Download presentation
Presentation is loading. Please wait.

1
Dawson’s beetle work shows that deleterious rare alleles may be very hard to eliminate from a gene pool because they remain hidden from selection as heterozygotes.

2
This only applies if the allele is not dominant. A dominant allele is expressed both as a heterozygote and a homozygote and so is always visible to selection.

3
One way in which multiple alleles may be maintained in a population is through heterozygote advantage (also called overdominance). Classic example is sickle cell allele.

4
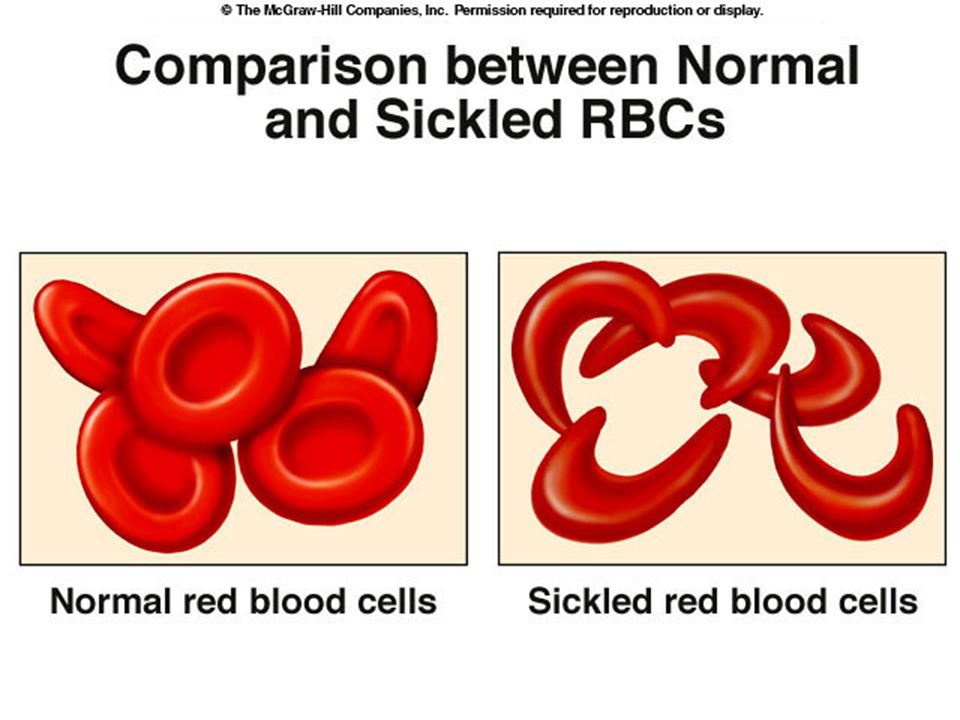
Sickle cell anemia is a condition common among West Africans and those of West African descent. Under low oxygen conditions the red blood corpuscles are sickle shaped. Untreated the condition usually causes death in childhood.

6
About 1% of West Africans have sickle cell anemia. A single mutation causes a valine amino acid to replace a glutamine in the alpha chain of hemoglobin The mutation causes hemoglobin molecules to stick together.

8
Only individuals homozygous for the allele get sickle cell anemia. Individuals with only one copy of the allele (heterozygotes) get sickle cell trait (a mild form of the disease) Individuals with the sickle cell allele (one or two copies) don’t get malaria.
 get sickle cell trait (a mild form of the disease) Individuals with the sickle cell allele (one or two copies) don’t get malaria..")
11
Heterozygotes have higher survival than either homozygote (heterozygote advantage). Sickle cell homozygotes die of sickle cell anemia, many “normal” homozygotes die of malaria. Stabilizing selection thus favors sickle cell allele.

12
A heterozygote advantage (or overdominance) results in a balanced polymorphism in a population. Both alleles are maintained in the population as the heterozygote is the best combination of alleles and a purely heterozygous population is not possible.

14
Underdominance is when the heterozygote has lower fitness than either homozygote. This situation is In this case one or other allele will go to fixation, but which depends on the starting allele frequencies

16
In some cases the costs and benefits of a trait depend on how common it is in a population.

17
In this case the commoner a phenotype is the more successful it is. If two phenotypes are determined by single alleles one allele will go to fixation and the other be lost, but which one depends on the starting frequencies.

19
In “flat” snails individuals mate face to face and physical constraints mean only individuals whose shells coil in the same direction can mate successfully. Higher frequencies of one coil direction leads to more mating for that phenotype and eventually it replaces the other types.

21
Under negative frequency-dependent selection a trait is increasingly favored the rarer it becomes.

23
Color polymorphism in Elderflower Orchid Two flower colors: yellow and purple. Offer no food reward to bees. Bees alternate visits to colors. How are two colors maintained in the population?

24
Gigord et al. hypothesis: Bees tend to visit equal numbers of each flower color so rarer color will have advantage (will get more visits from pollinators).
..")
25
Experiment: provided five arrays of potted orchids with different frequencies of yellow orchids in each. Monitored orchids for fruit set and removal of pollinaria (pollen bearing structures)
.")
26
As predicted, reproductive success of yellow varied with frequency.

27
5.21 a

28
Most mutations are deleterious and natural selection acts to remove them from population. Deleterious alleles persist, however, because mutation continually produces them.

29
When rate at which deleterious alleles being eliminated is equal to their rate of production by mutation we have mutation- selection balance.

30
Equilibrium frequency of deleterious allele q = square root of µ/s where µ is mutation rate and s is the selection coefficient (measure of strength of selection against allele; ranges from 0 to 1). See Box 6.6 for derivation of equation.

31
Equation makes intuitive sense. If s is small (mutation only mildly deleterious) and µ (mutation rate) is high than q (allele frequency) will also be relatively high. If s is large and µ is low, than q will be low too.
 and µ (mutation rate) is high than q (allele frequency) will also be relatively high. If s is large and µ is low, than q will be low too..")
32
Spinal muscular atrophy is a generally lethal condition caused by a mutation on chromosome 5. Selection coefficient estimated at 0.9. Deleterious allele frequency about 0.01 in Caucasians. Inserting above numbers into equation and solving for µ get estimated mutation rate of 0.9 X 10 -4

33
Observed mutation rate is about 1.1 X10 -4, very close agreement in estimates. High frequency of allele accounted for by observed mutation rate.

34
Cystic fibrosis is caused by a loss of function mutation at locus on chromosome 7 that codes for CFTR protein (cell surface protein in lungs and intestines). Major function of protein is to destroy Pseudomonas aeruginosa bacteria. Bacterium causes severe lung infections in CF patients.

35
Very strong selection against CF alleles, but CF frequency about 0.02 in Europeans. Can mutation rate account for high frequency?

36
Assume selection coefficient (s) of 1 and q = 0.02. Estimate mutation rate µ is 4.0 X 10 -4 But actual mutation rate is only 6.7 X 10 -7

37
Is there an alternative explanation?

38
May be heterozygote advantage. Pier et al. (1998) hypothesized CF heterozygotes may be resistant to typhoid fever. Typhoid fever caused by Salmonella typhi bacteria. Bacteria infiltrate gut by crossing epithelial cells.
 hypothesized CF heterozygotes may be resistant to typhoid fever. Typhoid fever caused by Salmonella typhi bacteria. Bacteria infiltrate gut by crossing epithelial cells..")
39
Hypothesized that S. typhi bacteria may use CFTR protein to enter cells. If so, CF-heterozygotes should be less vulnerable to S. typhi because their gut epithilial cells have fewer CFTR proteins on cell surface.

40
Experimental test. Produced mouse cells with three different CFTR genotypes CFTR homozygote (wild type) CFTR/ F508 heterozygote ( F508 most common CF mutant allele) F508/ F508 homozygote
 CFTR/ F508 heterozygote ( F508 most common CF mutant allele) F508/ F508 homozygote.")
41
Exposed cells to S. typhi bacteria. Measured number of bacteria that entered cells. Clear results

42
Fig 5.27a

43
F508/ F508 homozygote almost totally resistant to S. typhi. Wild type homozygote highly vulnerable Heterozygote contained 86% fewer bacteria than wild type.

44
Further support for idea F508 provides resistance to typhoid provided by positive relationship between F508 allele frequency in generation after typhoid outbreak and severity of the outbreak.

45
Fig 5.27b Data from 11 European countries

46
Another assumption of Hardy-Weinberg is that random mating takes place. The most common form of non-random mating is inbreeding which occurs when close relatives mate with each other.

47
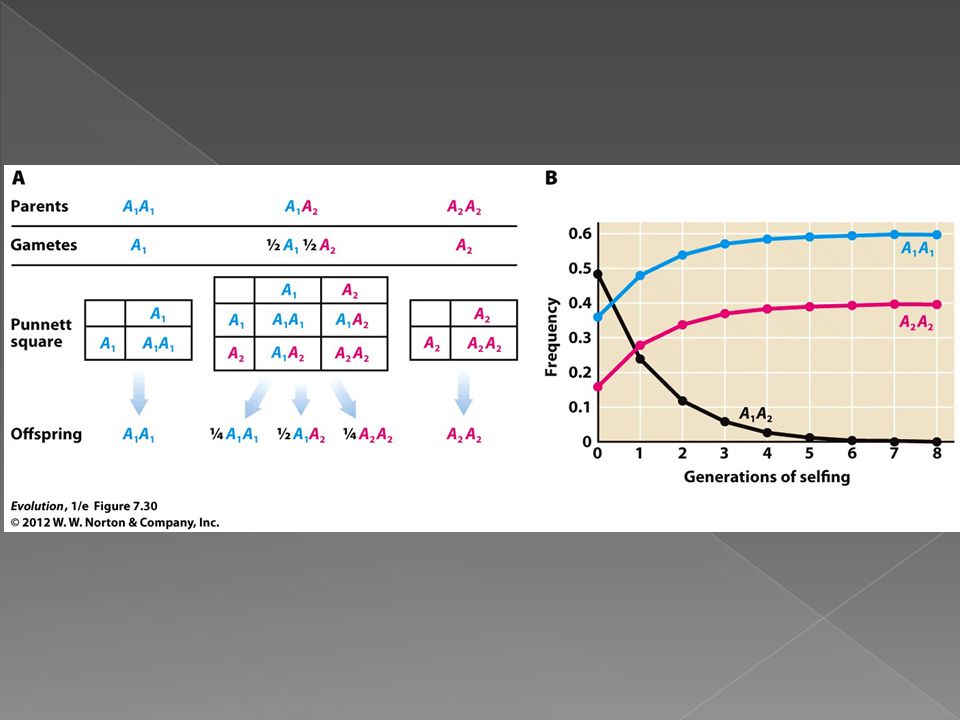
Most extreme form of inbreeding is self fertilization. In a population of self fertilizing organisms all homozygotes will produce only homozygous offspring. Heterozygotes will produce offspring 50% of which will be homozygous and 50% heterozygous. How will this affect the frequency of heterozygotes each generation?

48
In each generation the proportion of heterozygous individuals in the population will decline.

50
Because inbreeding produces an excess of homozygotes in a population, deviations from Hardy-Weinberg expectations can be used to detect such inbreeding in wild populations.

51
Sea otters once abundant along the west coast of the U.S were almost wiped out by fur hunters in the 18 th and 19 th centuries. California population reached a low of 50 individuals (now over 1,500). As a result of this bottleneck the population has less genetic diversity than it once had.
. As a result of this bottleneck the population has less genetic diversity than it once had..")
52
Population still at a low density and Lidicker and McCollum (1997) investigated whether this resulted in inbreeding. Determined genotypes of 33 otters for PAP locus, which has two alleles S (slow) and F (fast)
 and F (fast).")
53
The genotypes of the 33 otters were: › SS 16 › SF 7 › FF 10 This gives approximate allele frequencies of S= 0.6 and F = 0.4

54
If otter population in H-W equilibrium, genotype frequencies should be › SS = 0.6* 0.6 = 0.36 › SF =2*0.6*0.4 = 0.48 › FF = 0.4*0.4 = 0.16 However actual frequencies were: › SS= 0.485, SF= 0.212, FF =0.303

55
There are more homozygotes and fewer heterozygotes than expected for a random mating population. Having considered alternative explanations for deficit of heterozygotes Lidicker and McCollum (1997) concluded that sea otter populations show evidence of inbreedng.
 concluded that sea otter populations show evidence of inbreedng..")
56
Self-fertilization and sibling mating most extreme forms of inbreeding, but matings between more distant relatives (e.g. cousins) has same effect on frequency of homozygotes, but rate is slower.
 has same effect on frequency of homozygotes, but rate is slower..")
57
F = Coefficient of inbreeding: probability that two alleles in an individual are identical by descent (both alleles are copies of a particular ancestor’s allele in some previous generation). F increases as relatedness increases.

59
If we compare heterozygosity of inbred population H f with that of a random mating population H o relationship is H f = H o (1-F) Anytime F>0 frequency of heterozygotes is reduced and frequency of homozygotes naturally increases.
 Anytime F>0 frequency of heterozygotes is reduced and frequency of homozygotes naturally increases.")
60
Calculating F. Need to use pedigree diagrams. Example: Female is daughter of two half- siblings. Two ways female could receive alleles that are identical by descent.

61
Fig 6.27a MaleFemaleMale Female Male Half-sibling mating

62
Fig 6.27b

63
Total probability of scenario is 1/16 + 1/16 = 1/8.

64
Inbreeding increases frequency of homozygotes and thus the probability that deleterious alleles are visible to selection. In humans, children of first cousins have higher mortality rates than children of unrelated individuals.

65
Fig 6.28 Each dot on graph represents mortality rates for a human population. Mortality rate for children of cousins consistently about 4% higher than rate for children of non-relatives.

66
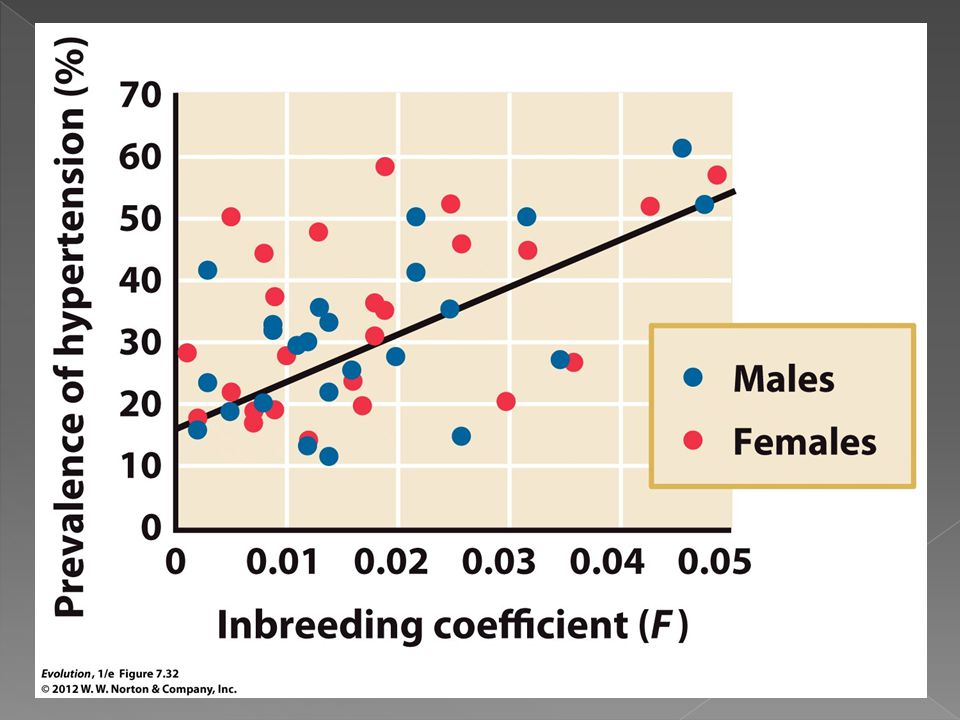
In a study of 2760 individuals from 25 Croatian islands Rudan et al. found a strong positive relationship between high blood pressure and the inbreeding coefficent.

68
Royal families have been particularly prone to inbreeding. In Ancient Egypt because royal women were considered to carry the royal bloodline the pharaoh routinely was married to a sister or half-sister.

69
The most famous example of a genetic disorder exacerbated by inbreeding is the Hapsburg jaw or Hapsburg lip [severe lower jaw protrusion]. (Hapsburgs were the ruling family of Austria and Spain for much of the 1400’s- 1700’s)
![ The most famous example of a genetic disorder exacerbated by inbreeding is the Hapsburg jaw or Hapsburg lip [severe lower jaw protrusion].](http://images.slideplayer.com/13/3882249/slides/slide_69.jpg " (Hapsburgs were the ruling family of Austria and Spain for much of the 1400’s- 1700’s).")
71
The last of the Spanish Hapsburgs, Charles II (1661- 1700) had such severe jaw protrusion he could not chew his food properly. Charles II also had a large number of other recessively inherited genetic problems that caused physical, mental, sexual and other problems. Charles was infertile and the last of the Spanish Hapsburg kings.

72
http://en.wikipedia.org/wiki/Charles_II_of_Spain

Similar presentations

















 2.Male and female parts of the plant.>")





>")






