Download presentation
Presentation is loading. Please wait.

2
Summary of sixth lesson Janzen-Connol hypothesis; explanation of why diseases lead to spatial heterogeneity Diseases also lead to heterogeneity or changes through time –Driving succession –The Red Queen Hypothesis: selection pressure will increase number of resistant plant genotypes Co-evolution: pathogen increase virulence in short term, but in long term balance between host and pathogen Complexity of forest diseases: primary vs. secondaruy, modes of dispersal etc

3
Summary of seventh lesson SEX; the great homogenizing force, and also ability to create new alleles INTERSTERILITY/ MATING> SOMATIC COMPATIBILITY NEED TO USE MULTIPLE MARKERS; SC does that, otherwise go to molecular markers PCR/ RAPDS

7
How to get multiple loci? Random genomic markers: –RAPDS –Total genome RFLPS (mostly dominant) –AFLPS Microsatellites SNPs Multiple specific loci –SSCP –RFLP –Sequence information Watch out for linked alleles (basically you are looking at the same thing!)
 –AFLPS Microsatellites SNPs Multiple specific loci –SSCP –RFLP –Sequence information Watch out for linked alleles (basically you are looking at the same thing!).")
8
Sequence information Codominant Molecules have different rates of mutation, different molecules may be more appropriate for different questions 3rd base mutation Intron vs. exon Secondary tertiary structure limits Homoplasy

9
Sequence information Multiple gene genealogies=definitive phylogeny Need to ensure gene histories are comparable” partition of homogeneity test Need to use unlinked loci

10
Thermalcycler DNA template Forward primer Reverse primer

11
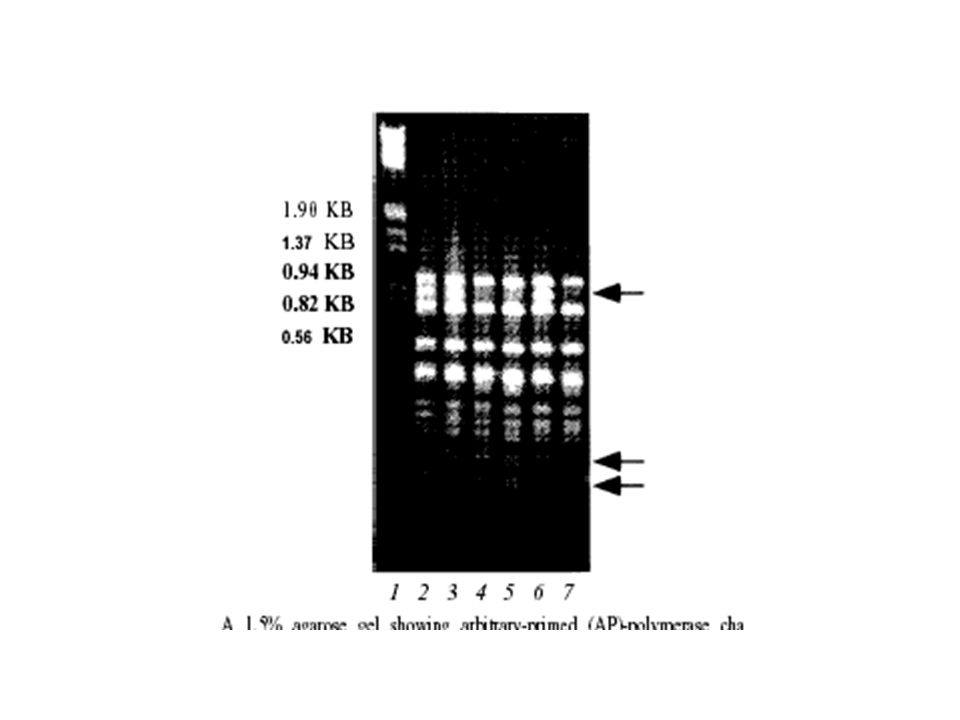
Gel electrophoresis to visualize PCR product Ladder (to size DNA product)
")
12
From DNA to genetic information (alleles are distinct DNA sequences) Presence or absence of a specific PCR amplicon (size based/ specificity of primers) Differerentiate through: –Sequencing –Restriction endonuclease –Single strand conformation polymorphism
 Presence or absence of a specific PCR amplicon (size based/ specificity of primers) Differerentiate through: –Sequencing –Restriction endonuclease –Single strand conformation polymorphism")
13
Presence absence of amplicon AAAGGGTTTCCCNNNNNNNNN CCCGGGTTTAAANNNNNNNNN AAAGGGTTTCCC (primer)
")
14
Presence absence of amplicon AAAGGGTTTCCCNNNNNNNNN CCCGGGTTTAAANNNNNNNNN AAAGGGTTTCCC (primer)
")
15
Result: series of bands that are present or absent (1/0)
")
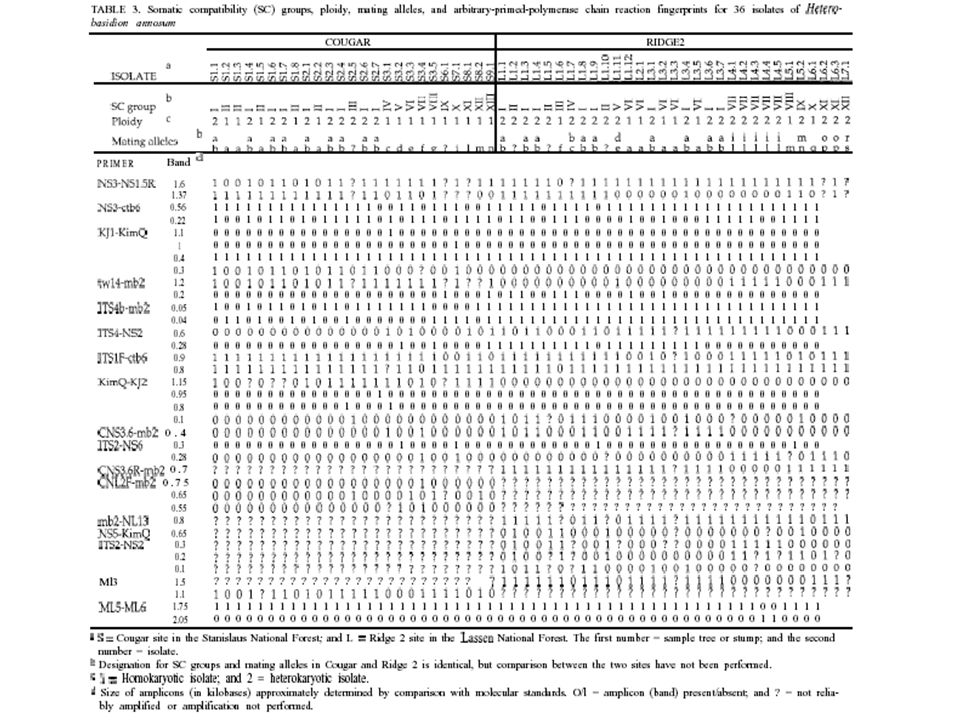
17
Root disease center in true fir caused by H. annosum

18
Ponderosa pineIncense cedar

19
Yosemite Lodge 1975 Root disease centers outlined

20
Yosemite Lodge 1997 Root disease centers outlined

24
Are my haplotypes sensitive enough? To validate power of tool used, one needs to be able to differentiate among closely related individual Generate progeny Make sure each meiospore has different haplotype Calculate P

25
RAPD combination 1 2 1010101010 1010000000 1011101010 1010111010 1010001010 1011001010 1011110101

26
Conclusions Only one RAPD combo is sensitive enough to differentiate 4 half-sibs (in white) Mendelian inheritance? By analysis of all haplotypes it is apparent that two markers are always cosegregating, one of the two should be removed

27
AFLP Amplified Fragment Length Polymorphisms Dominant marker Scans the entire genome like RAPDs More reliable because it uses longer PCR primers less likely to mismatch Priming sites are a construct of the sequence in the organism and a piece of synthesized DNA

28
How are AFLPs generated? AGGTCGCTAAAATTTT (restriction site in red) AGGTCG CTAAATTT Synthetic DNA piece ligated –NNNNNNNNNNNNNNCTAAATTTTT Created a new PCR priming site –NNNNNNNNNNNNNNCTAAATTTTT Every time two PCR priming sitea are within 400- 1600 bp you obtain amplification
 AGGTCG CTAAATTT Synthetic DNA piece ligated –NNNNNNNNNNNNNNCTAAATTTTT Created a new PCR priming site –NNNNNNNNNNNNNNCTAAATTTTT Every time two PCR priming sitea are within bp you obtain amplification.")
29
AFLPs are read like RAPDs (0/1)
")
30
Dealing with dominant anonymous multilocus markers Need to use large numbers (linkage) Repeatability Graph distribution of distances Calculate distance using Jaccard’s similarity index
 Repeatability Graph distribution of distances Calculate distance using Jaccard’s similarity index")
31
Jaccard’s Only 1-1 and 1-0 count, 0-0 do not count 1010011 1001011 1001000

32
Jaccard’s Only 1-1 and 1-0 count, 0-0 do not count A: 1010011 AB= 0.60.4 (1-AB) B: 1001011 BC=0.50.5 C: 1001000 AC=0.20.8
 B: BC= C: AC=0.20.8")
33
Now that we have distances…. Plot their distribution (clonal vs. sexual)
")
34
Now that we have distances…. Plot their distribution (clonal vs. sexual) Analysis: –Similarity (cluster analysis); a variety of algorithms. Most common are NJ and UPGMA
 Analysis: –Similarity (cluster analysis); a variety of algorithms. Most common are NJ and UPGMA.")
35
Now that we have distances…. Plot their distribution (clonal vs. sexual) Analysis: –Similarity (cluster analysis); a variety of algorithms. Most common are NJ and UPGMA –AMOVA; requires a priori grouping
 Analysis: –Similarity (cluster analysis); a variety of algorithms. Most common are NJ and UPGMA –AMOVA; requires a priori grouping.")
36
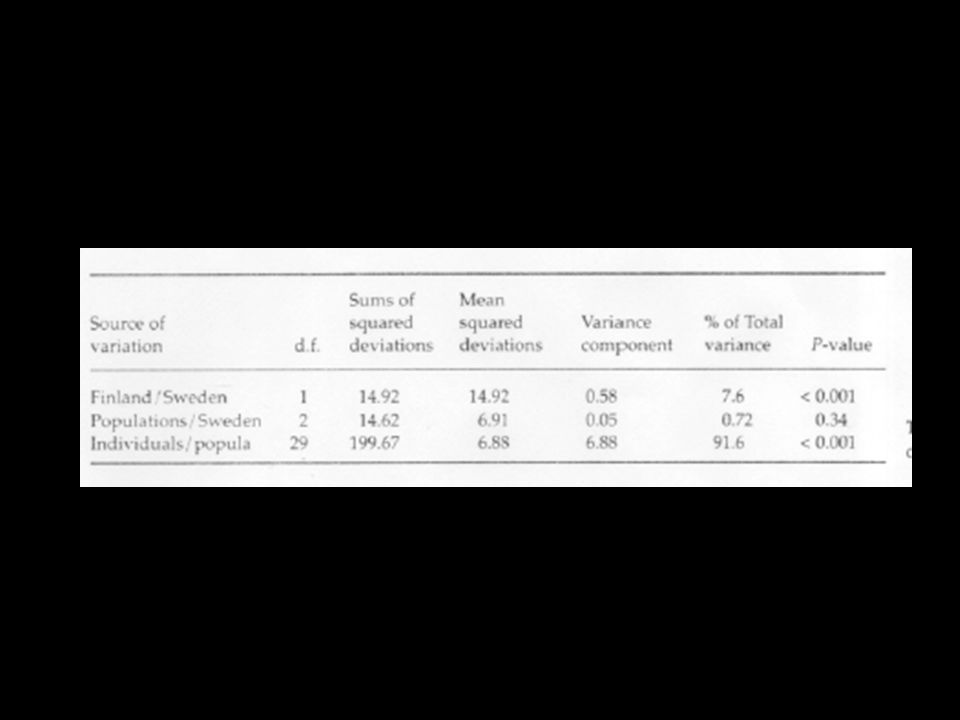
AMOVA groupings Individual Population Region AMOVA: partitions molecular variance amongst a priori defined groupings

37
Now that we have distances…. Plot their distribution (clonal vs. sexual) Analysis: –Similarity (cluster analysis); a variety of algorithms. Most common are NJ and UPGMA –AMOVA; requires a priori grouping –Discriminant, canonical analysis
 Analysis: –Similarity (cluster analysis); a variety of algorithms. Most common are NJ and UPGMA –AMOVA; requires a priori grouping –Discriminant, canonical analysis.")
38
Results: Jaccard similarity coefficients 0.3 0.900.920.94 0.960.98 1.00 0 0.1 0.2 0.4 0.5 0.6 0.7 Coefficient Frequency P. nemorosa P. pseudosyringae: U.S. and E.U. 0.3 Coefficient 0.900.920.940.960.981.00 0 0.1 0.2 0.4 0.5 0.6 0.7 Frequency

39
0.90.910.920.930.940.950.960.970.980.99 Pp U.S. Pp E.U. 0.0 0.1 0.2 0.3 0.4 0.5 0.6 Jaccard coefficient of similarity 0.7 P. pseudosyringae genetic similarity patterns are different in U.S. and E.U.

40
P. nemorosa P. ilicis P. pseudosyringae Results: P. nemorosa

41
Results: P. pseudosyringae P. nemorosa P. ilicis P. pseudosyringae = E.U. isolate

42
Now that we have distances…. Plot their distribution (clonal vs. sexual) Analysis: –Similarity (cluster analysis); a variety of algorithms. Most common are NJ and UPGMA –AMOVA; requires a priori grouping –Discriminant, canonical analysis –Frequency: does allele frequency match expected (hardy weinberg), F or Wright’s statistsis
 Analysis: –Similarity (cluster analysis); a variety of algorithms. Most common are NJ and UPGMA –AMOVA; requires a priori grouping –Discriminant, canonical analysis –Frequency: does allele frequency match expected (hardy weinberg), F or Wright’s statistsis.")
43
The “scale” of disease Dispersal gradients dependent on propagule size, resilience, ability to dessicate, NOTE: not linear Important interaction with environment, habitat, and niche availability. Examples: Heterobasidion in Western Alps, Matsutake mushrooms that offer example of habitat tracking Scale of dispersal (implicitely correlated to metapopulation structure)---
---.")
44
Have we sampled enough? Resampling approaches Saturation curves –A total of 30 polymorphic alleles –Our sample is either 10 or 20 –Calculate whether each new sample is characterized by new alleles

45
Saturation curves 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 No Of New alleles

46
If we have codominant markers how many do I need IDENTITY tests = probability calculation based on allele frequency… Multiplication of frequencies of alleles 10 alleles at locus 1 P1=0.1 5 alleles at locus 2 P2=0,2 Total P= P1*P2=0.02

47
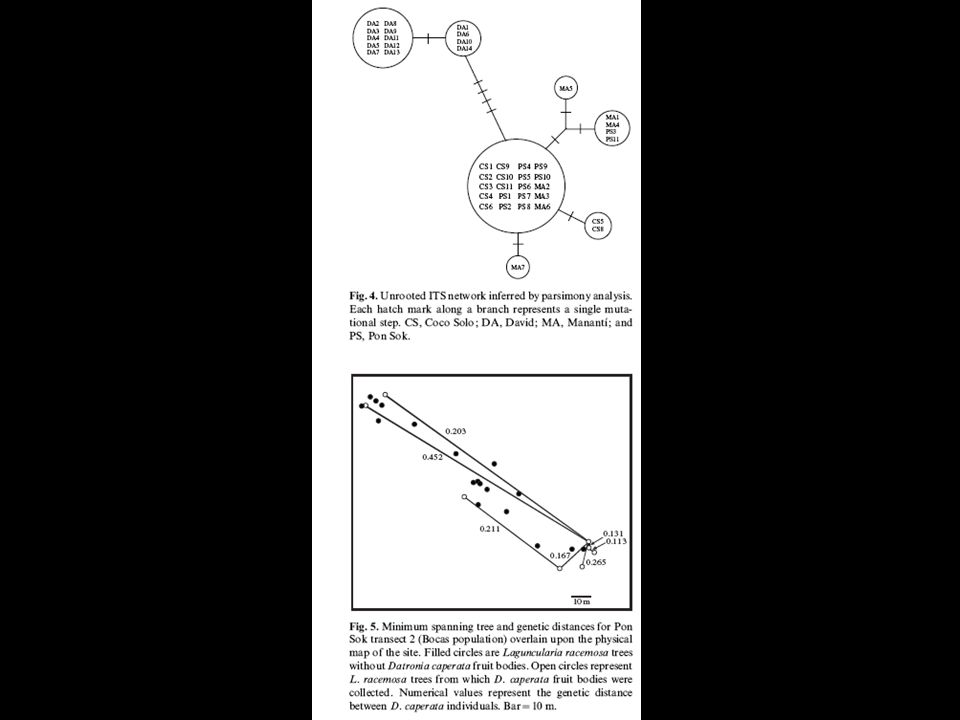
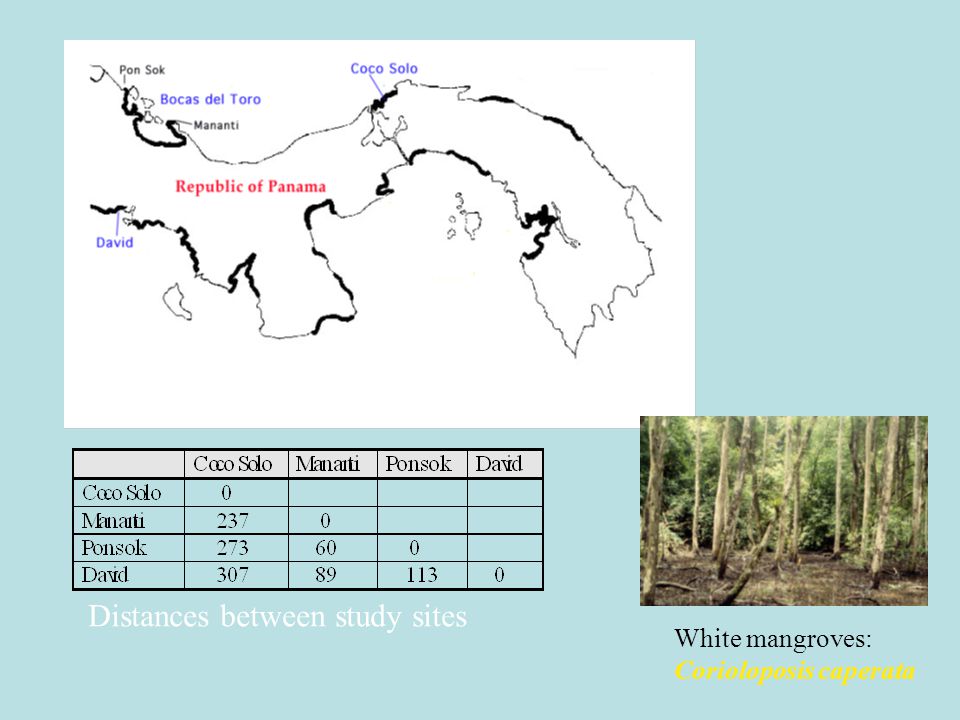
White mangroves: Corioloposis caperata

51
White mangroves: Corioloposis caperata Distances between study sites

52
Coriolopsis caperata on Laguncularia racemosa Forest fragmentation can lead to loss of gene flow among previously contiguous populations. The negative repercussions of such genetic isolation should most severely affect highly specialized organisms such as some plant- parasitic fungi. AFLP study on single spores

54
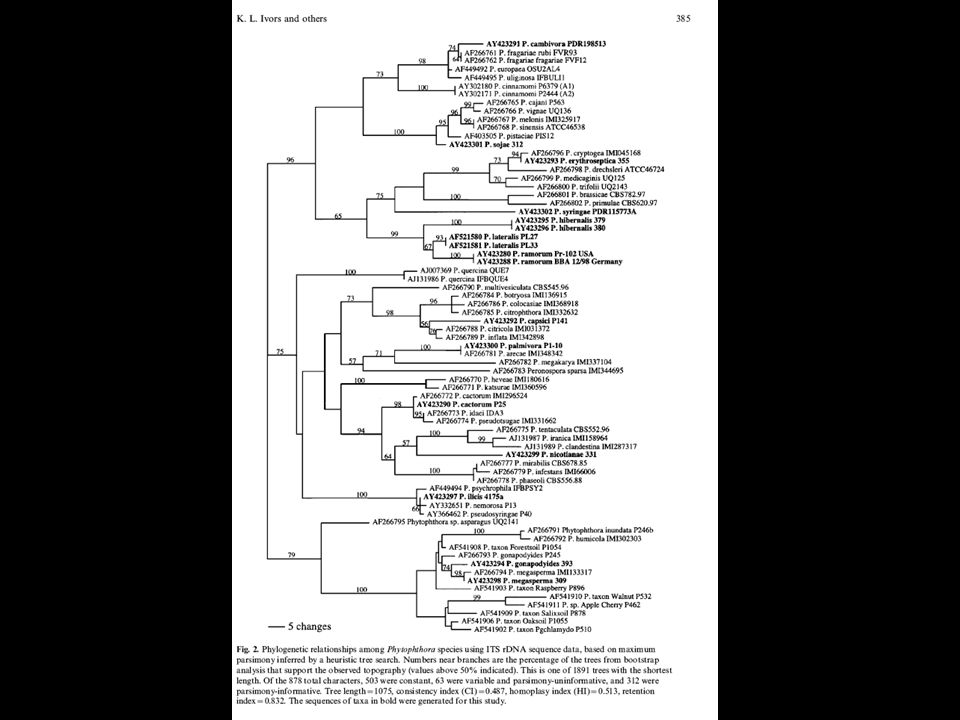
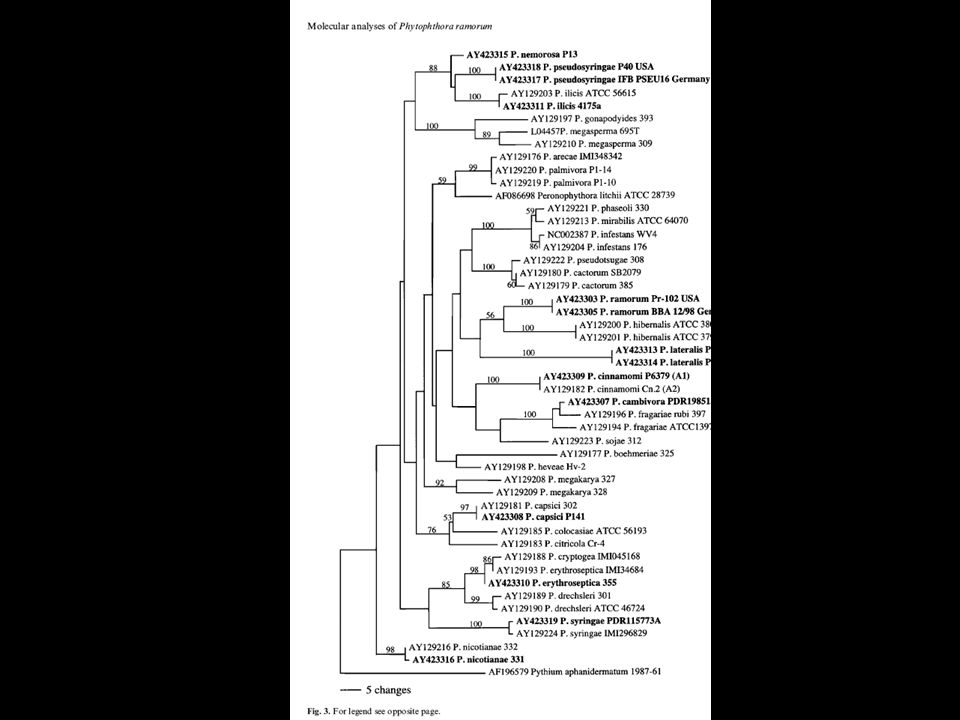
Using DNA sequences Obtain sequence Align sequences, number of parsimony informative sites Gap handling Picking sequences (order) Analyze sequences (similarity/parsimony/exhaustive/bayesian Analyze output; CI, HI Bootstrap/decay indices
 Analyze sequences (similarity/parsimony/exhaustive/bayesian Analyze output; CI, HI Bootstrap/decay indices")
55
Using DNA sequences Testing alternative trees: kashino hasegawa Molecular clock Outgroup Spatial correlation (Mantel) Networks and coalescence approaches
 Networks and coalescence approaches")
56
Pacifico Caribe

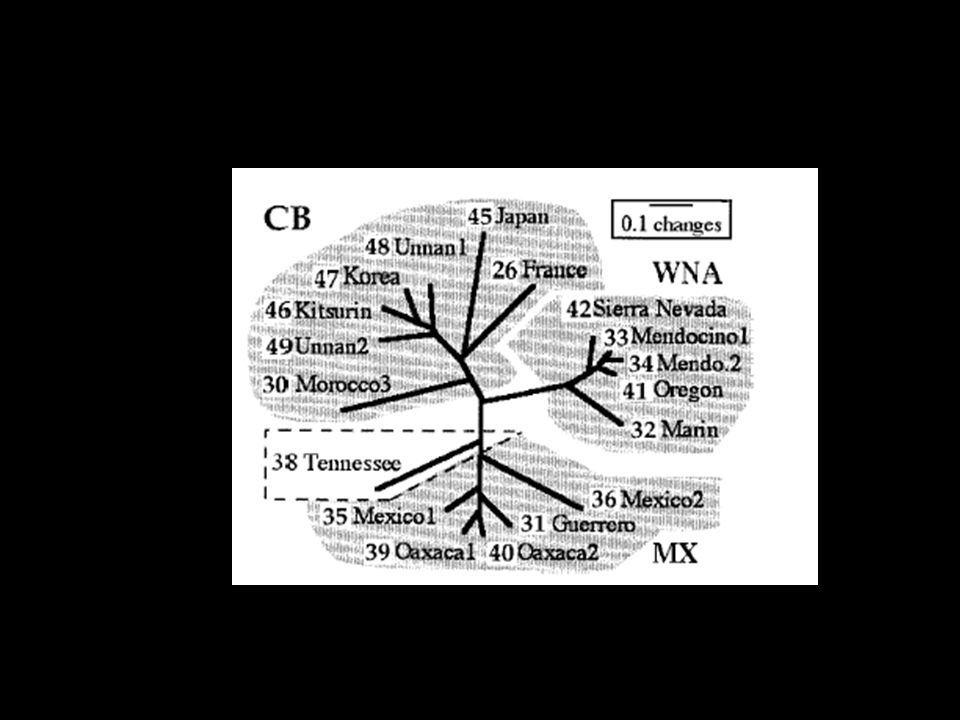
61
From Garbelotto and Chapela, Evolution and biogeography of matsutakes Biodiversity within species as significant as between species

Similar presentations








>")
 Molecular Medicine 2) Energy sources and environmental applications 3) Risk assessment 4) Bioarchaeology,>")






 The order of the base pairs in the sequence of every human varies In a single.>")



 Known as the Humungous Fungus, or honey mushroom Form rhizomorphs, which make up much of the “humungous” part Basidiocarp:>")
 Known as the Humungous Fungus, or honey mushroom Form rhizomorphs, which make up much of the “humungous” part Basidiocarp:>")





