Proteoid roots or cluster roots: dense clusters of rootlets of limited growth that develop under low phosphate or low iron conditions in Proteaceae and some plants from the following families Betulaceae, Casuarinaceae, Eleagnaceae, Moraceae, Fabaceae and Myricaceae. Proteoid rootsphosphateironProteaceaeBetulaceaeCasuarinaceaeEleagnaceaeMoraceaeFabaceaeMyricaceae Stilt roots: these are adventitious support roots, common among mangroves. They grow down from lateral branches, branching in the soil.mangroves Storage roots: these roots are modified for storage of food or water, such as carrots and beets. They include some taproots and tuberous roots.carrotsbeets Structural roots: large roots that have undergone considerable secondary thickening and provide mechanical support to woody plants and trees. Surface roots: These proliferate close below the soil surface, exploiting water and easily available nutrients. Where conditions are close to optimum in the surface layers of soil, the growth of surface roots is encouraged and they commonly become the dominant roots. Tuberous roots: A portion of a root swells for food or water storage, e.g. sweet potato. A type of storage root distinct from taproot.sweet potato.")
24
[edit] Rooting depthsedit The distribution of vascular plant roots within soil depends on plant form, the spatial and temporal availability of water and nutrients, and the physical properties of the soil. The deepest roots are generally found in deserts and temperate coniferous forests; the shallowest in tundra, boreal forest and temperate grasslands. The deepest observed living root, at least 60 m below the ground surface, was observed during the excavation of an open-pit mine in Arizona, USA. Some roots can grow as deep as the tree is high. The majority of roots on most plants are however found relatively close to the surface where nutrient availability and aeration are more favourable for growth. Rooting depth may be physically restricted by rock or compacted soil close below the surface, or by anaerobic soil conditions.
![[edit] Rooting depthsedit The distribution of vascular plant roots within soil depends on plant form, the spatial and temporal availability of water and nutrients, and the physical properties of the soil.](http://images.slideplayer.com/6/5660178/slides/slide_24.jpg "The deepest roots are generally found in deserts and temperate coniferous forests; the shallowest in tundra, boreal forest and temperate grasslands. The deepest observed living root, at least 60 m below the ground surface, was observed during the excavation of an open-pit mine in Arizona, USA. Some roots can grow as deep as the tree is high. The majority of roots on most plants are however found relatively close to the surface where nutrient availability and aeration are more favourable for growth. Rooting depth may be physically restricted by rock or compacted soil close below the surface, or by anaerobic soil conditions..")
25
Root architecture The pattern of development of a root system is termed 'root architecture', and is important in providing a plant with a secure supply of nutrients and water as well as anchorage and support. The architecture of a root system can be considered in a similar way to above-ground architecture of a plant - i.e. in terms of the size, branching and distribution of the component parts. In roots, the architecture of fine roots and coarse roots can both be described by variation in topology and distribution of biomass within and between roots. Having a balanced architecture allows fine roots to exploit soil efficiently around a plant, but the 'plastic' nature of root growth allows the plant to then concentrate its resources where nutrients and water are more easily available. A balanced coarse root architecture, with roots distributed relatively evenly around the stem base, is necessary to provide support to larger plants and trees.

26
Economic importance Roots can also protect the environment by holding the soil to prevent soil erosion Tree roots at Cliffs of the Neuse State Park, NC The term root crops refers to any edible underground plant structure, but many root crops are actually stems, such as potato tubers. Edible roots include cassava, sweet potato, beet, carrot, rutabaga, turnip, parsnip, radish, yam and horseradish. Spices obtained from roots include sassafras, angelica, sarsaparilla and licorice.root cropspotatocassavasweet potatobeetcarrot rutabagaturnipparsnipradishyamhorseradish sassafrasangelicasarsaparillalicorice Sugar beet is an important source of sugar. Yam roots are a source of estrogen compounds used in birth control pills. The fish poison and insecticide rotenone is obtained from roots of Lonchocarpus spp. Important medicines from roots are ginseng, aconite, ipecac, gentian and reserpine. Several legumes that have nitrogen-fixing root nodules are used as green manure crops, which provide nitrogen fertilizer for other crops when plowed under. Specialized bald cypress roots, termed knees, are sold as souvenirs, lamp bases and carved into folk art. Native Americans used the flexible roots of white spruce for basketry. Sugar beetYamrotenoneginsengaconiteipecacgentianreserpinebald cypresswhite spruce Tree roots can heave and destroy concrete sidewalks and crush or clog buried pipes. The aerial roots of strangler fig have damaged ancient Mayan temples in Central America and the temple of Angkor Wat in Cambodia. Treestrangler figMayantemplesCentral America Angkor WatCambodia Vegetative propagation of plants via cuttings depends on adventitious root formation. Hundreds of millions of plants are propagated via cuttings annually including chrysanthemum, poinsettia, carnation, ornamental shrubs and many houseplants. Vegetative propagationchrysanthemumpoinsettia carnationshrubshouseplants Roots can also protect the environment by holding the soil to prevent soil erosion.

40
Root water uptake is an important process of water circle and a component of water balance in the field, and it should be understood better and effectively. A quantitative means of describing root water uptake should be established for efficient water use

41
The objectives of this research are to develop a two-dimensional (2D) model of root water uptake for single apple trees and to validate the model with sap flow and soil water content measurements in an orchard.
 model of root water uptake for single apple trees and to validate the model with sap flow and soil water content measurements in an orchard.")
42
Tube-time domain reflectometry (TDR) was used to measure soil volumetric water content, and sap flow sensors based on heat-pulse technology were used to monitor locally the rates of sap flow in the trunk of the apple tree. Also leaf area index (LAI) was measured using the Hemiview system, root density distribution was determined and soil hydraulic characteristics parameters were fitted from measurements. A 2D model of root water uptake was established, which includes root density distribution function, potential transpiration and soil water stress-modified factor.
 was measured using the Hemiview system, root density distribution was determined and soil hydraulic characteristics parameters were fitted from measurements. A 2D model of root water uptake was established, which includes root density distribution function, potential transpiration and soil water stress-modified factor..")
43
The measured data were compared against the outputs of transpiration rate and soil water contents from the numerical simulation of the soil water dynamics that uses Richards' equation for 2D water flow and the established root uptake model. The results showed an excellent agreement between the measured data and the simulated outputs, which indicate that the developed root water uptake model is effective and feasible. Agricultural water management ISSN 0378- 3774 CODEN AWMADF Source / Source 2006, vol. 83, n o 1-2, pp. 119-129 [11 page(s) (article)] (29 ref.)
 (article)] (29 ref.).")
44
A cell wall is a tough, flexible and sometimes fairly rigid layer surrounding a cell, located external to the cell membrane, which provides the cell with structural support, protection, and acts as a filtering mechanism. A major function of the cell wall is to act as a pressure vessel, preventing over-expansion when water enters the cell. They are found in plants, bacteria, fungi, algae, and some archaea. Animals and protozoa do not have cell walls.cellcell membraneplants bacteriafungialgaearchaea Animalsprotozoa

45
The materials in a cell wall vary between species, and in plants and fungi also differ between cell types and developmental stages. In plants, the strongest component of the complex cell wall is a carbohydrate, the glucose polymer called cellulose. In bacteria, peptidoglycan forms the cell wall. Archaean cell walls have various compositions, and may be formed of glycoprotein S-layers, pseudopeptidoglycan, or polysaccharides. Fungi possess cell walls of the glucosamine polymer chitin, and algae typically possess walls constructed of glycoproteins and polysaccharides. However the diatoms have a cell wall composed of silicic acid. Often, other accessory molecules are found anchored to the cell wall.carbohydrateglucose polymercellulosepeptidoglycanglycoproteinS-layers pseudopeptidoglycanpolysaccharidesglucosaminechitindiatomssilicic acid

47
Rigidity The rigidity of cell walls is often over-estimated. In most cells, the cell wall is flexible, meaning that it will bend rather than holding a fixed shape, but has considerable tensile strength. The apparent rigidity of primary plant tissues is a function of hydraulic turgor pressure of the cells and not due to rigid cell walls. This flexibility is seen when plants wilt, so that the stems and leaves begin to droop, or in seaweeds that bend in water currents. The rigidity of healthy plants results from a combination of the wall construction and turgor pressureturgor pressureseaweeds water currents

48
As John Howland states it: “ Think of the cell wall as a wicker basket in which a balloon has been inflated so that it exerts pressure from the inside. Such a basket is very rigid and resistant to mechanical damage. Thus does the prokaryote cell (and eukaryotic cell that possesses a cell wall) gain strength from a flexible plasma membrane pressing against a rigid cell wall. [1] [1]
 gain strength from a flexible plasma membrane pressing against a rigid cell wall. [1] [1].")
49
The rigidity of the cell wall thus results in part from inflation of the cell contained. This inflation is a result of the passive uptake of water.passive uptake of water In plants, a secondary cell wall is a thicker additional layer of cellulose which increases wall rigidity. Additional layers may be formed containing lignin in xylem cell walls, or containing suberin in cork cell walls. These compounds are rigid and waterproof, making the secondary wall stiff. Both wood and bark cells of trees have secondary walls. Other parts of plants such as the leaf stalk may acquire similar reinforcement to resist the strain of physical forces.lignin xylemsuberincorkrigidwaterproofwoodbarktrees leaf stalk

50
Certain single-cell protists and algae also produce a rigid wall. Diatoms build a frustule from silica extracted from the surrounding water; radiolarians also produce a test from minerals. Many green algae, such as the Dasycladales encase their cells in a secreted skeleton of calcium carbonate. In each case, the wall is rigid and essentially inorganic.protistsalgaeDiatomssilicaradiolariansgreen algae Dasycladalescalcium carbonate

52
Permeability The primary cell wall of most plant cells is semi-permeable and permit the passage of small molecules and small proteins, with size exclusion estimated to be 30-60 kDa. Key nutrients, especially water and carbon dioxide, are distributed throughout the plant from cell wall to cell wall in apoplastic flow.plant cells semi-permeablewatercarbon dioxideapoplastic

53
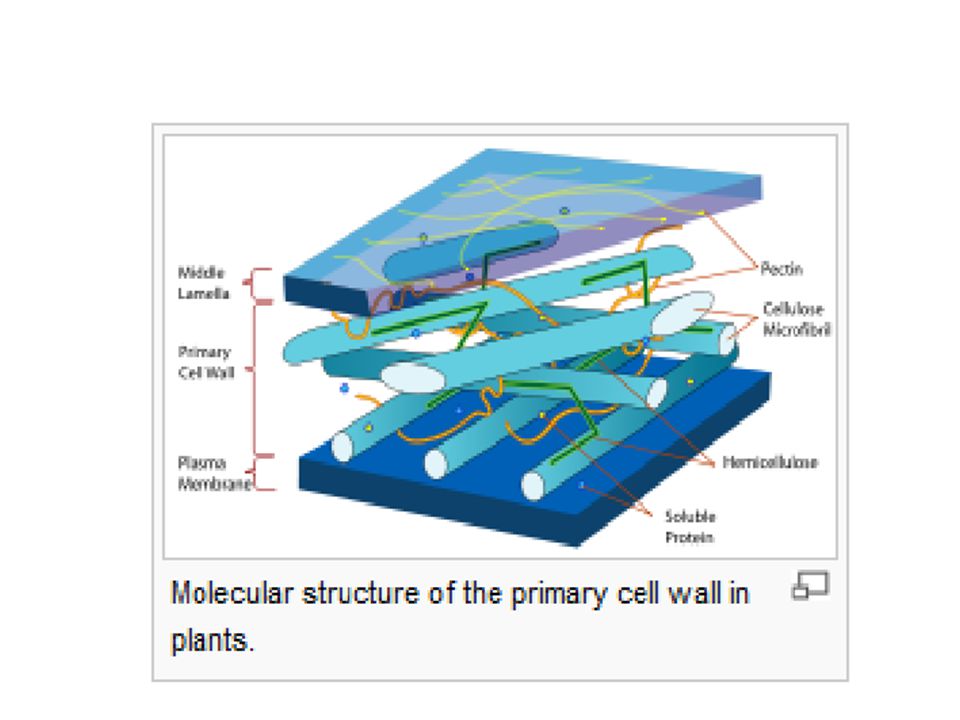
Plant cell walls Composition Molecular structure of the primary cell wall in plants. The major carbohydrates making up the primary (growing) plant cell wall are cellulose, hemicellulose and pectin. The cellulose microfibrils are linked via hemicellulosic tethers to form the cellulose-hemicellulose network, which is embedded in the pectin matrix. The most common hemicellulose in the primary cell wall is xyloglucan. In grass cell walls, xyloglucan and pectin are reduced in abundance and partially replaced by glucuronarabinoxylan, a hemicellulose. Primary cell walls characteristically extend (grow) by a mechanism called acid growth, which involves turgor- driven movement of the strong cellulose microfibrils within the weaker hemicellulose/pectin matrix, catalyzed by expansin proteins. The outer part of the primary cell wall of the plant epidermis is usually impregnated with cutin and wax, forming a permeability barrier known as the plant cuticle.carbohydratescellulosehemicellulosepectinmicrofibrilsxyloglucan glucuronarabinoxylanacid growthturgorexpansincutinwaxplant cuticle
 plant cell wall are cellulose, hemicellulose and pectin. The cellulose microfibrils are linked via hemicellulosic tethers to form the cellulose-hemicellulose network, which is embedded in the pectin matrix. The most common hemicellulose in the primary cell wall is xyloglucan. In grass cell walls, xyloglucan and pectin are reduced in abundance and partially replaced by glucuronarabinoxylan, a hemicellulose. Primary cell walls characteristically extend (grow) by a mechanism called acid growth, which involves turgor- driven movement of the strong cellulose microfibrils within the weaker hemicellulose/pectin matrix, catalyzed by expansin proteins. The outer part of the primary cell wall of the plant epidermis is usually impregnated with cutin and wax, forming a permeability barrier known as the plant cuticle.carbohydratescellulosehemicellulosepectinmicrofibrilsxyloglucan glucuronarabinoxylanacid growthturgorexpansincutinwaxplant cuticle.")
54
Secondary cell walls contain a wide range of additional compounds that modify their mechanical properties and permeability. The major polymers that make up wood (largely secondary cell walls) include cellulose (35 to 50%), xylan, a type of hemicellulose, (20 to 35%) and a complex phenolic polymer called lignin (10 to 25%). Lignin penetrates the spaces in the cell wall between cellulose, hemicellulose and pectin components, driving out water and strengthening the wall. The walls of cork cells in the bark of trees are impregnated with suberin, and suberin also forms the permeability barrier in primary roots known as the Casparian strip. Secondary walls - especially in grasses - may also contain microscopic silica crystals, which may strengthen the wall and protect it from herbivores.polymerswood xylanlignincorksuberinCasparian stripsilica
 include cellulose (35 to 50%), xylan, a type of hemicellulose, (20 to 35%) and a complex phenolic polymer called lignin (10 to 25%). Lignin penetrates the spaces in the cell wall between cellulose, hemicellulose and pectin components, driving out water and strengthening the wall. The walls of cork cells in the bark of trees are impregnated with suberin, and suberin also forms the permeability barrier in primary roots known as the Casparian strip. Secondary walls - especially in grasses - may also contain microscopic silica crystals, which may strengthen the wall and protect it from herbivores.polymerswood xylanlignincorksuberinCasparian stripsilica.")
55
Plant cells walls also contain numerous enzymes, such as hydrolases, esterases, peroxidases, and transglycosylases, that cut, trim and cross link wall polymers. Small amounts (1-5%) of structural proteins are found in most plant cell walls; they are classified as hydroxyproline-rich glycoproteins (HRGP), arabinogalactan proteins (AGP), glycine-rich proteins (GRPs), and proline-rich proteins (PRPs). Each class of glycoprotein is defined by a characteristic, highly repetitive protein sequence. Most are glycosylated, contain hydroxyproline (Hyp) and become cross-linked in the cell wall. These proteins are often concentrated in specialized cells and in cell corners. Cell walls of the epidermis and endodermis may also contain suberin or cutin, two polyester-like polymers that protect the cell from herbivores. [2] The relative composition of carbohydrates, secondary compounds and protein varies between plants and between the cell type and age.proteins glycosylatedhydroxyprolineepidermisendodermissuberincutin [2]
 of structural proteins are found in most plant cell walls; they are classified as hydroxyproline-rich glycoproteins (HRGP), arabinogalactan proteins (AGP), glycine-rich proteins (GRPs), and proline-rich proteins (PRPs). Each class of glycoprotein is defined by a characteristic, highly repetitive protein sequence. Most are glycosylated, contain hydroxyproline (Hyp) and become cross-linked in the cell wall. These proteins are often concentrated in specialized cells and in cell corners. Cell walls of the epidermis and endodermis may also contain suberin or cutin, two polyester-like polymers that protect the cell from herbivores. [2] The relative composition of carbohydrates, secondary compounds and protein varies between plants and between the cell type and age.proteins glycosylatedhydroxyprolineepidermisendodermissuberincutin [2].")
56
Up to three strata or layers may be found in plant cell walls: [3] [3] The middle lamella, a layer rich in pectins. This outermost layer forming the interface between adjacent plant cells and glues them together.middle lamellapectins The primary cell wall, generally a thin, flexible and extensible layer formed while the cell is growing. The secondary cell wall, a thick layer formed inside the primary cell wall after the cell is fully grown. It is not found in all cell types. In some cells, such as found xylem, the secondary wall contains lignin, which strengthens and waterpoofs the wall.secondary cell wall xylemlignin
![Up to three strata or layers may be found in plant cell walls: [3] [3] The middle lamella, a layer rich in pectins.](http://images.slideplayer.com/6/5660178/slides/slide_56.jpg "This outermost layer forming the interface between adjacent plant cells and glues them together.middle lamellapectins The primary cell wall, generally a thin, flexible and extensible layer formed while the cell is growing. The secondary cell wall, a thick layer formed inside the primary cell wall after the cell is fully grown. It is not found in all cell types. In some cells, such as found xylem, the secondary wall contains lignin, which strengthens and waterpoofs the wall.secondary cell wall xylemlignin.")
57
Cell walls in some plant tissues also function as storage depots for carbohydrates that can be broken down and resorbed to supply the metabolic and growth needs of the plant. For example, endosperm cell walls in the seeds of cereal grasses, nasturtium, and other species, are rich in glucans and other polysaccharides that are readily digested by enzymes during seed germination to form simple sugars that nourish the growing embryo. Cellulose microfibrils are not readily digested by plants, however.

61
Making Life Easier. Donate Now » [Expand]Expand Support Wikipedia: a non-profit project Donate Now » [Expand]Expand Support Wikipedia: a non-profit project — Donate NowDonate Now Photosynthesis From Wikipedia, the free encyclopedia Jump to: navigation, searchnavigationsearch This article or section is in need of attention from an expert on the subject. WikiProject Plants or the Plants Portal may be able to help recruit one. If a more appropriate WikiProject or portal exists, please adjust this template accordingly. (November 2008) WikiProject PlantsPlants PortalWikiProjectportal The leaf is the primary site of photosynthesis in plants.leaf Photosynthesis is a metabolic pathway that converts light energy into chemical energy. Its initial substrates are carbon dioxide and water; the energy source is sunlight (electromagnetic radiation); and the end-products are oxygen and (energy-containing) carbohydrates, such as sucrose, glucose or starch. This process is one of the most important biochemical pathways, [1] since nearly all life on Earth either directly or indirectly depends on it as a source of energy. It is a complex process occurring in plants, algae, as well as bacteria such as cyanobacteria. Photosynthetic organisms are also referred to as photoautotrophs. [1]metabolic pathwaylightenergychemical energysubstrates carbon dioxidewaterelectromagnetic radiation oxygencarbohydratessucroseglucosestarch [1]Earthplantsalgaebacteriacyanobacteriaphotoautotrophs [1] The word comes from the Greek φώτο- (photo-), "light," and σύνθεσις (synthesis), "placing with."Greeksynthesis Contents [hide]
 WikiProject PlantsPlants PortalWikiProjectportal The leaf is the primary site of photosynthesis in plants.leaf Photosynthesis is a metabolic pathway that converts light energy into chemical energy. Its initial substrates are carbon dioxide and water; the energy source is sunlight (electromagnetic radiation); and the end-products are oxygen and (energy-containing) carbohydrates, such as sucrose, glucose or starch. This process is one of the most important biochemical pathways, [1] since nearly all life on Earth either directly or indirectly depends on it as a source of energy. It is a complex process occurring in plants, algae, as well as bacteria such as cyanobacteria. Photosynthetic organisms are also referred to as photoautotrophs. [1]metabolic pathwaylightenergychemical energysubstrates carbon dioxidewaterelectromagnetic radiation oxygencarbohydratessucroseglucosestarch [1]Earthplantsalgaebacteriacyanobacteriaphotoautotrophs [1] The word comes from the Greek φώτο- (photo-), light, and σύνθεσις (synthesis), placing with. Greeksynthesis Contents [hide].")
62
Overview Photosynthesis splits water to liberate O 2 and fixes CO 2 into sugar Photosynthesis uses light energy and carbon dioxide to make triose phosphates (G3P). G3P is generally considered the first end-product of photosynthesis. [citation needed] It can be used as a source of metabolic energy, or combined and rearranged to form monosaccharide or disaccharide sugars, such as glucose or sucrose, respectively, which can be transported to other cells, stored as insoluble polysaccharides such as starch, or converted to structural carbohydrates, such as cellulose or glucans.triose phosphatescitation neededmonosaccharidedisaccharideglucosesucrose polysaccharidesstarchcellulose glucans A commonly used slightly simplified equation for photosynthesis is:equation 6 CO 2(g) + 12 H 2 O (l) + photons → C 6 H 12 O 6(aq) + 6 O 2(g) + 6 H 2 O (l) carbon dioxide + water + light energy → glucose + oxygen + water The equation is often presented in introductory chemistry texts in an even more simplified form as: [2]photonsC 6 H 12 O 6 [2] 6 CO 2(g) + 6 H 2 O (l) + photons → C 6 H 12 O 6(aq) + 6 O 2(g) Photosynthesis occurs in two stages. In the first stage, light-dependent reactions or photosynthetic reactions (also called the Light Reactions) capture the energy of light and use it to make high-energy molecules. During the second stage, the light-independent reactions (also called the Calvin-Benson Cycle, and formerly known as the Dark Reactions) use the high-energy molecules to capture and chemically reduce carbon dioxide (CO 2 ) (also called carbon fixation) to make the precursors of carbohydrates.photonsCalvin-Benson Cyclechemically reducecarbon fixationprecursorscarbohydrates
 + 12 H 2 O (l) + photons → C 6 H 12 O 6(aq) + 6 O 2(g) + 6 H 2 O (l) carbon dioxide + water + light energy → glucose + oxygen + water The equation is often presented in introductory chemistry texts in an even more simplified form as: [2]photonsC 6 H 12 O 6 [2] 6 CO 2(g) + 6 H 2 O (l) + photons → C 6 H 12 O 6(aq) + 6 O 2(g) Photosynthesis occurs in two stages. In the first stage, light-dependent reactions or photosynthetic reactions (also called the Light Reactions) capture the energy of light and use it to make high-energy molecules. During the second stage, the light-independent reactions (also called the Calvin-Benson Cycle, and formerly known as the Dark Reactions) use the high-energy molecules to capture and chemically reduce carbon dioxide (CO 2 ) (also called carbon fixation) to make the precursors of carbohydrates.photonsCalvin-Benson Cyclechemically reducecarbon fixationprecursorscarbohydrates.")
63
In the Light-independent or dark reactions the enzyme RuBisCO captures CO 2 from the atmosphere and in a process that requires the newly formed NADPH, called the Calvin-Benson Cycle, releases three-carbon sugars, which are later combined to form sucrose and starch.Light-independentenzymeRuBisCOCO 2atmosphere Photosynthesis may simply be defined as the conversion of light energy into chemical energy by living organisms. It is affected by its surroundings, and the rate of photosynthesis is affected by the concentration of carbon dioxide in the air, the light intensity, and the temperature.chemical energyorganismstemperature Photosynthesis uses only 1% of the entire electromagnetic spectrum, and 2% of the visible spectrum. [citation needed] It has been estimated that the productivity of photosythesis is 115 petagrams (Pg, equals 10 15 grams or 10 9 metric tons). [citation needed]citation needed
. [citation needed]citation needed.")
65
In plants Most plants are photoautotrophs, which means that they are able to synthesize food directly from inorganic compounds using light energy - for example from the sun, instead of eating other organisms or relying on nutrients derived from them. This is distinct from chemoautotrophs that do not depend on light energy, but use energy from inorganic compounds.photoautotrophs synthesizeinorganic compoundschemoautotrophs 6 CO 2 + 12 H 2 O → C 6 H 12 O 6 + 6 O 2 + 6 H 2 O The energy for photosynthesis ultimately comes from absorbed photons and involves a reducing agent, which is water in the case of plants, releasing oxygen as product. The light energy is converted to chemical energy (known as light-dependent reactions), in the form of ATP and NADPH, which are used for synthetic reactions in photoautotrophs. The overall equation for the light- dependent reactions under the conditions of non-cyclic electron flow in green plants is: [3]photonsreducing agentwateroxygenproductlight-dependent reactionsATPNADPH [3] 2 H 2 O + 2 NADP + + 2 ADP + 2 P i + light → 2 NADPH + 2 H + + 2 ATP + O 2
, in the form of ATP and NADPH, which are used for synthetic reactions in photoautotrophs. The overall equation for the light- dependent reactions under the conditions of non-cyclic electron flow in green plants is: [3]photonsreducing agentwateroxygenproductlight-dependent reactionsATPNADPH [3] 2 H 2 O + 2 NADP ADP + 2 P i + light → 2 NADPH + 2 H ATP + O 2.")
66
Most notably, plants use the chemical energy to fix carbon dioxide into carbohydrates and other organic compounds through light-independent reactions. The overall equation for carbon fixation (sometimes referred to as carbon reduction) in green plants is: [3]carbon dioxide carbohydrateslight-independent reactions [3] 3 CO 2 + 9 ATP + 6 NADPH + 6 H + → C 3 H 6 O 3 -phosphate + 9 ADP + 8 P i + 6 NADP + + 3 H 2 O To be more specific, carbon fixation produces an intermediate product, which is then converted to the final carbohydrate products. The carbon skeletons produced by photosynthesis are then variously used to form other organic compounds, such as the building material cellulose, as precursors for lipid and amino acid biosynthesis, or as a fuel in cellular respiration. The latter occurs not only in plants but also in animals when the energy from plants gets passed through a food chain. Organisms dependent on photosynthetic and chemosynthetic organisms are called heterotrophs. In general outline, cellular respiration is the opposite of photosynthesis: Glucose and other compounds are oxidized to produce carbon dioxide, water, and chemical energy. However, the two processes take place through a different sequence of chemical reactions and in different cellular compartments.celluloselipid amino acidcellular respirationanimalsfood chainchemosyntheticheterotrophs
 in green plants is: [3]carbon dioxide carbohydrateslight-independent reactions [3] 3 CO ATP + 6 NADPH + 6 H + → C 3 H 6 O 3 -phosphate + 9 ADP + 8 P i + 6 NADP H 2 O To be more specific, carbon fixation produces an intermediate product, which is then converted to the final carbohydrate products. The carbon skeletons produced by photosynthesis are then variously used to form other organic compounds, such as the building material cellulose, as precursors for lipid and amino acid biosynthesis, or as a fuel in cellular respiration. The latter occurs not only in plants but also in animals when the energy from plants gets passed through a food chain. Organisms dependent on photosynthetic and chemosynthetic organisms are called heterotrophs. In general outline, cellular respiration is the opposite of photosynthesis: Glucose and other compounds are oxidized to produce carbon dioxide, water, and chemical energy. However, the two processes take place through a different sequence of chemical reactions and in different cellular compartments.celluloselipid amino acidcellular respirationanimalsfood chainchemosyntheticheterotrophs.")
67
Plants absorb light primarily using the pigment chlorophyll, which is the reason that most plants have a green color. The function of chlorophyll is often supported by other accessory pigments such as carotenes and xanthophylls. Both chlorophyll and accessory pigments are contained in organelles (compartments within the cell) called chloroplasts. Although all cells in the green parts of a plant have chloroplasts, most of the energy is captured in the leaves. The cells in the interior tissues of a leaf, called the mesophyll, can contain between 450,000 and 800,000 chloroplasts for every square millimeter of leaf. The surface of the leaf is uniformly coated with a water-resistant waxy cuticle that protects the leaf from excessive evaporation of water and decreases the absorption of ultraviolet or blue light to reduce heating. The transparent epidermis layer allows light to pass through to the palisade mesophyll cells where most of the photosynthesis takes place.pigmentchlorophyllaccessory pigments carotenesxanthophyllsorganelles cellchloroplasts leaves mesophyllwaxycuticleevaporationultravioletbluelightheatingepidermis palisade
 called chloroplasts. Although all cells in the green parts of a plant have chloroplasts, most of the energy is captured in the leaves. The cells in the interior tissues of a leaf, called the mesophyll, can contain between 450,000 and 800,000 chloroplasts for every square millimeter of leaf. The surface of the leaf is uniformly coated with a water-resistant waxy cuticle that protects the leaf from excessive evaporation of water and decreases the absorption of ultraviolet or blue light to reduce heating. The transparent epidermis layer allows light to pass through to the palisade mesophyll cells where most of the photosynthesis takes place.pigmentchlorophyllaccessory pigments carotenesxanthophyllsorganelles cellchloroplasts leaves mesophyllwaxycuticleevaporationultravioletbluelightheatingepidermis palisade.")
68
Plant convert light into chemical energy with a maximum photosynthetic efficiency of approximately 6%. [4][5][6] By comparison solar panels convert light into electric energy at a photosynthetic efficiency of approximately 10- 20%. Actual plant's photosynthetic efficiency varies with the frequency of the light being converted, light intensity, temperature and proportion of CO 2 in atmosphere.chemical energyphotosynthetic efficiency [4][5][6]solar panelselectric energyCO 2

69
Origin of chloroplasts In plants photosynthesis occurs in organelles called chloroplasts. Chloroplasts have many similarities with photosynthetic bacteria including a circular chromosome, prokaryotic-type ribosomes, and similar proteins in the photosynthetic reaction center. [14][15]organelles chloroplasts photosynthetic bacteriachromosomeribosomes [14][15] The endosymbiotic theory suggests that photosynthetic bacteria were acquired (by endocytosis) by early eukaryotic cells to form the first plant cells. Therefore, chloroplasts may be photosynthetic bacteria that adapted to life inside plant cells. Like mitochondria, chloroplasts still possess their own DNA, separate from the nuclear DNA of their plant host cells and the genes in this chloroplast DNA resemble those in cyanobacteria. [16]endosymbiotic theoryendocytosiseukaryoticplantmitochondriacyanobacteria [16]
 by early eukaryotic cells to form the first plant cells. Therefore, chloroplasts may be photosynthetic bacteria that adapted to life inside plant cells. Like mitochondria, chloroplasts still possess their own DNA, separate from the nuclear DNA of their plant host cells and the genes in this chloroplast DNA resemble those in cyanobacteria. [16]endosymbiotic theoryendocytosiseukaryoticplantmitochondriacyanobacteria [16].")
70
Marine molluscs Elysia viridis and Elysia chlorotica likewise maintain a symbiotic relationship with chloroplasts that they capture from the algae that they ingest. This allows the molluscs to survive solely by photosynthesis for several months at a time. [17]molluscsElysia viridisElysia chloroticasymbiotic [17]

73
Molecular production Light-dependent reactions of photosynthesis at the thylakoid membrane Temporal Order The overall process of photosynthesis takes place in four stages. The first, energy transfer in antenna chlorophyll takes place in the femtosecond [1 femtosecond (fs) = 10, −15 s] to picosecond [1 picosecond (ps) = 10 −12 s] time scale. The next phase, the transfer of electrons in photochemical reactions, takes place in the picosecond to nanosecond time scale [1 nanosecond (ns) = 10 −9 s]. The third phase, the electron transport chain and ATP synthesis, takes place on the microsecond [1 microsecond (μs) = 10 −6 s] to millisecond [1 millisecond (ms) = 10 −3 s) time scale. The final phase is carbon fixation and export of stable products and takes place in the millisecond to second time scale. The first three stages occur in the thylakoid membranes.
 = 10, −15 s] to picosecond [1 picosecond (ps) = 10 −12 s] time scale. The next phase, the transfer of electrons in photochemical reactions, takes place in the picosecond to nanosecond time scale [1 nanosecond (ns) = 10 −9 s]. The third phase, the electron transport chain and ATP synthesis, takes place on the microsecond [1 microsecond (μs) = 10 −6 s] to millisecond [1 millisecond (ms) = 10 −3 s) time scale. The final phase is carbon fixation and export of stable products and takes place in the millisecond to second time scale. The first three stages occur in the thylakoid membranes..")
75
Light to chemical energy Main article: Light-dependent reactionLight-dependent reaction The light energy is converted to chemical energy using the light-dependent reactions. This chemical energy production is about 5-6% efficient, with the majority of the light that strikes a plant reflected and not absorbed. [4] However, of the energy that is absorbed, approximately 30-50% is captured as chemical energy. [19] The products of the light-dependent reactions are ATP from photophosphorylation and NADPH from photoreduction. Both are then utilized as an energy source for the light-independent reactions.light-dependent reactions [4] [19]light-dependent reactionsATPphotophosphorylation NADPHlight-independent reactions

76
Not all wavelengths of light can support photosynthesis. The photosynthetic action spectrum depends on the type of accessory pigments present. For example, in green plants, the action spectrum resembles the absorption spectrum for chlorophylls and carotenoids with peaks for violet-blue and red light. In red algae, the action spectrum overlaps with the absorption spectrum of phycobilins for blue-green light, which allows these algae to grow in deeper waters that filter out the longer wavelengths used by green plants. The non-absorbed part of the light spectrum is what gives photosynthetic organisms their color (e.g., green plants, red algae, purple bacteria) and is the least effective for photosynthesis in the respective organisms.wavelengthsaccessory pigmentsaction spectrumabsorption spectrumchlorophyllscarotenoidsphycobilins
 and is the least effective for photosynthesis in the respective organisms.wavelengthsaccessory pigmentsaction spectrumabsorption spectrumchlorophyllscarotenoidsphycobilins.")
77
Z scheme A Photosystem: A light-harvesting cluster of photosynthetic pigments present in the thylakoid membrane of chloroplasts. The "Z scheme" In plants, light-dependent reactions occur in the thylakoid membranes of the chloroplasts and use light energy to synthesize ATP and NADPH. The light-dependent reaction has two forms; cyclic and non-cyclic reaction. In the non-cyclic reaction, the photons are captured in the light-harvesting antenna complexes of photosystem II by chlorophyll and other accessory pigments (see diagram at right). When a chlorophyll molecule at the core of the photosystem II reaction center obtains sufficient excitation energy from the adjacent antenna pigments, an electron is transferred to the primary electron-acceptor molecule, Pheophytin, through a process called Photoinduced charge separation. These electrons are shuttled through an electron transport chain, the so called Z-scheme shown in the diagram, that initially functions to generate a chemiosmotic potential across the membranelight-dependent reactionsthylakoid membraneschloroplastsphotons antenna complexesphotosystem IIchlorophyllaccessory pigments Photoinduced charge separation electron transport chainchemiosmotic potential
. When a chlorophyll molecule at the core of the photosystem II reaction center obtains sufficient excitation energy from the adjacent antenna pigments, an electron is transferred to the primary electron-acceptor molecule, Pheophytin, through a process called Photoinduced charge separation. These electrons are shuttled through an electron transport chain, the so called Z-scheme shown in the diagram, that initially functions to generate a chemiosmotic potential across the membranelight-dependent reactionsthylakoid membraneschloroplastsphotons antenna complexesphotosystem IIchlorophyllaccessory pigments Photoinduced charge separation electron transport chainchemiosmotic potential.")
78
An ATP synthase enzyme uses the chemiosmotic potential to make ATP during photophosphorylation, whereas NADPH is a product of the terminal redox reaction in the Z-scheme. The electron enters the Photosystem I molecule. The electron is excited due to the light absorbed by the photosystem. A second electron carrier accepts the electron, which again is passed down lowering energies of electron acceptors. The energy created by the electron acceptors is used to move hydrogen ions across the thylakoid membrane into the lumen. The electron is used to reduce the co-enzyme NADP, which has functions in the light-independent reaction. The cyclic reaction is similar to that of the non-cyclic, but differs in the form that it generates only ATP, and no reduced NADP (NADPH) is created. The cyclic reaction takes place only at photosystem I. Once the electron is displaced from the photosystem, the electron is passed down the electron acceptor molecules and returns back to photosystem I, from where it was emitted, hence the name cyclic reaction.ATP synthaseNADPHredoxphotosystemelectron acceptors
 is created. The cyclic reaction takes place only at photosystem I. Once the electron is displaced from the photosystem, the electron is passed down the electron acceptor molecules and returns back to photosystem I, from where it was emitted, hence the name cyclic reaction.ATP synthaseNADPHredoxphotosystemelectron acceptors.")
80
Water photolysis Main articles: Photodissociation and Oxygen evolutionPhotodissociationOxygen evolution The NADPH is the main reducing agent in chloroplasts, providing a source of energetic electrons to other reactions. Its production leaves chlorophyll with a deficit of electrons (oxidized), which must be obtained from some other reducing agent. The excited electrons lost from chlorophyll in photosystem I are replaced from the electron transport chain by plastocyanin. However, since photosystem II includes the first steps of the Z-scheme, an external source of electrons is required to reduce its oxidized chlorophyll a molecules. The source of electrons in green-plant and cyanobacterial photosynthesis is waterreducing agentphotosystem Iplastocyanin photosystem II
, which must be obtained from some other reducing agent. The excited electrons lost from chlorophyll in photosystem I are replaced from the electron transport chain by plastocyanin. However, since photosystem II includes the first steps of the Z-scheme, an external source of electrons is required to reduce its oxidized chlorophyll a molecules. The source of electrons in green-plant and cyanobacterial photosynthesis is waterreducing agentphotosystem Iplastocyanin photosystem II.")
81
Two water molecules are oxidized by four successive charge-separation reactions by photosystem II to yield a molecule of diatomic oxygen and four hydrogen ions; the electron yielded in each step is transferred to a redox-active tyrosine residue that then reduces the photoxidized paired- chlorophyll a species called P680 that serves as the primary (light-driven) electron donor in the photosystem II reaction center. The oxidation of water is catalyzed in photosystem II by a redox-active structure that contains four manganese ions and a calcium ion; this oxygen-evolving complex binds two water molecules and stores the four oxidizing equivalents that are required to drive the water-oxidizing reaction. Photosystem II is the only known biological enzyme that carries out this oxidation of water. The hydrogen ions contribute to the transmembrane chemiosmotic potential that leads to ATP synthesis. Oxygen is a waste product of light-independent reactions, but the majority of organisms on Earth use oxygen for cellular respiration, including photosynthetic organisms. [20][21]oxygenhydrogentyrosinecatalyzedmanganeseoxygen-evolving complexenzymecellular respiration [20][21]

83
Quantum mechanical effects Through photosynthesis, sunlight energy is transferred to molecular reaction centers for conversion into chemical energy with nearly 100-percent efficiency. The transfer of the solar energy takes place almost instantaneously, so little energy is wasted as heat. Of the total incident solar radiation only 43% can be used (only light in the range 400-700 nm), 80% of light makes it through the canopy, photosynthesis stores 28.6% of the energy, and plant respiration uses some energy which leaves 67% of the stored energy behind. This brings the actual efficiency of photosynthesis to about 6.6%. [6] [6] A study led by researchers with the U.S. Department of Energy’s Lawrence Berkeley National Laboratory (Berkeley Lab) and the University of California at Berkeley suggests that long-lived wavelike electronic quantum coherence plays an important part in this instantaneous transfer of energy by allowing the photosynthetic system to simultaneously try each potential energy pathway and choose the most efficient option. Results of the study are presented in the April 12, 2007 issue of the journal Nature. [22]U.S. Department of EnergyLawrence Berkeley National LaboratoryUniversity of California at Berkeleyquantum coherenceNature [22]
, 80% of light makes it through the canopy, photosynthesis stores 28.6% of the energy, and plant respiration uses some energy which leaves 67% of the stored energy behind. This brings the actual efficiency of photosynthesis to about 6.6%. [6] [6] A study led by researchers with the U.S. Department of Energy’s Lawrence Berkeley National Laboratory (Berkeley Lab) and the University of California at Berkeley suggests that long-lived wavelike electronic quantum coherence plays an important part in this instantaneous transfer of energy by allowing the photosynthetic system to simultaneously try each potential energy pathway and choose the most efficient option. Results of the study are presented in the April 12, 2007 issue of the journal Nature. [22]U.S. Department of EnergyLawrence Berkeley National LaboratoryUniversity of California at Berkeleyquantum coherenceNature [22].")
84
Oxygen and photosynthesis With respect to oxygen and photosynthesis, there are two important concepts. Plant and cyanobacterial (blue-green algae) cells also use oxygen for cellular respiration, although they have a net output of oxygen since much more is produced during photosynthesis.cyanobacterial Oxygen is a product of the light-driven water-oxidation reaction catalyzed by photosystem II; it is not generated by the fixation of carbon dioxide. Consequently, the source of oxygen during photosynthesis is water, not carbon dioxide.photosystem II
 cells also use oxygen for cellular respiration, although they have a net output of oxygen since much more is produced during photosynthesis.cyanobacterial Oxygen is a product of the light-driven water-oxidation reaction catalyzed by photosystem II; it is not generated by the fixation of carbon dioxide. Consequently, the source of oxygen during photosynthesis is water, not carbon dioxide.photosystem II.")
85
Bacterial variation The concept that oxygen production is not directly associated with the fixation of carbon dioxide was first proposed by Cornelis Van Niel in the 1930s, who studied photosynthetic bacteria. Aside from the cyanobacteria, bacteria only have one photosystem and use reducing agents other than water. They get electrons from a variety of different inorganic chemicals including sulfide or hydrogen, so for most of these bacteria oxygen is not produced.Cornelis Van Nielcyanobacteriaphotosystemsulfidehydrogen Others, such as the halophiles (an Archaea), produced so-called purple membranes where the bacteriorhodopsin could harvest light and produce energy. The purple membranes was one of the first to be used to demonstrate the chemiosmotic theory: light hit the membranes and the pH of the solution that contained the purple membranes dropped as protons were pumping out of the membrane.halophilesArchaeabacteriorhodopsinchemiosmotic
, produced so-called purple membranes where the bacteriorhodopsin could harvest light and produce energy. The purple membranes was one of the first to be used to demonstrate the chemiosmotic theory: light hit the membranes and the pH of the solution that contained the purple membranes dropped as protons were pumping out of the membrane.halophilesArchaeabacteriorhodopsinchemiosmotic.")
86
Carbon fixation Main articles: Carbon fixation and Light-independent reactionCarbon fixationLight-independent reaction The fixation or reduction of carbon dioxide is a light-independent process in which carbon dioxide combines with a five-carbon sugar, ribulose 1,5- bisphosphate (RuBP), to yield two molecules of a three-carbon compound, glycerate 3-phosphate (GP), also known as 3-phosphoglycerate (PGA). GP, in the presence of ATP and NADPH from the light-dependent stages, is reduced to glyceraldehyde 3-phosphate (G3P). This product is also referred to as 3-phosphoglyceraldehyde (PGAL) or even as triose phosphate. Triose is a 3-carbon sugar (see carbohydrates). Most (5 out of 6 molecules) of the G3P produced is used to regenerate RuBP so the process can continue (see Calvin-Benson cycle). The 1 out of 6 molecules of the triose phosphates not "recycled" often condense to form hexose phosphates, which ultimately yield sucrose, starch and cellulose. The sugars produced during carbon metabolism yield carbon skeletons that can be used for other metabolic reactions like the production of amino acids and lipids.carbon dioxideribulose 1,5- bisphosphate glycerate 3-phosphateATPNADPHglyceraldehyde 3-phosphatePGALTriosecarbohydratesCalvin-Benson cyclehexosesucrosestarchcellulosemetabolismamino acidslipids
. This product is also referred to as 3-phosphoglyceraldehyde (PGAL) or even as triose phosphate. Triose is a 3-carbon sugar (see carbohydrates). Most (5 out of 6 molecules) of the G3P produced is used to regenerate RuBP so the process can continue (see Calvin-Benson cycle). The 1 out of 6 molecules of the triose phosphates not recycled often condense to form hexose phosphates, which ultimately yield sucrose, starch and cellulose. The sugars produced during carbon metabolism yield carbon skeletons that can be used for other metabolic reactions like the production of amino acids and lipids.carbon dioxideribulose 1,5- bisphosphate glycerate 3-phosphateATPNADPHglyceraldehyde 3-phosphatePGALTriosecarbohydratesCalvin-Benson cyclehexosesucrosestarchcellulosemetabolismamino acidslipids.")
88
Seed germination (1) Seed Germination A seed certainly looks dead. It does not seem to move, to grow, nor do anything. In fact, even with biochemical tests for the metabolic processes we associate with life (respiration, etc.) the rate of these processes is so slow that it would be difficult to determine whether there really was anything alive in a seed.
 the rate of these processes is so slow that it would be difficult to determine whether there really was anything alive in a seed..")
89
Seed germination (2) Indeed if a seed is not allowed to germinate (sprout) within some certain length of time, the embryo inside will die. Each species of seed has a certain length of viability. Some maple species have seeds that need to sprout within two weeks of being dispersed, or they die. Some seeds of Lotus plants are known to be up to 2000 years old and still can be germinated.

90
Seed, dormancy, germination Seeds Lacking True Dormancy Common vegetable garden seeds generally lack any kind of dormancy. The seeds are ready to sprout. All they need is some moisture to get their biochemistry activated, and temperature warm enough to allow the chemistry of life to proceed. Seeds taken from the wild, however, are frequently endowed with deeper forms of dormancy.

91
Assuming the seed is still viable, the embryo inside the seed coat needs something to get its metabolism activated to start the embryo growing. The process of getting a seed to germinate can be simple or complicated, and this our present subject.

92
Thick seed coat Thick Seed Coat Many kinds of seeds have very thick seed coats. These obviously keep water out of the seed, so the embryo cannot get the water needed to activate its metabolism and start growing. The lotus seeds are an example of this. An outstanding example from the northern temperate zone is the Kentucky coffee tree (Gymnocladus dioica). The seed coat is perhaps two millimeters thick! You can throw them as hard as you can against a concrete sidewalk and they just bounce! How could such a seed actually sprout?
. The seed coat is perhaps two millimeters thick. You can throw them as hard as you can against a concrete sidewalk and they just bounce. How could such a seed actually sprout .")
93
The Kentucky coffee tree holds its seed pods in the the top of the tree all winter. The inside of the pod is fleshy (lots of water). The pods are very dark in color. If you put the fickle winter and sunshine and darkness into this picture, I think you can come up with the answer. Here is a hint: you might want to recall what happens if you fill the ice cube trays in your freezer too full with water, or you might recall what happens to a container filled with soda that is then frozen.
. The pods are very dark in color. If you put the fickle winter and sunshine and darkness into this picture, I think you can come up with the answer. Here is a hint: you might want to recall what happens if you fill the ice cube trays in your freezer too full with water, or you might recall what happens to a container filled with soda that is then frozen..")
94
scarification Other species might use some pounding along a river or drop seeds into seacoast surf to abrade the thick seed coat. Some of the sea beans do this. Other seeds might need an vertebrate or other animal to attack the seed coat (but give up trying to eat the seed) and thereby weaken the coat. The process of nicking the thick seed coat to initiate germination is called scarification.
 and thereby weaken the coat. The process of nicking the thick seed coat to initiate germination is called scarification..")
95
A final, and very common, example of a way to scarify a seed coat is observed in strawberry and raspberry. The thick seed coat is designed to be swallowed by the frugivore. The animal digests the fruit pulp, but the seed coat passes through the digestive system still protecting the viable embryo inside, but weakened enough to allow sprouting! The seed is deposited with a little organic fertilizer in the environment and can now sprout!

96
Thin seed coat (1) Thin Seed Coat A thin seed coat is so thin that it is no barrier to water. Some other kind of dormancy mechanism is needed. Knowing that light can penetrate thin layers of plant tissue (leaves for example) should give you the idea that light might be a signal. That plants can absorb light and respond biochemically is a fact you know from your study of photosynthesis. All we need is a pigment molecule that can absorb light and cause a change in the behavior of the embryo.
 should give you the idea that light might be a signal. That plants can absorb light and respond biochemically is a fact you know from your study of photosynthesis. All we need is a pigment molecule that can absorb light and cause a change in the behavior of the embryo..")
97
Thin seed coat (2) The pigment is phytochrome. Like chlorophyll, it is made of a chromophore with tetrapyrole structure and is associated with proteins. This pigment is different from chlorophyll, however, in one critical way. It exists in two inter-convertible form

98
Phytochrome,Pfr One form of phytochrome, named Pfr, is the form of the phytochome found in plant cells that are exposed to red (660 nm) or common white light. This form of phytochrome is biologically very active and plays a role in all systems when a plant needs to know if the lights are "on" or "off." In lettuce (Lactuca sativa) seeds, Pfr causes the seeds to begin to germinate as we will soon see. Thus lettuce seeds germinate only when placed in white or red light. Buried in deep soil, they will not germinate. Given that lettuce has a small seed, I think you can figure out why evolution arrived at this solution.
 seeds, Pfr causes the seeds to begin to germinate as we will soon see. Thus lettuce seeds germinate only when placed in white or red light. Buried in deep soil, they will not germinate. Given that lettuce has a small seed, I think you can figure out why evolution arrived at this solution..")
99
Phytochrome, Pr The other form of phytochrome, named Pr, is formed when phytochrome is exposed to far-red (730 nm) light. This form is biologically inactive or inhibits responses. Thus if lettuce seeds are placed in far-red light they do not germinate. Large seeds have lots of storage material. If their seed coat is very thin, their evolution may have arrived at a completely different response. Think about pea seeds. They are large and have very thin seed coats. How would they respond to light?

100
Insufficient Development If a seed's embryo is not completely developed, some additional maturation may be needed before the seed can sprout. This happens in seeds with little-to-no storage material invested in the seed. Examples include orchid seeds. They are the size of dust and have almost nothing but a very immature embryo on-board. Such a seed needs an association with fungi in the soil or other environments to feed the developing embryo until the embryo is mature enough to actually penetrate the seed coat. These seeds are also likely to have a very brief viability. The fungal association must be established rapidly or the embryo dies.

101
Inhibitors Present Many plant species invest chemicals in the developing seeds, and these chemicals inhibit the development of the embryos. They keep the embryos dormant. Obviously the seed must have some way to eliminate these chemicals before they can sprout.

102
Abscisic Acid Many temperate zone species that use inhibitors use abscisic acid. This chemical induces dormancy in the embryo. The chemical is produced in abundance in the late summer and early fall. The seeds in the fruits become dormant so, even if they are dispersed in autumn, they cannot sprout. During the winter enzymes in the seeds degrade the abscisic acid. By spring the abscisic acid is gone and the seed can sprout. We can collect seeds of these species and get them to sprout early. The seeds are put in moist soil and refrigerated for about four weeks (a process often called stratification). This is sufficient time to degrade the abscisic acid. Then the planted seeds are placed in a warm greenhouse. The seeds assume winter is over, spring has come, and they begin to sprout. This process is called vernalization. If you think of "vernal" as meaning "spring" then you understand how we got this name!
. This is sufficient time to degrade the abscisic acid. Then the planted seeds are placed in a warm greenhouse. The seeds assume winter is over, spring has come, and they begin to sprout. This process is called vernalization. If you think of vernal as meaning spring then you understand how we got this name!.")
103
Phenolic Compounds Plants that live in deserts have a different problem. There is no cold, moist, winter to allow vernalization of abscisic acid. These plants instead use more potent toxins, phenolic compounds, to keep their seeds dormant until the proper season for germination. Phenolic compounds are freely water-soluble, the plant is living in a desert. Deserts typically have very long dry seasons and a short wet season accompanied by flash floods and so on. How do you think the phenolic compounds are lost? How would the mechanism ensure that seeds do not sprout in the dry season, but only after the seed could be sure it is in the wet season? The word leaching might give you a hint?

104
This is a diagram of the seed of barley (Hordeum jubatum) As any "typical" seed it has three fundamental parts: a seed coat a storage area (in this case the endosperm) and a dormant embryo The seed coat is really a fruit coat. In all grains, the seed coat (former ovule integument) is fused to the ovary wall (the true fruit wall). So in fact, the grain is technically a fruit (caryopsis) even though we often call it a seed. Barley is used primarily in beer making. Brewers use barley as a source of sugar to make the alcohol for beer. Brewers knew that sprouting the barley seeds improves the sugar yields tremendously, but they wanted even more sugar yield. The brewers came to plant physiologists to find out if anything could be done. Study of the seed showed that the seed/fruit coat is water-resistant and thereby reduces the rate of water uptake by the seed, and water uptake is essential for seed germination and the improved sugar yields.
 is fused to the ovary wall (the true fruit wall). So in fact, the grain is technically a fruit (caryopsis) even though we often call it a seed. Barley is used primarily in beer making. Brewers use barley as a source of sugar to make the alcohol for beer. Brewers knew that sprouting the barley seeds improves the sugar yields tremendously, but they wanted even more sugar yield. The brewers came to plant physiologists to find out if anything could be done. Study of the seed showed that the seed/fruit coat is water-resistant and thereby reduces the rate of water uptake by the seed, and water uptake is essential for seed germination and the improved sugar yields..")
105
The sugar content of a dry barley seed is actually quite low, but the endosperm holds a huge reserve of starch. Starch is a polymer of sugars. In a sense it is a long chain of sugar molecules linked together. This is probably the source of the sugar. The barley embryo has three parts: the cotyledon (or seed leaf). Since there is only one, barley is a monocot. the epicotyl (becomes the shoot) the radicle (becomes the root) Seed germination is said to have occurred when growth of the radicle bursts the seed coat and protrudes as a young root. The energy for seed germination probably comes from respiration of the sugar in the endosperm. However, the embryo and the starch are separated from each other. There must be some chemical communications that physiologists could manipulate to enhance sugar yield for the brewers.
. Since there is only one, barley is a monocot. the epicotyl (becomes the shoot) the radicle (becomes the root) Seed germination is said to have occurred when growth of the radicle bursts the seed coat and protrudes as a young root. The energy for seed germination probably comes from respiration of the sugar in the endosperm. However, the embryo and the starch are separated from each other. There must be some chemical communications that physiologists could manipulate to enhance sugar yield for the brewers..")
106
The endosperm of the seed has two parts. The bulk of the volume is a starch storage area. The covering layer is called the aleurone layer. The aleurone is made of cells that store protein in abundance. You probably know about this in the case of rice. Rice grains are treated before we buy them for cooking. The fruit/seed coat is milled off and discarded as it has little food value. The embryo breaks out and is called the "germ." In wheat, the embryos are sold as wheat germ. The milling process leaves the rice endosperm surrounded by aleurone. This is sold as brown rice and has food value in starch and protein. It has an interesting texture after cooking. If the endosperm is polished before it is sold, the aleurone can be removed. Polishing results in white rice which has food value in starch only. Thus white rice, while considered much "finer" than brown rice, is actually a lower-quality food.

107
Now we will proceed with what the physiologists figured out... The first step in barley seed germination is imbibition. In this process, water penetrates the seed coat and begins to soften the hard, dry tissues inside. The water uptake causes the grain to swell up. The seed/fruit coat usually splits open allowing water to enter even faster. The water begins to activate the biochemistry of the dormant embryo.

108
The water coming into the seed and embryo dissolves a chemical made inside the embryo. This chemical is called Gibberellic Acid (GA). It is a plant hormone, not too different from steroids. The dissolved GA is transported with the water through the rest of the seed tissues until it arrives at the aleurone layer.
. It is a plant hormone, not too different from steroids. The dissolved GA is transported with the water through the rest of the seed tissues until it arrives at the aleurone layer..")
109
The Gibberelic Acid which is transported by the water arrives at the aleurone layer. The GA crosses into the cytoplasm of the aleurone cells and turns on certain genes in the nuclear DNA. DNA is, of course, the hereditary molecule and contains the instructions for making every protein needed for the survival of a barley plant. The precise mechanism of how GA turns on the DNA is unknown at present. It is clear however that the mode of action is to turn on just certain genes in the DNA. The genes that are turned on are transcribed. The information archived in the DNA is precious, so the aleurone cells make a disposable RNA copy of the gene that is turned on. This disposable copy of the information, a kind of blueprint, is often called messenger RNA. The process of making this RNA copy is called transcription.

110
The RNA that was made in the transcription process is transported into the cytoplasm of the aleurone cells. In the cytoplasm, the messenger RNA joins up with a ribosome to begin the process of making a protein. This process is often called protein synthesis or translation. In this process, the ribosome examines the information held in the sequence of bases in the RNA. Transfer RNAs charged with particular amino acids are moved into the position specified by the instructions in the messenger RNA, and the amino acids are joined in a proper sequence by the ribosome. The sequence of amino acids determines the properties of the protein being assembled.

111
In this case, the critical protein made with the information held in the RNA is amylase. This protein turns out to be an enzyme of great importance. The process of information held in the genes of DNA being transcribed into RNA and then translated into protein constitutes the central dogma of genetics. You will want to remember later, that it is the signal of Gibberellic acid that initiates this whole chain of events.

112
A question one might ask right away is: from where do the amino acid building blocks for the amylase come? The answer is: from some other biochemistry in the aleurone cells. This biochemistry causes the storage proteins in the aleurone cells to be digested by hydrolytic enzymes. The hydrolysis is accelerated by enzymes known as proteases. These enzymes increase the rate at which the storage protein is cut into individual amino acids. The amino acids released by the hydrolysis are then free to be reassembled by the ribosomes into the structure of amylase.

113
The same thing happens in people. You are not what you eat! You do not slowly become a steer by eating beef. The protein in your hamburger is digested into amino acids. Those amino acids are then reassembled into human proteins. Since the instructions for reassembling the amino acids come from your human DNA, the proteins produced are human, not bovine! In a similar way, barley aleurone storage proteins are digested and amylase is made from the released amino acids.

114
So, we have our amylase produced, so what? This is when the payoff begins! The amylase is secreted (transported out) from the aleurone cells into the endosperm. Amylase is not just any old protein. It happens to be an enzyme. It catalyzes (accelerates) a particular chemical reaction. Specifically, the amylase speeds the hydrolysis of starch into its component sugar units. Hurrah! The physiologists now know how the sugar is made in a seed. They can also tell the brewers how to increase the sugar yields for beer making. What do they tell them? I am sure your powers of recall will help you. What signal started this whole cascade of events? Note that Gibberellic Acid was the first plant hormone purified in large quantities to be successfully used to increase the yield of any economically-important biochemical in plants!
 from the aleurone cells into the endosperm. Amylase is not just any old protein. It happens to be an enzyme. It catalyzes (accelerates) a particular chemical reaction. Specifically, the amylase speeds the hydrolysis of starch into its component sugar units. Hurrah. The physiologists now know how the sugar is made in a seed. They can also tell the brewers how to increase the sugar yields for beer making. What do they tell them. I am sure your powers of recall will help you. What signal started this whole cascade of events. Note that Gibberellic Acid was the first plant hormone purified in large quantities to be successfully used to increase the yield of any economically-important biochemical in plants!.")
115
Okay, so we helped out our brewer friends, but we need to finish up this project with barley seed germination. Remember that we wanted to get the seed germinated (sprouted). This does not occur until after the released sugar is transported from the endosperm to the embryo. The cotyledon is a major sugar transfer area in grains. Active sugar uptake is its specialized function. The sugar is transported into the embryo and used as a fuel and building block for growth of the embryo itself. The resulting embryo growth will include the emergence of the radicle from the seed/fruit coat. Germination is thus complete! Almost the end of the story...
. This does not occur until after the released sugar is transported from the endosperm to the embryo. The cotyledon is a major sugar transfer area in grains. Active sugar uptake is its specialized function. The sugar is transported into the embryo and used as a fuel and building block for growth of the embryo itself. The resulting embryo growth will include the emergence of the radicle from the seed/fruit coat. Germination is thus complete. Almost the end of the story....")
117
Our barley seed story is great, but it does not explain some other forms of seed germination. The next great finding in the study of seed germination is what happens in lettuce seeds (Lactuca sativa). Lettuce seeds look somewhat different from barley seeds, because they are dicots. They also have separate seed coat and fruit walls. But the things you buy as lettuce "seeds" are really lettuce fruits (called cypselas). As different as they are, the seed germination process in lettuce needs just one step more than barley to get its seeds germinating.
. Lettuce seeds look somewhat different from barley seeds, because they are dicots. They also have separate seed coat and fruit walls. But the things you buy as lettuce seeds are really lettuce fruits (called cypselas). As different as they are, the seed germination process in lettuce needs just one step more than barley to get its seeds germinating..")
118
Lettuce cypselas have very thin fruit walls and seed coats, so light can penetrate these coverings. As is typical of small seeds, light is needed to stimulate seed germination. One might wonder how this would fit into what we just found out with barley. To keep things simple, I am not drawing a lettuce seed here, but am showing you what happens in lettuce seeds by attaching the additional step to the diagram of a barley seed we have been studying (remember lettuce is a dicot and lacks an aleurone, etc.):
:.")
119
Notice the new step in the diagram. The DNA must first be photoactivated before GA can have its effect. This is accomplished by the formation of a chemical signal known as Pfr. Pfr is one of two possible chemical forms of a molecule known as phytochrome. Phytochrome is a pigment that absorbs light energy. When it does absorb light energy its internal chemical structure is altered instantly. Thus Pfr can be converted to the alternate Pr form of phytochrome by simply hitting the seeds with a flash of far-red light (730 nm). Similarly, the Pr form of phytochrome can be converted back into Pfr with a flash of red light (660 nm).
. Similarly, the Pr form of phytochrome can be converted back into Pfr with a flash of red light (660 nm)..")
120
Dry lettuce seeds have slightly more than half of their phytochrome in the Pfr form. So if they are moistened and kept in the dark, some of the seeds will actually germinate. If we shine red (or it turns out even white) light on imbibing seeds, then virtually all of the phytochrome is converted to the Pfr form and this photoactivates the genes in the DNA. Almost all of the seeds will germinate in this light! On the other hand, if we shine far-red light on imbibing seeds, then virtually all of the phytochrome in the seeds is converted to the Pr form and none of the seeds will germinate.
 light on imbibing seeds, then virtually all of the phytochrome is converted to the Pfr form and this photoactivates the genes in the DNA. Almost all of the seeds will germinate in this light. On the other hand, if we shine far-red light on imbibing seeds, then virtually all of the phytochrome in the seeds is converted to the Pr form and none of the seeds will germinate..")
121
Thus phytochrome and light tell lettuce seeds when they are exposed to sunlight and can begin to germinate. Small seeds, such as those in lettuce, do not have reserves enough to start germinating without being able to quickly get to light. It is no surprise that small-seeded species have evolved this mechanism for photoactivation of seed germination. It prevents a deeply-buried seed from trying to germinate! As a challenge to your own thinking, you might wonder what happens in the case of a species with large seeds (peas = Pisum sativum) but with thin seed coats. How might the strategy be similar? How might it differ significantly?
 but with thin seed coats. How might the strategy be similar. How might it differ significantly .")
122
SEED GERMINATION A seed contains an embryonic plant in a resting condition, and germination is its resumption of growth. Seeds will begin to germinate when the soil temperature is in the appropriate range and when water and oxygen are available. However, most of your wildflower seeds will only germinate between specific soil temperatures. Optimum soil germination temperatures will vary greatly from one species to another. With soil temperatures extremely low or excessively high, growth of the seed is either slow and erratic or germination is entirely prevented. Not all of your seeds will sprout at the same time due to constant temperature fluctuations typically observed in nature. The most common cause of poor germination associated with wildflowers is the depth at which the seeds are sown.

123
Small seeds should be planted on the soil surface and pressed or rolled in for best results since they contain only enough stored food for a limited period of growth. If the seedling is to survive, it must emerge from the soil and quickly begin to produce its own food. If seeds are too deeply buried beneath the soil surface, the seedling will either exhaust its food reserve prior to reaching the soil surface causing its death, or lack of sufficient oxygen will prohibit germination altogether. Within the descriptive literature of each species, we have listed the following germination data to assist you in your planting.

124
GERMINATION DAYS: The average time period in which a particular variety will germinate given optimum conditions. Please allow a variance of plus or minus 25%. OPTIMUM SOIL TEMPERATURE FOR GERMINATION: The optimum temperature in degrees Fahrenheit for rapid germination. A steady temperature between the published limits, based on laboratory data, is recommended for best results. SOWING DEPTH: An increment in inches has been included for each variety as a quick reference for optimum sowing depth. A good rule to follow, if in doubt about any variety, is to sow the seed at a shallow depth. SURFACE SOW: Due to the extremely small size of the seed, the area should be prepared and the seed sown directly on the surface of the soil, then pressed or rolled in. DO NOT COVER.

Similar presentations