Download presentation
Presentation is loading. Please wait.

1
THE CHROMOSOMAL BASIS OF INHERITANCE
CHAPTER 15 THE CHROMOSOMAL BASIS OF INHERITANCE

2
I. Concept 15.1: Behavior of Chromosomes
A. Background 1. Genetics 1860’s—Mendel proposed that discrete inherited factors segregate and assort independently during gamete formation 2. Cytology 1875—Cytologists worked out process of mitosis 1890’s—Cytologists worked out process of meiosis 3. Genetics 1900—Correns, von Tschermak, and de Vries independently discovered Mendel’s work

3
4. Cytology and Genetics 1902—2 areas converge as Walter Sutton, Theodor Boveri, and others noticed parallels between the behavior of Mendel’s factors and the behavior of chromosomes: Chromosomes and genes both present in pairs in diploid cells. Homologous chromosomes separate and allele pairs segregate during meiosis. Fertilization restores the diploid condition for both. 5. Chromosomal Theory of Inheritance is based on these observations. According to this theory: Mendelian factors (genes) are located on chromosomes. Chromosomes segregate and assort independently during meiosis
 are located on chromosomes. Chromosomes segregate and assort independently during meiosis.")
4
Here we correlate the results of one of Mendel’s dihybrid crosses with the behavior of chromosomes during meiosis. The arrangement of chromosomes at metaphase I of meiosis and their movement during anaphase I account for the segregation and independent assortment of the alleles for seed color and shape. Each cell that undergoes meiosis in an F1 plant produces 2 kinds of gametes. If we count the results for all cells, however, each F1 plant produces equal numbers of all four kinds of gametes because the alternative chromosome arrangements at metaphase I are equally likely.

5
B. Contributions to the Chromosomal Theory
1. Walter Flemming (1882) First to observe chromosomes in nuclei of dividing salamander cells Called process “mitosis” 2. August Weismann (1887) Each gamete has half the number of chromosomes as a fertilized egg. Proposed that a special division process reduced the chromosome number by one half
 First to observe chromosomes in nuclei of dividing salamander cells. Called process mitosis 2. August Weismann (1887) Each gamete has half the number of chromosomes as a fertilized egg. Proposed that a special division process reduced the chromosome number by one half.")
6
Discovered the centriole Actually observed meiosis in cells of Ascaris
3. Theodore Boveri (1888) Determined that chromosomes were essential for fertilization and development Discovered the centriole Actually observed meiosis in cells of Ascaris 4. Walter Sutton (1902) Found relationship between meiosis and Mendel’s laws Predicted gene linkage Following Mendel’s studies with peas, another biologist by the name of Thomas Hunt Morgan began his studies on fruit flies (drosophila melanogaster) ie: red eyes in drosophila (w+)- a superscript + identifies the allele for the wild-type trait. ie: white eyes in drosophila- these are assumed to have originated as changes or mutations in the wild type allele (w)
 Determined that chromosomes were essential for fertilization and development. Discovered the centriole. Actually observed meiosis in cells of Ascaris. 4. Walter Sutton (1902) Found relationship between meiosis and Mendel’s laws. Predicted gene linkage. Following Mendel’s studies with peas, another biologist by the name of Thomas Hunt Morgan began his studies on fruit flies (drosophila melanogaster) ie: red eyes in drosophila (w+)- a superscript + identifies the allele for the wild-type trait. ie: white eyes in drosophila- these are assumed to have originated as changes or mutations in the wild type allele (w)")
7
Began experiments with Drosophila melanogaster (fruit fly)
5. Thomas Hunt Morgan (1910) Began experiments with Drosophila melanogaster (fruit fly) Discovered X-linked (sex linked) inheritance Discovered sex determination (X and Y chromosomes) Provided convincing evidence that Mendel’s factors are located on chromosomes Following Mendel’s studies with peas, another biologist by the name of Thomas Hunt Morgan began his studies on fruit flies (drosophila melanogaster) ie: red eyes in drosophila (w+)- a superscript + identifies the allele for the wild-type trait. ie: white eyes in drosophila- these are assumed to have originated as changes or mutations in the wild type allele (w)
 Began experiments with Drosophila melanogaster (fruit fly) Discovered X-linked (sex linked) inheritance. Discovered sex determination (X and Y chromosomes) Provided convincing evidence that Mendel’s factors are located on chromosomes. Following Mendel’s studies with peas, another biologist by the name of Thomas Hunt Morgan began his studies on fruit flies (drosophila melanogaster) ie: red eyes in drosophila (w+)- a superscript + identifies the allele for the wild-type trait. ie: white eyes in drosophila- these are assumed to have originated as changes or mutations in the wild type allele (w)")
8
C. Morgan and Drosophila
1. Reasons for using Drosophila Easy to raise Produce a large number of offspring Short life cycle (2 weeks) Mutants easily recognized Only 4 pairs of chromosomes (3 pr. of autosomes, 1 pr. of sex chromosomes XX female, XY male)
 Mutants easily recognized. Only 4 pairs of chromosomes (3 pr. of autosomes, 1 pr. of sex chromosomes XX female, XY male)")
9
2. Sex-linked Traits Wild type—normal character phenotype Ex: w+-red eyes Mutant phenotype -alternatives to wild type -due to mutations in wild type gene Ex: w-white eyes Discovered by Morgan Refers to genes on the X chromosome From this experiment, Morgan deduced that: a. Eye color is linked to sex. b. The gene for eye color is located on the X chromosome.

11
EXPERIMENT RESULTS P Generation F1 All offspring Generation
Fig. 15-4b EXPERIMENT P Generation F1 All offspring had red eyes Generation RESULTS F2 Generation When Morgan mated his white-eyed male fly with a red-eyed female, all the F1 offspring had red eyes, suggesting that the wild-type allele is dominant. When Morgan bred the F1 flies to one another, he observed the classical 3:1 phenotypic ratio among the F2 offspring. However, there was a surprising additional result: the white-eye trait showed up only in males. All the F2 females had red eyes, while half of the males had red eyes and half had white eyes. Therefore, Morgan concluded that somehow a fly’s eye color was linked to its sex. (If the eye-color were unrelated to sex, one would have expected half of the white-eyed flies to be male and half female). Recall that a female fly is XX while the male is XY. The correlation between the trait of white eye color and the male sex of the affected F2 flies suggested to Morgan that the gene involved in his white-eyed mutant was located exclusively on the X chromosome, with no corresponding allele present on the Y chromosome. Morgan’s finding of the correlation between a particular trait and an individual’s sex provided support for the chromosome theory of inheritance, namely that a specific gene is carried on a specific chromosome (in this case, an eye-color gene on the X chromosome).
. Recall that a female fly is XX while. the male is XY. The correlation between the trait of white eye color and the male sex of the affected F2 flies suggested to Morgan that the. gene involved in his white-eyed mutant was located exclusively on the X chromosome, with no corresponding allele present on the Y. chromosome. Morgan’s finding of the correlation between a particular trait and an individual’s sex provided support for the chromosome. theory of inheritance, namely that a specific gene is carried on a specific chromosome (in this case, an eye-color gene on the X chromosome).")
12
Experiment:

13
3. Morgan’s finding of the correlation between a particular trait
3. Morgan’s finding of the correlation between a particular trait and an individual’s sex provided support for the chromosome theory of inheritance

14
II. Concept 15.2 Sex-Linked Genes
Heterogametic sex—produces 2 kinds of gametes which determine the sex of offspring Homogametic sex—produces 1 kind of gamete with respect to sex chromosome In humans and some other animals, there is a chromosomal basis of sex determination Each gamete has only 1 sex chromosome.

15
A. The Chromosomal Basis of Sex
In humans and other mammals, there are two varieties of sex chromosomes: a larger X chromosome and a smaller Y chromosome Only the ends of the Y chromosome have regions that are homologous with the X chromosome In both testes and ovaries, the two sex chromosomes segregate during meiosis, and each gamete receives one. Each egg contains one X chromosome. In contrast, sperm fall into two categories: half the sperm cells a male produces contain an X chromosome and the other half contain a Y chromosome. At the time of conception, sex is formed (whether an X sperm fertilizes the egg to form a female, or the Y sperm fertilizes the egg to form a male). Thus sex determination is a matter of chance. In humans the anatomical signs of sex begin to emerge when the embryo is about 2 months old. Before then the gonads are generic, meaning they can develop in either testes or ovaries, depending on whether a Y chromosome is present. In 1990, the gene on the Y chromosome required for the development of testes was identified. They named the gene SRY, for Sex-determining Region of Y.
. Thus sex determination is a matter of chance. In humans the anatomical signs of sex begin to emerge when the embryo is about 2 months old. Before then the gonads are generic, meaning they can develop in either testes or ovaries, depending on whether a Y chromosome is present. In 1990, the gene on the Y chromosome required for the development of testes was identified. They named the gene SRY, for Sex-determining Region of Y.")
16
The Chromosomal Basis of Sex
3. The SRY gene on the Y chromosome codes for the development of testes SRY- Sex determining Region of Y -The presence of this gene on the Y chromosome codes for the development of testes. In the absence of this gene, the gonads develop into ovaries In both testes and ovaries, the two sex chromosomes segregate during meiosis, and each gamete receives one. Each egg contains one X chromosome. In contrast, sperm fall into two categories: half the sperm cells a male produces contain an X chromosome and the other half contain a Y chromosome. At the time of conception, sex is formed (whether an X sperm fertilizes the egg to form a female, or the Y sperm fertilizes the egg to form a male). Thus sex determination is a matter of chance. In humans the anatomical signs of sex begin to emerge when the embryo is about 2 months old. Before then the gonads are generic, meaning they can develop in either testes or ovaries, depending on whether a Y chromosome is present. In 1990, the gene on the Y chromosome required for the development of testes was identified. They named the gene SRY, for Sex-determining Region of Y.
. Thus sex determination is a matter of chance. In humans the anatomical signs of sex begin to emerge when the embryo is about 2 months old. Before then the gonads are generic, meaning they can develop in either testes or ovaries, depending on whether a Y chromosome is present. In 1990, the gene on the Y chromosome required for the development of testes was identified. They named the gene SRY, for Sex-determining Region of Y.")
17
B. Systems of Sex Determination
1. X-Y Mammals, humans, Drosophila ♂--heterogametic (XY) ♀--homogametic (XX) Y chromosome determines sex of offspring 2. X-O Grasshopper, cricket, roach, and other insects ♂--heterogametic (XO)
 ♀--homogametic (XX) Y chromosome determines sex of offspring. 2. X-O. Grasshopper, cricket, roach, and other insects. ♂--heterogametic (XO)")
18
3. Z-W Birds, some fish, some insects—butterflies and moths ♀--heterogametic (ZW) ♂--homogametic (ZZ) 4. Haplo-diploidy Bees and ants Have no sex chromosomes ♀--develops from fertilized egg (2n) ♂--develops parthenogenetically from unfertilized egg (n)
 ♂--develops parthenogenetically from unfertilized egg (n)")
20
C. Inheritance of Sex-Linked Genes
The sex chromosomes have genes for many characters unrelated to sex A gene located on the X chromosome is called a sex-linked gene Genes on the Y chromosome are called holandric genes and are found in males only In humans, sex-linked usually refers to a gene on the larger X chromosome Sex-linked genes follow specific patterns of inheritance In addition to their role as carriers of genes that determine sex, the sex chromosomes, especially X chromosomes, have genes for many characters unrelated to sex. A gene located on either sex chromosome is called a sex-linked gene, although in humans the term has historically refereed specifically to a gene on the X chromosome. Fathers pass sex-linked alleles to all of their daughters but to none of their sons. In contrast, mothers can pass sex-linked alleles to both sons and daughters (see diagram). If a sex-linked trait is due to a recessive allele, a female will express the phenotype only if she is a homozygote. Because males have only one locus, the terms homozygous and heterozygous lack meaning for describing their sex-linked genes; the term hemizygous is used in such cases. Any male receiving the recessive allele from his mother will express the trait. For this reason, far more males than females have sex-linked recessive disorders. However, even though the chance of a female inheriting a double dose of the mutant allele is much less than the probability of a male inheriting a single dose, there are females with sex-linked disorders. For instance, color blindness is a mild disorder inherited as a sex-linked trait. A color-blind daughter may be born to a color-blind father whose mate is a carrier. Because the sex-linked allele for color blindness is relatively rare, though, the probability that such a man and woman will mate is low. Only the ends of the Y chromosome have regions that are homologous with the X chromosome Have students complete the following crosses: XNXN x XnY, XNXn x XNY, XNXn x XnY (see next slide for actual crosses) N= normal color vision, n=colorblind
. If a sex-linked trait is due to a recessive allele, a female will express the phenotype only if she is a homozygote. Because males have only one locus, the terms homozygous and heterozygous lack meaning for describing their sex-linked genes; the term hemizygous is used in such cases. Any male receiving the recessive allele from his mother will express the trait. For this reason, far more males than females have sex-linked recessive disorders. However, even though the chance of a female inheriting a double dose of the mutant allele is much less than the probability of a male inheriting a single dose, there are females with sex-linked disorders. For instance, color blindness is a mild disorder inherited as a sex-linked trait. A color-blind daughter may be born to a color-blind father whose mate is a carrier. Because the sex-linked allele for color blindness is relatively rare, though, the probability that such a man and woman will mate is low. Only the ends of the Y chromosome have regions that are homologous with the X chromosome. Have students complete the following crosses: XNXN x XnY, XNXn x XNY, XNXn x XnY (see next slide for actual crosses) N= normal color vision, n=colorblind.")
21
6. For a recessive sex-linked trait to be expressed:
A female needs two copies of the allele A male needs only one copy of the allele 7. Sex-linked recessive disorders are much more common in males than in females 8. If a sex-linked trait is due to a recessive allele, a female will express the trait only if she is homozygous. 9. A heterozygous female is a carrier. 10. Males only need 1 allele of a sex-linked trait to show the trait. 11. Males are hemizygous (only one copy of a gene is present in a diploid organism)
")
22
N= normal color vision XNXN XnY XNXn XNY XNXn XnY Fig. 15-7
A number of human sex-linked disorders caused by recessive alleles are much more serious than color blindness, such as Duchenne muscular dystrophy. In this disorder, there is an absence of a key muscle protein which is located on the X chromosome. Hemophilia is another example. Since female mammals, including humans, inherit two X chromosomes, you may wonder whether females make twice as much of the proteins encoded by genes on the X chromosome, compared to the amounts in males. In fact, one X chromosome in each cell in females becomes almost completely inactivated during embryonic development. As a result, the cells of females and males have the same effective does (one copy) of these genes. The inactive X in each cell of a female condenses into a compact object called a Barr body, which lies along the inside of the nuclear envelope. Most of the genes of the X chromosome that forms the Barr body are not expressed. In the ovaries, Barr body chromosomes are reactivated in the cells that give rise to eggs, so every female gamete has an active X. Selection of which X chromosome becomes inactivated and forms the Barr body occurs randomly and independently, therefore females consist of a mosaic of two types of cells: an active X derived from their mother, or an active X derived from their father. After an X chromosome is inactivated in a particular cell, all mitotic descendants of that cell have the same inactive X. Thus, if a female is heterozygous for a sex-linked trait, about half her cells will express one allele, while the others will express the alternate allele. In humans, mosaicism can be observed in a recessive X-linked mutation that prevents the development of sweat glands. A woman who is heterozygous for this trait has patches of normal skin and patches of skin lacking sweat glands.
 of these genes. The inactive X in each cell of a female condenses into a compact object called a Barr body, which lies along the inside of the nuclear envelope. Most of the genes of the X chromosome that forms the Barr body are not expressed. In the ovaries, Barr body chromosomes are reactivated in the cells that give rise to eggs, so every female gamete has an active X. Selection of which X chromosome becomes inactivated and forms the Barr body occurs randomly and independently, therefore females consist of a mosaic of two types of cells: an active X derived from their mother, or an active X derived from their father. After an X chromosome is inactivated in a particular cell, all mitotic descendants of that cell have the same inactive X. Thus, if a female is heterozygous for a sex-linked trait, about half her cells will express one allele, while the others will express the alternate allele. In humans, mosaicism can be observed in a recessive X-linked mutation that prevents the development of sweat glands. A woman who is heterozygous for this trait has patches of normal skin and patches of skin lacking sweat glands.")
23
12. Fathers pass sex-linked alleles to only and all of their daughters
XA—Normal gene Xa—Sex-linked gene XAXA—Normal Female XAXa—Carrier Female XaXa—Affected Female XAY—Normal Male XaY—Affected Male

24
13. Mothers can pass sex- linked alleles to both sons and daughters.

25
14. If a carrier mates with a male who has the disorder, there is a 50% chance that each child born to them will have the disorder, regardless of sex. Daughters who do not have the disorder will be carriers, whereas males without the disorder will be completely free of the recessive allele

26
15. Examples of Sex-linked Traits
a. Color Blindness -recessive -can’t distinguish certain colors -red-green most common b. Duchenne’s Muscular Dystrophy -atrophy of muscle c. Hemophilia -blood fails to clot because of lack of a clotting factor

27
D. X-inactivation in Females
1. Female mammals have only one fully functional X chromosome in diploid cells. 2. Proposed by Mary F. Lyon and known as the Lyon Hypothesis 3. Each of the embryonic cells inactivates one of the two X chromosomes. 4. Inactive X contracts into densely staining object called a Barr Body. 5. Ex: Mosaic coloration of calico cats Normal sweat gland development in humans

28
X-Inactivation in Females

29
E. Other Disorders Associated with Sex
1. Sex-limited Traits Appear exclusively in one sex Ex: uterine cancer in ♀; prostate cancer in ♂ 2. Sex-influenced Traits Expression is influenced by presence of ♀ or ♂ sex hormones Acts as a dominant in one sex and recessive in the other Ex: ♂--baldness, stomach ulcers; ♀--breast cancer both—length of index finger as compared to ring finger -dominant in ♂ shorter index finger -dominant in ♀ longer index finger

30
III. Concept 15.3: Linked Genes
Each chromosome has hundreds or thousands of genes Genes located on the same chromosome that tend to be inherited together are called linked genes Do not assort independently Dihybrid crosses deviate from expected 9:3:3:1 phenotypic ratio 1. The number of genes in a cell is far greater than the number of chromosomes; in fact, each chromosome has hundreds or thousands of genes. (The Y chromosome only has 78 identified so far, although more may continue to be found). Note the distinction between the terms sex-linked gene, referring to a single gene on a sex chromosome, and linked genes, referring to two or more genes on the same chromosome that tend to be inherited together. See next slide for diagram of Morgan’s experiment. 2.
. Note the distinction between the terms sex-linked gene, referring to a single gene on a sex chromosome, and linked genes, referring to two or more genes on the same chromosome that tend to be inherited together. See next slide for diagram of Morgan’s experiment. 2.")
31
A. Genetic Recombination
1. Production of offspring with new combinations of traits different from those combinations found in the parents (crossing over) 2. Results from the events of meiosis and random fertilization B. Recombination of Unlinked Genes: Independent Assortment Parental types—progeny that have the same phenotype as one or the other of the parents Recombinant types—progeny whose phenotypes differ from either parent
 2. Results from the events of meiosis and random fertilization. B. Recombination of Unlinked Genes: Independent Assortment. Parental types—progeny that have the same phenotype as one or the other of the parents. Recombinant types—progeny whose phenotypes differ from either parent.")
32
YR yr Yr yR yr YyRr yyrr Yyrr yyRr
Parental Types- offspring that inherit a phenotype that matches one of the parental phenotypes. Gametes from yellow-round heterozygous parent (YyRr) YR yr Yr yR Gametes from green- wrinkled homozygous recessive parent ( yyrr) yr YyRr yyrr Yyrr yyRr Mendel learned from his crosses that some offspring have combinations of traits that do not match those of either parent. Notice in this punnett square that one-half of the offspring are expected to inherit a phenotype that matches one of the parental phenotypes, but two nonparental phenotypes are also found in the offspring. When 50% of all offspring are recombinants, as in this example, geneticists say that there is a 50% frequency of recombination. A 50% frequency or recombination in such testcrosses is observed for any two genes that are located on different chromosomes and are thus unlinked. The physical basis of recombination between unlinked genes is the random orientation of homologous chromosomes at metaphase I of meiosis, which leads to the independent assortment of the two unlinked genes. Thinking back to Morgan’s color and wing size cross of the fruit flies, we can conclude that the two genes were on the same chromosome since the occurrence of the parental types with a frequency greater than 50% indicates that the genes are linked. This experiment exhibited ~17% recombinants. Because of these results, Morgan proposed that some process must occasionally break the physical connection between specific alleles of genes on the same chromosome. Subsequent experiments demonstrated that this process, now called crossing over, accounts for the recombination of link genes Parental- type offspring Recombinant offspring Recombinant Types or Recombinants- offspring that have new combinations of phenotypes.
 YR. yr. Yr. yR. Gametes from green- wrinkled homozygous. recessive parent ( yyrr) yr. YyRr. yyrr. Yyrr. yyRr. Mendel learned from his crosses that some offspring have combinations of traits that do not match those of either parent. Notice in this punnett square that one-half of the offspring are expected to inherit a phenotype that matches one of the parental phenotypes, but two nonparental phenotypes are also found in the offspring. When 50% of all offspring are recombinants, as in this example, geneticists say that there is a 50% frequency of recombination. A 50% frequency or recombination in such testcrosses is observed for any two genes that are located on different chromosomes and are thus unlinked. The physical basis of recombination between unlinked genes is the random orientation of homologous chromosomes at metaphase I of meiosis, which leads to the independent assortment of the two unlinked genes. Thinking back to Morgan’s color and wing size cross of the fruit flies, we can conclude that the two genes were on the same chromosome since the occurrence of the parental types with a frequency greater than 50% indicates that the genes are linked. This experiment exhibited ~17% recombinants. Because of these results, Morgan proposed that some process must occasionally break the physical connection between specific alleles of genes on the same chromosome. Subsequent experiments demonstrated that this process, now called crossing over, accounts for the recombination of link genes. Parental- type. offspring. Recombinant. offspring. Recombinant Types or Recombinants- offspring that have new. combinations of phenotypes.")
33
P Yy Rr x yyrr (testcross)
(yellow, round) (green, wrinkled) F1 ¼ YyRr ¼ yyrr Parental types % ¼ yyRr ¼ Yyrr Recombinant types 50% When half the progeny are recombinants, there is a 50% frequency of recombination. A 50% frequency of recombination usually indicates that the two genes are on different chromosomes, because it is the expected result if the two genes assort randomly.
 (green, wrinkled) F1 ¼ YyRr ¼ yyrr Parental types 50% ¼ yyRr ¼ Yyrr Recombinant types 50% When half the progeny are recombinants, there is a 50% frequency of recombination. A 50% frequency of recombination usually indicates that the two genes are on different chromosomes, because it is the expected result if the two genes assort randomly.")
34
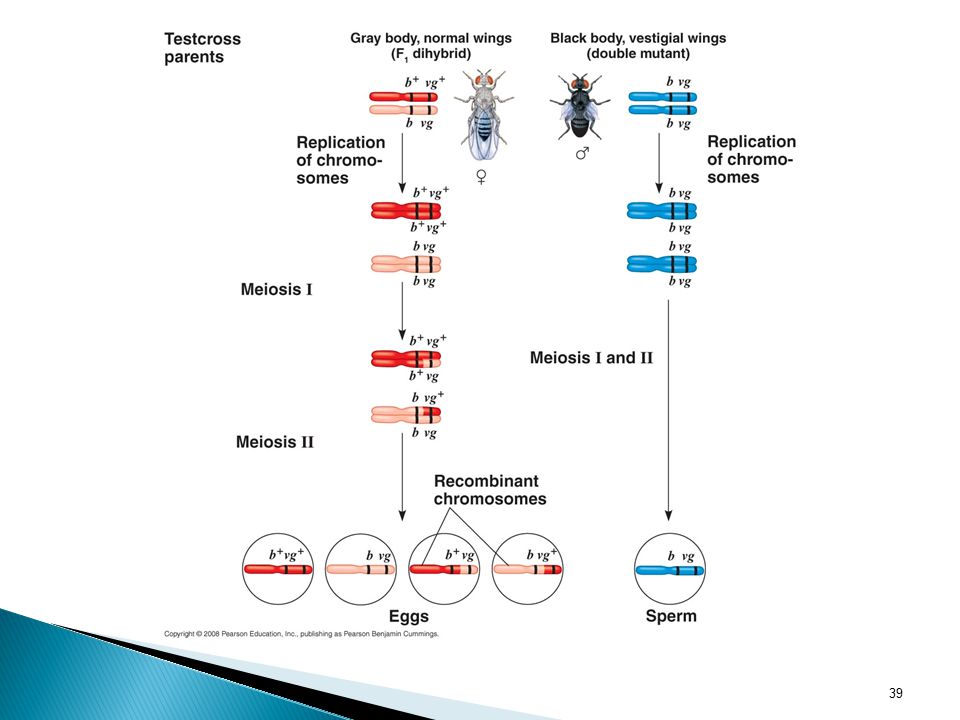
b+--gray body vg+--wild type wings b+ b vg+ vg x b b vg vg (testcross)
C. Recombination of Linked Genes: Crossing Over Crossing Over- accounts for the recombination of linked genes. b—black body vg—vestigial wings b+--gray body vg+--wild type wings b+ b vg+ vg x b b vg vg (testcross) (gray, normal wings) (black, vestigial wings) The discovery of linked genes and recombination due to crossing over led one of Morgan’s students, Sturtevant, to a method for constructing a genetic map, an ordered list of the genetic loci along a particular chromosome. He hypothesized that recombination frequencies depend on the distances between genes on a chromosome. He assumed that crossing over is a random event, with the chance equal at all points along a chromosome. Based on these assumptions, he predicted that he farther apart two genes are, the higher the probability that a crossover will occur between them and therefore the higher the recombination frequency. His reasoning was simple: the greater the distance between two genes, the more points there are between them where crossing over can occur. Using recombinant data from various fruit fly crosses, Sturtevant proceeded to assign relative positions to genes on the same chromosomes- that is, to map genes. The map depicted here shows three genes: body color (b), cinnabar gene which affects eye color (cn), and wing size (vg). The recombination frequency between cn and b is 9%, between cn and vg is 9.5% and between b and vg is 17%. In other words, crossovers between cn and b and between cn and vg are about half as frequent as crossovers between b and vg. Only a map that locates cn about midway between b and vg is consistent with this data. Sturtevant expressed the distances in map units equal to 1% recombination. Today map units are often called centimorgans in honor of Morgan. In practice, the interpretation of recombination data is more complicated than this example suggests. For example, some genes on a chromosome are so far from each other that a crossover between them is virtually certain. The observed frequency of recombination in crosses involving two such genes can have a maximum value of 50%, a result indistinguishable from that for genes on different chromosomes. In this case, on a chromosome are mapped by adding the recombination frequencies from crosses involving a set of closer pairs of genes lying between the two different genes. Other methods enable geneticists to construct cytogenic maps of chromosomes, which locate genes with respect to chromosomal features such as stained bands, that can be see in the microscope. The ultimate maps, the genome, shows the physical distances between gene loci in DNA nucleotides. (to be discussed in detail later) Genome- the genetic material of an organism or virus; the complete complement of an organism’s or virus’s genes along with its noncoding nucleic acid sequences.
 (gray, normal wings) (black, vestigial wings) The discovery of linked genes and recombination due to crossing over led one of Morgan’s students, Sturtevant, to a method for constructing a genetic map, an ordered list of the genetic loci along a particular chromosome. He hypothesized that recombination frequencies depend on the distances between genes on a chromosome. He assumed that crossing over is a random event, with the chance equal at all points along a chromosome. Based on these assumptions, he predicted that he farther apart two genes are, the higher the probability that a crossover will occur between them and therefore the higher the recombination frequency. His reasoning was simple: the greater the distance between two genes, the more points there are between them where crossing over can occur. Using recombinant data from various fruit fly crosses, Sturtevant proceeded to assign relative positions to genes on the same chromosomes- that is, to map genes. The map depicted here shows three genes: body color (b), cinnabar gene which affects eye color (cn), and wing size (vg). The recombination frequency between cn and b is 9%, between cn and vg is 9.5% and between b and vg is 17%. In other words, crossovers between cn and b and between cn and vg are about half as frequent as crossovers between b and vg. Only a map that locates cn about midway between b and vg is consistent with this data. Sturtevant expressed the distances in map units equal to 1% recombination. Today map units are often called centimorgans in honor of Morgan. In practice, the interpretation of recombination data is more complicated than this example suggests. For example, some genes on a chromosome are so far from each other that a crossover between them is virtually certain. The observed frequency of recombination in crosses involving two such genes can have a maximum value of 50%, a result indistinguishable from that for genes on different chromosomes. In this case, on a chromosome are mapped by adding the recombination frequencies from crosses involving a set of closer pairs of genes lying between the two different genes. Other methods enable geneticists to construct cytogenic maps of chromosomes, which locate genes with respect to chromosomal features such as stained bands, that can be see in the microscope. The ultimate maps, the genome, shows the physical distances between gene loci in DNA nucleotides. (to be discussed in detail later) Genome- the genetic material of an organism or virus; the complete complement of an organism’s or virus’s genes along with its noncoding nucleic acid sequences.")
35
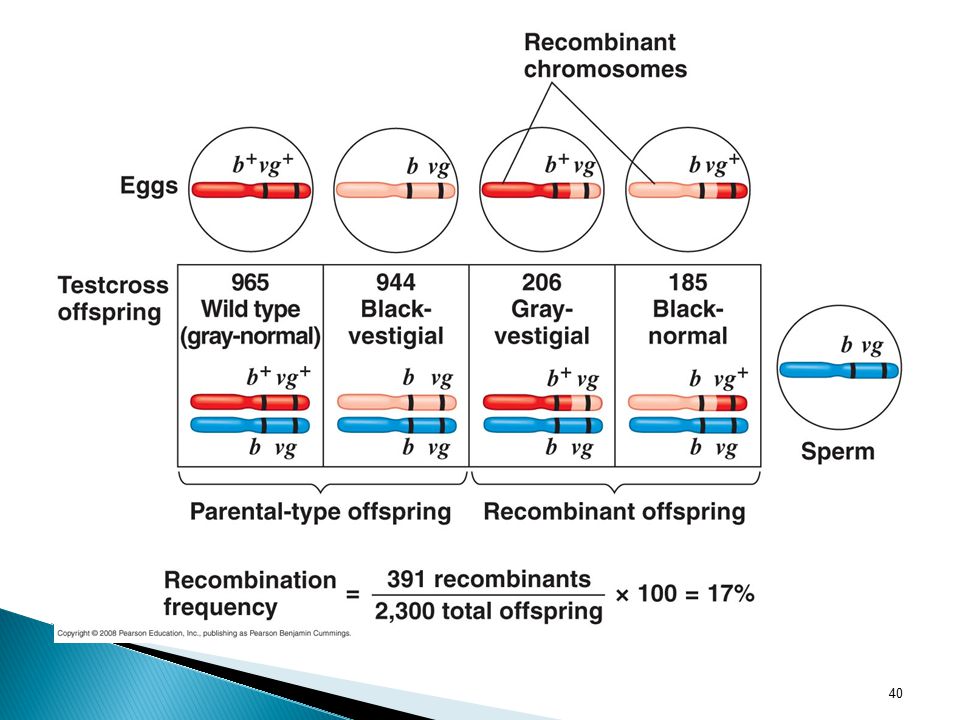
Possible Results b vg Pheno-types Expected if genes not linked
Expected if genes totally linked Actual b vg+ b b vg+vg black normal 575 206 b+ vg+ b+ b vg+vg gray 1150 965 b b vg vg vestigial 944 b+ vg b+ b vg vg 185

37
What type of ratios would you expect to see in the testcross
Answer: 1:1:1:1 Answer: 1:1:0:0 The ratios turned out to be 965:944:206:185. In these crosses, Morgan observed a much higher proportion of parental phenotypes than would be expected if the two genes assorted independently. Based on these results, he concluded that body color and wing size are usually inherited together in specific combinations (the parental combinations) because the genes for these characters are on the same chromosomes. Because both of the non-parental phenotypes were also produced, the study suggested that the two traits are only partially linked genetically. To understand this conclusion, we need to explore genetic recombination. What type of ratios would you expect to see in the testcross offspring if the genes were located on different chromosomes? What if they were located on the same chromosome and parental alleles are always inherited together?
 because the genes for these characters are on the same chromosomes. Because both of the non-parental phenotypes were also produced, the study suggested that the two traits are only partially linked genetically. To understand this conclusion, we need to explore genetic recombination. What type of ratios would you expect to see in the testcross. offspring if the genes were located on different chromosomes What if they were located on the same chromosome and parental. alleles are always inherited together")
41
Recombination Frequency (RF) = # recombinants x 100
total # offspring Proposed by Morgan Process of crossing over during meiosis accounts for the recombination of linked genes (genes on same chromosome) Crossing over—breakage and exchange of corresponding segments between homologous chromosomes --results in new allelic combination Probability of crossing over (recombination) between two genes is proportional to the distance separating those genes The closer together two genes are, the less likely that a cross over will occur. Proved by A. H. Sturtevant
 Crossing over—breakage and exchange of corresponding segments between homologous chromosomes. --results in new allelic combination. Probability of crossing over (recombination) between two genes is proportional to the distance separating those genes. The closer together two genes are, the less likely that a cross over will occur. Proved by A. H. Sturtevant.")
42
D. Chromosome Mapping 1. Recombination frequencies are used to construct chromosome maps [show locations of genes on a particular chromosome (linear)] 2. Sturtevant constructed chromosome maps for Drosophila using recombination frequencies 3. 1 map unit = 1% recombination frequency (now called centimorgans)
] 2. Sturtevant constructed chromosome maps for Drosophila using recombination frequencies 3. 1 map unit = 1% recombination frequency (now called centimorgans)")
43
Recombination Genetic Map- an ordered list of the genetic loci along a particular chromosome. Linkage Map- a genetic map based on recombination frequencies. Displays order but not precise location. Distances are expressed in map units- equivalent to 1% recombination frequency. (Centimorgans) The discovery of linked genes and recombination due to crossing over led one of Morgan’s students, Sturtevant, to a method for constructing a genetic map, an ordered list of the genetic loci along a particular chromosome. He hypothesized that recombination frequencies depend on the distances between genes on a chromosome. He assumed that crossing over is a random event, with the chance equal at all points along a chromosome. Based on these assumptions, he predicted that he farther apart two genes are, the higher the probability that a crossover will occur between them and therefore the higher the recombination frequency. His reasoning was simple: the greater the distance between two genes, the more points there are between them where crossing over can occur. Using recombinant data from various fruit fly crosses, Sturtevant proceeded to assign relative positions to genes on the same chromosomes- that is, to map genes. The map depicted here shows three genes: body color (b), cinnabar gene which affects eye color (cn), and wing size (vg). The recombination frequency between cn and b is 9%, between cn and vg is 9.5% and between b and vg is 17%. In other words, crossovers between cn and b and between cn and vg are about half as frequent as crossovers between b and vg. Only a map that locates cn about midway between b and vg is consistent with this data. Sturtevant expressed the distances in map units equal to 1% recombination. Today map units are often called centimorgans in honor of Morgan. In practice, the interpretation of recombination data is more complicated than this example suggests. For example, some genes on a chromosome are so far from each other that a crossover between them is virtually certain. The observed frequency of recombination in crosses involving two such genes can have a maximum value of 50%, a result indistinguishable from that for genes on different chromosomes. In this case, on a chromosome are mapped by adding the recombination frequencies from crosses involving a set of closer pairs of genes lying between the two different genes. Other methods enable geneticists to construct cytogenic maps of chromosomes, which locate genes with respect to chromosomal features such as stained bands, that can be see in the microscope. The ultimate maps, the genome, shows the physical distances between gene loci in DNA nucleotides. (to be discussed in detail later) Genome- the genetic material of an organism or virus; the complete complement of an organism’s or virus’s genes along with its noncoding nucleic acid sequences.
 The discovery of linked genes and recombination due to crossing over led one of Morgan’s students, Sturtevant, to a method for constructing a genetic map, an ordered list of the genetic loci along a particular chromosome. He hypothesized that recombination frequencies depend on the distances between genes on a chromosome. He assumed that crossing over is a random event, with the chance equal at all points along a chromosome. Based on these assumptions, he predicted that he farther apart two genes are, the higher the probability that a crossover will occur between them and therefore the higher the recombination frequency. His reasoning was simple: the greater the distance between two genes, the more points there are between them where crossing over can occur. Using recombinant data from various fruit fly crosses, Sturtevant proceeded to assign relative positions to genes on the same chromosomes- that is, to map genes. The map depicted here shows three genes: body color (b), cinnabar gene which affects eye color (cn), and wing size (vg). The recombination frequency between cn and b is 9%, between cn and vg is 9.5% and between b and vg is 17%. In other words, crossovers between cn and b and between cn and vg are about half as frequent as crossovers between b and vg. Only a map that locates cn about midway between b and vg is consistent with this data. Sturtevant expressed the distances in map units equal to 1% recombination. Today map units are often called centimorgans in honor of Morgan. In practice, the interpretation of recombination data is more complicated than this example suggests. For example, some genes on a chromosome are so far from each other that a crossover between them is virtually certain. The observed frequency of recombination in crosses involving two such genes can have a maximum value of 50%, a result indistinguishable from that for genes on different chromosomes. In this case, on a chromosome are mapped by adding the recombination frequencies from crosses involving a set of closer pairs of genes lying between the two different genes. Other methods enable geneticists to construct cytogenic maps of chromosomes, which locate genes with respect to chromosomal features such as stained bands, that can be see in the microscope. The ultimate maps, the genome, shows the physical distances between gene loci in DNA nucleotides. (to be discussed in detail later) Genome- the genetic material of an organism or virus; the complete complement of an organism’s or virus’s genes along with its noncoding nucleic acid sequences.")
44
4. How to use XO (crossover) data to construct a chromosome map:
Ex: RF b vg 17% cn b 9% cn vg 9.5% a. Establish the distance between the genes with the highest RF b vg b. Determine RF between third gene and first. cn b c. Consider the two possible placements of the third gene. cn b vg or b cn vg

45
d. Determine the RF between the third gene and the second gene to eliminate the incorrect sequence
b cn vg 17 --correct sequence b-cn-vg

46
5. If linked genes are so far apart on a chromosome that the RF is
5. If linked genes are so far apart on a chromosome that the RF is 50%, they are indistinguishable from unlinked genes that assort independently. Can map such genes if RF can be determined between those two genes and intermediate genes. 6. Maps from XO data give relative positions of linked genes 7. Cytological mapping pinpoints actual location of genes and real distance between them. May differ from XO maps in distance but not sequence.

48
IV. Concept 15.4: Alternations of Chromosome Number or Structure
Large-scale chromosomal alterations often lead to spontaneous abortions (miscarriages) or cause a variety of developmental disorders Meiotic errors and mutagens can cause major chromosomal changes such as altered chromosome numbers or altered chromosomal structure. A. Alterations of Chromosomal Number 1. Nondisjunction In nondisjunction, pairs of homologous chromosomes do not separate normally during meiosis Results in one gamete receiving two of the same type of chromosome (n+1) and the other gamete receiving none (n-1) Large-scale chromosomal alterations often lead to spontaneous abortions (miscarriages) or cause a variety of developmental disorders Ideally, the meiotic spindle distributes chromosomes to daughter cells without error. But there is an occasional mishap, called a nondisjunction. (The other gametes are usually distributed normally). See next slide for diagram If either of the aberrant gametes unites with a normal one at fertilization, the zygote will also have an abnormal number of a chromosome, a condition known as aneuploidy, which may involve more than one chromosome. Fertilization involving a gamete that has no copy of a particular chromosome will lead to a missing chromosome in the zygote (so that the cell has 2n-1 chromosomes). The aneuploid zygote is said to be monosomic for that chromosome. If a chromosome is present in triplicate in the zygote (so that the cell has 2n+1), the aneuploid cell is trosomic for that chromosome. Mitosis will subsequently transmit the anomaly to all embryonic cells. If the organism survives, it usually has a set of traits caused by the abnormal does of the genes associated with the extra or missing chromosome. Down syndrome is an example of trisomy in human (on the 21st chromosome)- to be discussed more later. Nondisjunction can also occur during mitosis. If such an error takes place early in embryonic development, then the aneuploid condition is passed along by mitosis to a large number of cells and is likely to have a substantial effect. Some organisms have more than two complete chromosome sets in all somatic cells. The general term for this chromosomal alteration is polyploidy; the specific terms triploidy (3n) and tetraploidy (4n) indicated three or four chromosomal sets, respectively. One way a triploid cell may arise is by the fertilization of an abnormal diploid egg produced by nondisjunction of all its chromosomes. Tetraploidy could result from the failure of a 2n zygote to divide after replicating its chromosomes. Subsequent normal mitotic division would then produce a 4n embryo. Polyploidy is very common in plants, in fact many of the plants we eat are polyploid. Bananas are triploid and wheat is hexaploid (6n). In the animal kingdom, ployploid species are much less common, although they do occur in fish and amphibians. In general, polyploids are more nearly normal in appearance than aneuploids. One extra (or mission) chromosome apparently disrupts genetic balance more than does an entire extra set of chromosomes. *Each of these are a result of an accident in cell division (meiosis)
 or cause a variety of developmental disorders. Meiotic errors and mutagens can cause major chromosomal changes such as altered chromosome numbers or altered chromosomal structure. A. Alterations of Chromosomal Number. 1. Nondisjunction. In nondisjunction, pairs of homologous chromosomes do not separate normally during meiosis. Results in one gamete receiving two of the same type of chromosome (n+1) and the other gamete receiving none (n-1) Large-scale chromosomal alterations often lead to spontaneous abortions (miscarriages) or cause a variety of developmental disorders. Ideally, the meiotic spindle distributes chromosomes to daughter cells without error. But there is an occasional mishap, called a nondisjunction. (The other gametes are usually distributed normally). See next slide for diagram. If either of the aberrant gametes unites with a normal one at fertilization, the zygote will also have an abnormal number of a chromosome, a condition known as aneuploidy, which may involve more than one chromosome. Fertilization involving a gamete that has no copy of a particular chromosome will lead to a missing chromosome in the zygote (so that the cell has 2n-1 chromosomes). The aneuploid zygote is said to be monosomic for that chromosome. If a chromosome is present in triplicate in the zygote (so that the cell has 2n+1), the aneuploid cell is trosomic for that chromosome. Mitosis will subsequently transmit the anomaly to all embryonic cells. If the organism survives, it usually has a set of traits caused by the abnormal does of the genes associated with the extra or missing chromosome. Down syndrome is an example of trisomy in human (on the 21st chromosome)- to be discussed more later. Nondisjunction can also occur during mitosis. If such an error takes place early in embryonic development, then the aneuploid condition is passed along by mitosis to a large number of cells and is likely to have a substantial effect. Some organisms have more than two complete chromosome sets in all somatic cells. The general term for this chromosomal alteration is polyploidy; the specific terms triploidy (3n) and tetraploidy (4n) indicated three or four chromosomal sets, respectively. One way a triploid cell may arise is by the fertilization of an abnormal diploid egg produced by nondisjunction of all its chromosomes. Tetraploidy could result from the failure of a 2n zygote to divide after replicating its chromosomes. Subsequent normal mitotic division would then produce a 4n embryo. Polyploidy is very common in plants, in fact many of the plants we eat are polyploid. Bananas are triploid and wheat is hexaploid (6n). In the animal kingdom, ployploid species are much less common, although they do occur in fish and amphibians. In general, polyploids are more nearly normal in appearance than aneuploids. One extra (or mission) chromosome apparently disrupts genetic balance more than does an entire extra set of chromosomes. *Each of these are a result of an accident in cell division (meiosis)")
49
Nondisjunction Figure 15.13 Meiotic nondisjunction
Return to previous slide after discussion with this slide

50
A monosomic zygote has only one copy of a particular chromosome
2. Aneuploidy Aneuploidy results from the fertilization of gametes in which nondisjunction occurred Offspring with this condition have an abnormal number of a particular chromosome a chromosomal aberration in which one or more chromosomes are present in extra copies or deficient in number A monosomic zygote has only one copy of a particular chromosome A trisomic zygote has three copies of a particular chromosome Ex: Down’s Syndrome or Trisomy 21 Large-scale chromosomal alterations often lead to spontaneous abortions (miscarriages) or cause a variety of developmental disorders Ideally, the meiotic spindle distributes chromosomes to daughter cells without error. But there is an occasional mishap, called a nondisjunction. (The other gametes are usually distributed normally). See next slide for diagram If either of the aberrant gametes unites with a normal one at fertilization, the zygote will also have an abnormal number of a chromosome, a condition known as aneuploidy, which may involve more than one chromosome. Fertilization involving a gamete that has no copy of a particular chromosome will lead to a missing chromosome in the zygote (so that the cell has 2n-1 chromosomes). The aneuploid zygote is said to be monosomic for that chromosome. If a chromosome is present in triplicate in the zygote (so that the cell has 2n+1), the aneuploid cell is trosomic for that chromosome. Mitosis will subsequently transmit the anomaly to all embryonic cells. If the organism survives, it usually has a set of traits caused by the abnormal does of the genes associated with the extra or missing chromosome. Down syndrome is an example of trisomy in human (on the 21st chromosome)- to be discussed more later. Nondisjunction can also occur during mitosis. If such an error takes place early in embryonic development, then the aneuploid condition is passed along by mitosis to a large number of cells and is likely to have a substantial effect. Some organisms have more than two complete chromosome sets in all somatic cells. The general term for this chromosomal alteration is polyploidy; the specific terms triploidy (3n) and tetraploidy (4n) indicated three or four chromosomal sets, respectively. One way a triploid cell may arise is by the fertilization of an abnormal diploid egg produced by nondisjunction of all its chromosomes. Tetraploidy could result from the failure of a 2n zygote to divide after replicating its chromosomes. Subsequent normal mitotic division would then produce a 4n embryo. Polyploidy is very common in plants, in fact many of the plants we eat are polyploid. Bananas are triploid and wheat is hexaploid (6n). In the animal kingdom, ployploid species are much less common, although they do occur in fish and amphibians. In general, polyploids are more nearly normal in appearance than aneuploids. One extra (or mission) chromosome apparently disrupts genetic balance more than does an entire extra set of chromosomes. *Each of these are a result of an accident in cell division (meiosis)
 or cause a variety of developmental disorders. Ideally, the meiotic spindle distributes chromosomes to daughter cells without error. But there is an occasional mishap, called a nondisjunction. (The other gametes are usually distributed normally). See next slide for diagram. If either of the aberrant gametes unites with a normal one at fertilization, the zygote will also have an abnormal number of a chromosome, a condition known as aneuploidy, which may involve more than one chromosome. Fertilization involving a gamete that has no copy of a particular chromosome will lead to a missing chromosome in the zygote (so that the cell has 2n-1 chromosomes). The aneuploid zygote is said to be monosomic for that chromosome. If a chromosome is present in triplicate in the zygote (so that the cell has 2n+1), the aneuploid cell is trosomic for that chromosome. Mitosis will subsequently transmit the anomaly to all embryonic cells. If the organism survives, it usually has a set of traits caused by the abnormal does of the genes associated with the extra or missing chromosome. Down syndrome is an example of trisomy in human (on the 21st chromosome)- to be discussed more later. Nondisjunction can also occur during mitosis. If such an error takes place early in embryonic development, then the aneuploid condition is passed along by mitosis to a large number of cells and is likely to have a substantial effect. Some organisms have more than two complete chromosome sets in all somatic cells. The general term for this chromosomal alteration is polyploidy; the specific terms triploidy (3n) and tetraploidy (4n) indicated three or four chromosomal sets, respectively. One way a triploid cell may arise is by the fertilization of an abnormal diploid egg produced by nondisjunction of all its chromosomes. Tetraploidy could result from the failure of a 2n zygote to divide after replicating its chromosomes. Subsequent normal mitotic division would then produce a 4n embryo. Polyploidy is very common in plants, in fact many of the plants we eat are polyploid. Bananas are triploid and wheat is hexaploid (6n). In the animal kingdom, ployploid species are much less common, although they do occur in fish and amphibians. In general, polyploids are more nearly normal in appearance than aneuploids. One extra (or mission) chromosome apparently disrupts genetic balance more than does an entire extra set of chromosomes. *Each of these are a result of an accident in cell division (meiosis)")
51
Trisomy 21

52
Triploidy (3n) is three sets of chromosomes
3. Polyploidy Polyploidy is a condition in which an organism has more than two complete sets of chromosomes a chromosomal alteration in which the organism possesses more than two complete chromosome sets. Triploidy (3n) is three sets of chromosomes Tetraploidy (4n) is four sets of chromosomes Polyploidy is common in plants, but not animals Polyploids are more normal in appearance than aneuploids Large-scale chromosomal alterations often lead to spontaneous abortions (miscarriages) or cause a variety of developmental disorders Ideally, the meiotic spindle distributes chromosomes to daughter cells without error. But there is an occasional mishap, called a nondisjunction. (The other gametes are usually distributed normally). See next slide for diagram If either of the aberrant gametes unites with a normal one at fertilization, the zygote will also have an abnormal number of a chromosome, a condition known as aneuploidy, which may involve more than one chromosome. Fertilization involving a gamete that has no copy of a particular chromosome will lead to a missing chromosome in the zygote (so that the cell has 2n-1 chromosomes). The aneuploid zygote is said to be monosomic for that chromosome. If a chromosome is present in triplicate in the zygote (so that the cell has 2n+1), the aneuploid cell is trosomic for that chromosome. Mitosis will subsequently transmit the anomaly to all embryonic cells. If the organism survives, it usually has a set of traits caused by the abnormal does of the genes associated with the extra or missing chromosome. Down syndrome is an example of trisomy in human (on the 21st chromosome)- to be discussed more later. Nondisjunction can also occur during mitosis. If such an error takes place early in embryonic development, then the aneuploid condition is passed along by mitosis to a large number of cells and is likely to have a substantial effect. Some organisms have more than two complete chromosome sets in all somatic cells. The general term for this chromosomal alteration is polyploidy; the specific terms triploidy (3n) and tetraploidy (4n) indicated three or four chromosomal sets, respectively. One way a triploid cell may arise is by the fertilization of an abnormal diploid egg produced by nondisjunction of all its chromosomes. Tetraploidy could result from the failure of a 2n zygote to divide after replicating its chromosomes. Subsequent normal mitotic division would then produce a 4n embryo. Polyploidy is very common in plants, in fact many of the plants we eat are polyploid. Bananas are triploid and wheat is hexaploid (6n). In the animal kingdom, ployploid species are much less common, although they do occur in fish and amphibians. In general, polyploids are more nearly normal in appearance than aneuploids. One extra (or mission) chromosome apparently disrupts genetic balance more than does an entire extra set of chromosomes. *Each of these are a result of an accident in cell division (meiosis)
 is three sets of chromosomes. Tetraploidy (4n) is four sets of chromosomes. Polyploidy is common in plants, but not animals. Polyploids are more normal in appearance than aneuploids. Large-scale chromosomal alterations often lead to spontaneous abortions (miscarriages) or cause a variety of developmental disorders. Ideally, the meiotic spindle distributes chromosomes to daughter cells without error. But there is an occasional mishap, called a nondisjunction. (The other gametes are usually distributed normally). See next slide for diagram. If either of the aberrant gametes unites with a normal one at fertilization, the zygote will also have an abnormal number of a chromosome, a condition known as aneuploidy, which may involve more than one chromosome. Fertilization involving a gamete that has no copy of a particular chromosome will lead to a missing chromosome in the zygote (so that the cell has 2n-1 chromosomes). The aneuploid zygote is said to be monosomic for that chromosome. If a chromosome is present in triplicate in the zygote (so that the cell has 2n+1), the aneuploid cell is trosomic for that chromosome. Mitosis will subsequently transmit the anomaly to all embryonic cells. If the organism survives, it usually has a set of traits caused by the abnormal does of the genes associated with the extra or missing chromosome. Down syndrome is an example of trisomy in human (on the 21st chromosome)- to be discussed more later. Nondisjunction can also occur during mitosis. If such an error takes place early in embryonic development, then the aneuploid condition is passed along by mitosis to a large number of cells and is likely to have a substantial effect. Some organisms have more than two complete chromosome sets in all somatic cells. The general term for this chromosomal alteration is polyploidy; the specific terms triploidy (3n) and tetraploidy (4n) indicated three or four chromosomal sets, respectively. One way a triploid cell may arise is by the fertilization of an abnormal diploid egg produced by nondisjunction of all its chromosomes. Tetraploidy could result from the failure of a 2n zygote to divide after replicating its chromosomes. Subsequent normal mitotic division would then produce a 4n embryo. Polyploidy is very common in plants, in fact many of the plants we eat are polyploid. Bananas are triploid and wheat is hexaploid (6n). In the animal kingdom, ployploid species are much less common, although they do occur in fish and amphibians. In general, polyploids are more nearly normal in appearance than aneuploids. One extra (or mission) chromosome apparently disrupts genetic balance more than does an entire extra set of chromosomes. *Each of these are a result of an accident in cell division (meiosis)")
53
B. Alterations of Chromosomal Structure
1. Breakage of a chromosome can lead to four types of changes in chromosome structure: a. Deletion removes a chromosomal segment The affected chromosome is then missing certain genes. (If the centromere is deleted, the entire chromosome will be lost). The deleted fragment may become attached as an extra segment to a sister chromatid, producing a duplication. Alternatively, a detached fragment could attach to a nonsister chromatid of a homologous chromsome. In that case, thought the duplicated segments might not be identical because the homologs could carry different alleles of certain genes. =A chromosomal fragment may also reattach to the original chromosome but in the reverse orientation, producing an inversion. A fourth possible result of chromosomal breakage is for the fragment to join a nonhomologous chromosome, a rearrangement called a translocation. Breakage of chromosomes can result in four types of structural changes (listed on ppt) A diploid embryo that is homozygous for a large deletion (or has a single X chromosome with a large deletion, in male) is usually missing a number of essential genes, a condition that is ordinarily lethal. Duplications and translocations also tend to be harmful. In reciprocal translocations, in which segment are exchanged between nonhomologous chromosomes, and inversions, the balance of genes is not abnormal- all genes are present in their normal doses. Nevertheless, translocations and inversions can alter phenotype because a gene’s expression can be influenced by its location among neighboring genes; such events sometimes have devastating effects. Deletions and duplications are especially likely to occur during meiosis. In crossing over, nonsister chromatids sometimes exchange unequal- sized segments of DNA, so that one partner gives up more genes than it receives. The products of such a non-reciprocal crossover are one chromosome with a deletion and one chromosome with a duplication.
. The deleted. fragment may become attached as an extra segment to a sister chromatid, producing a duplication. Alternatively, a detached fragment could. attach to a nonsister chromatid of a homologous chromsome. In that case, thought the duplicated segments might not be identical because. the homologs could carry different alleles of certain genes. =A chromosomal fragment may also reattach to the original chromosome but in. the reverse orientation, producing an inversion. A fourth possible result of chromosomal breakage is for the fragment to join a. nonhomologous chromosome, a rearrangement called a translocation. Breakage of chromosomes can result in four types of structural changes (listed on ppt) A diploid embryo that is homozygous for a large deletion (or has a single X chromosome with a large deletion, in male) is usually missing a number of essential genes, a condition that is ordinarily lethal. Duplications and translocations also tend to be harmful. In reciprocal translocations, in which segment are exchanged between nonhomologous chromosomes, and inversions, the balance of genes is not abnormal- all genes are present in their normal doses. Nevertheless, translocations and inversions can alter phenotype because a gene’s expression can be influenced by its location among neighboring genes; such events sometimes have devastating effects. Deletions and duplications are especially likely to occur during meiosis. In crossing over, nonsister chromatids sometimes exchange unequal- sized segments of DNA, so that one partner gives up more genes than it receives. The products of such a non-reciprocal crossover are one. chromosome with a deletion and one chromosome with a duplication.")
54
b. Duplication repeats a segment

55
c. An inversion occurs if the fragment reattaches to the original chromosome in reverse order.
Inversion reverses a segment within a chromosome

56
d. A translocation occurs if a chromosomal fragment joins to a nonhomologous chromosome

57
2. Crossing over can produce deletions or duplicatons.

58
C. Chromosomal Alterations in Human Disease 1
C. Chromosomal Alterations in Human Disease 1. Alterations of chromosome number and structure are associated with some serious disorders 2. Some types of aneuploidy appear to upset the genetic balance less than others, resulting in individuals surviving to birth and beyond 3. These surviving individuals have a set of symptoms, or syndrome, characteristic of the type of aneuploidy

59
4. Examples of Autosomal Aneuploidy:
a. Down’s Syndrome Trisomy 21 Related to age of parent Most common birth defect in U. S. (1/700 births) b. Patau Syndrome Trisomy 13 c. Edward’s Syndrome Trisomy 18
 b. Patau Syndrome. Trisomy 13. c. Edward’s Syndrome. Trisomy 18.")
60
5. Examples of Sex Chromosome Aneuploidy: (less severe)
a. Klinefelter’s Syndrome--Genotype usually XXY b. Extra Y--XYY c. Trisomy X (metafemales)--XXX d. Turner’s Syndrome (monosomy X)--XO 6. Examples of Deletions: a. Cri du chat Syndrome--Deletion on chromosome 5 7. Examples of Translocations: a. Chronic Myelogenous Leukemia (CML)--Portion of chromosome 22 switched with a fragment of chromosome 9 b. Type of Down’s Syndrome--Translocation from chromosome 21 to chromosome 15
--XXX. d. Turner’s Syndrome (monosomy X)--XO. 6. Examples of Deletions: a. Cri du chat Syndrome--Deletion on chromosome Examples of Translocations: a. Chronic Myelogenous Leukemia (CML)--Portion of chromosome 22 switched with a fragment of chromosome 9. b. Type of Down’s Syndrome--Translocation from chromosome 21 to chromosome 15.")
61
V. Concept 15.5: Exceptions to Chromosome Theory
There are two normal exceptions to Mendelian genetics One exception involves genes located in the nucleus, and the other exception involves genes located outside the nucleus Note that this phenomenon is different from sex linkage; most imprinted genes are on autosomes. Genomic imprinting occurs during the formation of gametes and results in the silencing of one allele of certain genes. Because these genes are imprinted differently in sperm and eggs, a zygote expresses only one allele of an imprinted gene, either an allele inherited from the female parent or the allele inherited from the male parent. The imprints are transmitted to all the body cells during development, so either the maternal or paternal allele of a given imprinted gene is expressed in every cell of that organism. In each generation, the old imprints are “erased” in gamete-producing cells, and the chromosomes of the developing gametes are newly imprinted in the same way. For instance, a gene imprinted for a maternal allele expression is always imprinted for maternal allele expression, generation after generation. 2. Mitochondria, as well as cholorplasts and other plant plastids, contain small circular DNA molecules that carry a number of genes. These organelles reproduce themselves and transmit their genes to daughter organelles. Organelle genes are not distributed to offspring according to the same rules that direct the distribution of nuclear chromosomes during meiosis, so they do not display Mendelian inheritance. Extranuclear genes are inherited maternally because the zygote’s cytoplasm comes from the egg. The first evidence of extranuclear genes came from studies on the inheritance of yellow or white patches on leaves of an otherwise green plant.

62
V. Concept 15.5: Exceptions to Chromosome Theory
A. Genomic Imprinting For a few mammalian traits, the phenotype depends on which parent passed along the alleles for those traits Such variation in phenotype is called genomic imprinting Genomic imprinting involves the silencing of certain genes that are “stamped” with an imprint during gamete production “a variation in phenotype depending on whether an allele is inherited from the male or female parent. Note that this phenomenon is different from sex linkage; most imprinted genes are on autosomes. Genomic imprinting occurs during the formation of gametes and results in the silencing of one allele of certain genes. Because these genes are imprinted differently in sperm and eggs, a zygote expresses only one allele of an imprinted gene, either an allele inherited from the female parent or the allele inherited from the male parent. The imprints are transmitted to all the body cells during development, so either the maternal or paternal allele of a given imprinted gene is expressed in every cell of that organism. In each generation, the old imprints are “erased” in gamete-producing cells, and the chromosomes of the developing gametes are newly imprinted in the same way. For instance, a gene imprinted for a maternal allele expression is always imprinted for maternal allele expression, generation after generation. 2. Mitochondria, as well as cholorplasts and other plant plastids, contain small circular DNA molecules that carry a number of genes. These organelles reproduce themselves and transmit their genes to daughter organelles. Organelle genes are not distributed to offspring according to the same rules that direct the distribution of nuclear chromosomes during meiosis, so they do not display Mendelian inheritance. Extranuclear genes are inherited maternally because the zygote’s cytoplasm comes from the egg. The first evidence of extranuclear genes came from studies on the inheritance of yellow or white patches on leaves of an otherwise green plant.

63
4. It appears that imprinting is the result of the methylation
4. It appears that imprinting is the result of the methylation (addition of –CH3) of DNA 5. Genomic imprinting is thought to affect only a small fraction of mammalian genes 6. Most imprinted genes are critical for embryonic development
 of DNA. 5. Genomic imprinting is thought to affect only a small fraction of mammalian genes. 6. Most imprinted genes are critical for embryonic development.")
64
B. Inheritance of Organelle Genes
Extranuclear genes (or cytoplasmic genes) are genes found in organelles in the cytoplasm Mitochondria, chloroplasts, and other plant plastids carry small circular DNA molecules Extranuclear genes are inherited maternally because the zygote’s cytoplasm comes from the egg The first evidence of extranuclear genes came from studies on the inheritance of yellow or white patches on leaves of an otherwise green plant 5. Some defects in mitochondrial genes prevent cells from making enough ATP and result in diseases that affect the muscular and nervous systems 6. Cytoplasmic genes described in plants by Karl Correns (1909)
 are genes found in organelles in the cytoplasm. Mitochondria, chloroplasts, and other plant plastids carry small circular DNA molecules. Extranuclear genes are inherited maternally because the zygote’s cytoplasm comes from the egg. The first evidence of extranuclear genes came from studies on the inheritance of yellow or white patches on leaves of an otherwise green plant. 5. Some defects in mitochondrial genes prevent cells from making enough ATP and result in diseases that affect the muscular and nervous systems. 6. Cytoplasmic genes described in plants by Karl Correns (1909)")
65
You should now be able to:
Explain the chromosomal theory of inheritance and its discovery Explain why sex-linked diseases are more common in human males than females Distinguish between sex-linked genes and linked genes Explain how meiosis accounts for recombinant phenotypes Explain how linkage maps are constructed

66
Explain how nondisjunction can lead to aneuploidy
Define trisomy, triploidy, and polyploidy Distinguish among deletions, duplications, inversions, and translocations Explain genomic imprinting Explain why extranuclear genes are not inherited in a Mendelian fashion

Similar presentations