Download presentation
Presentation is loading. Please wait.

1
Population Genetics: Selection and mutation as mechanisms of evolution
Population genetics: study of Mendelian genetics at the level of the whole population.

2
Hardy-Weinberg Equilibrium
To understand the conditions under which evolution can occur, it is necessary to understand the population genetic conditions under which it will not occur.

3
Hardy-Weinberg Equilibrium
The Hardy-Weinberg Equilibrium Principle allows us do this. It enables us to predict allele and genotype frequencies from one generation to the next in the absence of evolution.

4
Hardy-Weinberg Equilibrium
Assume a gene with two alleles A and a with known frequencies (e.g. A = 0.6, a = 0.4.) There are only two alleles in the population so their frequencies must add up to 1.
 There are only two alleles in the population so their frequencies must add up to 1.")
5
Hardy-Weinberg Equilibrium
Using allele frequencies we can predict the expected frequencies of genotypes in the next generation. With two alleles only three genotypes are possible: AA, Aa and aa

6
Hardy-Weinberg Equilibrium
Assume alleles A and a are present in eggs and sperm in proportion to their frequency in population (i.e. 0.6 and 0.4) Also assume that sperm and eggs meet at random (one big gene pool).
 Also assume that sperm and eggs meet at random (one big gene pool).")
7
Hardy-Weinberg Equilibrium
Then we can calculate expected genotype frequencies. AA: To produce an AA individual, egg and sperm must each contain an A allele. This probability is 0.6 x 0.6 = 0.36 (probability sperm contains A times probability egg contains A).
.")
8
Hardy-Weinberg Equilibrium
Similarly, we can calculate frequency of aa. 0.4 x 04 = 0.16.

9
Hardy-Weinberg Equilibrium
Probability of Aa is given by probability sperm contains A (0.6) times probability egg contains a (0.4) x 04 = 0.24.
 times probability egg contains a (0.4). 0.6 x 04 =")
10
Hardy-Weinberg Equilibrium
But, there’s a second way to produce an Aa individual (egg contains A and sperm contains a). Same probability as before: 0.6 x 0.4= 0.24. Hence the overall probability of Aa = = 0.48.
. Same probability as before: 0.6 x 0.4= Hence the overall probability of Aa = =")
11
Hardy-Weinberg Equilibrium
Genotypes in next generation: AA = 0.36 Aa = 0.48 Aa= 0.16 These frequencies add up to one.

12
Hardy-Weinberg Equilibrium
General formula for Hardy-Weinberg. Let p= frequency of allele A and q = frequency of allele a. p2 + 2pq + q2 = 1.

13
Hardy Weinberg Equilibrium with more than 2 alleles
If three alleles with frequencies P1, P2 and P3 such that P1 + P2 + P3 = 1 Then genotype frequencies given by: P12 + P22 + P32 + 2P1P2 + 2P1 P3 + 2P2P3

14
Conclusions from Hardy-Weinberg Equilibrium
Allele frequencies in a population will not change from one generation to the next just as a result of assortment of alleles and zygote formation. Assortment of alleles simply means what occurs during meiosis when only one copy of each pair of alleles enters any given gamete (remember each gamete only contains half the DNA of a body cell).
.")
15
Conclusions from Hardy-Weinberg Equilibrium
If the allele frequencies in a gene pool with two alleles are given by p and q, the genotype frequencies will be given by p2, 2pq, and q2.

16
Working with the H-W equation
You need to be able to work with the Hardy-Weinberg equation. For example, if 9 of 100 individuals in a population suffer from a homozygous recessive disorder can you calculate the frequency of the disease-causing allele? Can you calculate how many heterozygotes are in the population?

17
Working with the H-W equation
p2 + 2pq + q2 = 1. The terms in the equation represent the frequencies of individual genotypes. [A genotype is possessed by an individual organism so there are two alleles present in each case.] P and q are allele frequencies. Allele frequencies are estimates of how common alleles are in the whole population. It is vital that you understand the difference between allele and genotye frequencies.

18
Working with the H-W equation
9 of 100 (frequency = 0.09) of individuals are homozygous for the recessive allele. What term in the H-W equation is that equal to?
 of individuals are homozygous for the recessive allele. What term in the H-W equation is that equal to")
19
Working with the H-W equation
It’s q2. If q2 = 0.09, what’s q? Get square root of q2, which is 0.3, which is the frequency of the allele a. If q=0.3 then p=0.7. Now plug p and q into equation to calculate frequencies of other genotypes.

20
Working with the H-W equation
p2 = (0.7)(0.7) = frequency of AA 2pq = 2 (0.3)(0.7) = 0.42 – frequency of Aa. To calculate the actual number of heterozygotes simply multiply 0.42 by the population size = (0.42)(100) = 42.
(0.7) = frequency of AA. 2pq = 2 (0.3)(0.7) = 0.42 – frequency of Aa. To calculate the actual number of heterozygotes simply multiply 0.42 by the population size = (0.42)(100) = 42.")
21
Working with the H-W equation: 3 alleles
There are three alleles in a population A1, A2 and A3 whose frequencies respectively are 0.2, 0.2 and 0.6 and there are 100 individuals in the population. How many A1A2 heterozygotes will there be in the population?

22
Working with the H-W equation: 3 alleles
Just use the formulae P1 + P2 + P3 = 1 and P12 + P22 + P32 + 2P1P2 + 2P1 P3 + 2P2P3 = 1 Then substitute in the appropriate values for the appropriate term 2P1P2 = 2(0.2)(0.2) = 0.08 or 8 people out of 100.
(0.2) = 0.08 or 8 people out of 100.")
23
Other examples of working with HW equilibrium: is a population in HW equilibrium?
In a population there are 100 birds with the following genotypes: 44 AA 32 Aa 24 aa How would you demonstrate that this population is not in Hardy Weinberg equilibrium

24
Three steps Step 1: calculate the allele frequencies.
Step 2: Calculate expected numbers of each geneotype (i.e. figure out how many homozygotes and heterozygotes you would expect.) Step 3: compare your expected and observed data.
 Step 3: compare your expected and observed data.")
25
Step 1 allele frequencies
Step 1. How many “A” alleles are there in total? 44 AA individuals = 88 “A” alleles (because each individual has two copies of the “A” allele) 32 Aa alleles = 32 “A” alleles Total “A” alleles is =120.
 32 Aa alleles = 32 A alleles. Total A alleles is =120.")
26
Step 1 allele frequencies
Total number of “a” alleles is similarly calculated as 2* = 80 What are allele frequencies? Total number of alleles in population is = 200 (or you could calculate it by multiplying the number of individuals in the population by two 100*2 =200)
")
27
Step 1 allele frequencies
Allele frequencies are: A = 120/200= 0.6. Let p = 0.6 a = 80/200 = 0.4. Let q = 0.4

28
Step 2 Calculate expected number of each genotype
Use the Hardy_Weinberg equation p2 + 2pq + q2 = 1 to calculate what expected genotypes we should have given these observed frequencies of “A” and “a” Expected frequency of AA = p2 = 0.6 * 0.6 = 0.36 Expected frequency of aa = q2 = 0.4*0 .4 =0.16 Expected frequency of Aa = 2pq = 2*.6*.4 = 0.48

29
Step 2 Calculate expected number of each genotype
Convert genotype frequencies to actual numbers by multiplying by population size of 100 AA = 0.36*100 = 36 aa = 0.16*100 = 16 Aa = 0.48*100 = 48

30
Step 3 Compare Observed and Expected values
Observed population is: 44 AA 32 Aa 24 aa Expected population is: 36AA 48Aa 16aa These numbers are not the same so the population is not in Hardy-Weinberg equilibrium. An assumption of the Hardy Weinberg equilibrium is being violated. What are those assumptions?

31
Assumptions of Hardy-Weinberg
1. No selection. When individuals with certain genotypes survive better than others, allele frequencies may change from one generation to the next.

32
Assumptions of Hardy-Weinberg
2. No mutation If new alleles are produced by mutation or alleles mutate at different rates, allele frequencies may change from one generation to the next.

33
Assumptions of Hardy-Weinberg
3. No migration Movement of individuals in or out of a population will alter allele and genotype frequencies.

34
Assumptions of Hardy-Weinberg
4. No chance events. Luck plays no role. Eggs and sperm collide at same frequencies as the actual frequencies of p and q. When assumption is violated, and by chance some individuals contribute more alleles than others to next generation, allele frequencies may change. This mechanism of allele frequency change is called Genetic Drift.

35
Assumptions of Hardy-Weinberg
5. Individuals select mates at random. If this assumption is violated allele frequencies will not change, but genotype frequencies may.

36
Hardy-Weinberg Equilibrium
Hardy Weinberg equilibrium principle identifies the forces that can cause evolution. If a population is not in H-W equilibrium then one or more of the five assumptions is being violated.

37
5.10

38
Can selection change allele frequencies?
Two alleles B1 and B2 Frequency of B1= 0.6 and frequency of B2 = 0.4. Random mating gives genotype frequencies B1B B1B B2B2

39
Can selection change allele frequencies?
Assume 100 individuals 36 B1B B1B B2B2 Incorporate selection. Assume all B1B1 survive, 75% of B1B2 survive and 50% of B2B2 survive.

40
Can selection change allele frequencies?
Population now 80 individuals: 36 B1B B1B2 8 B2B2 Allele frequencies now: B1 = /160 = 0.675 B2 = 36+16/160 = 0.325 Selection resulted in allele frequency change.

41
FIG 5.11

42
Can selection change allele frequencies?
Selection in previous example very strong. What patterns expected with weaker selection. Initial frequencies B1 = 0.01, B2 = 0.99.

43
Fig 5.12 Note line colors yellow and red have been flipped in the table.

44
Can selection change allele frequencies?
Rate of change of B1is rapid when selection pressure is strong, but much slower, although still steady, under weak selection.

45
Empirical examples of allele frequency change under selection
Clavener and Clegg’s work on Drosophila. Two alleles for ADH (alcohol dehydrogenase breaks down ethanol) ADHF and ADHS
 ADHF and ADHS.")
46
Empirical examples of allele frequency change under selection
Two Drosophila populations maintained: one fed food spiked with ethanol, control fed unspiked food. Populations maintained for multiple generations.

47
Empirical examples of allele frequency change under selection
Experimental population showed consistent long-term increase in frequency of ADHF Flies with ADHF allele have higher fitness when ethanol is present. ADHF enzyme breaks down ethanol twice as fast as ADHS enzyme.

48
Fig 5.13

49
Empirical examples of allele frequency change under selection: Jaeken syndrome
Jaeken syndrome: patients severely disabled with skeletal deformities and inadequate liver function.

50
Jaeken syndrome Autosomal recessive condition caused by loss-of-function mutation of gene PMM2 codes for enzyme phosphomannomutase. Patients unable to join carbohydrates and proteins to make glycoproteins at a high enough rate. Glycoproteins involved in movement of substances across cell membranes.

51
Jaeken syndrome Many different loss-of-function mutations can cause Jaeken Syndrome. Team of researchers led by Jaak Jaeken investigated whether different mutations differed in their severity. Used Hardy-Weinberg equilibrium to do so.

52
Jaeken syndrome People with Jaeken syndrome are homozygous for the disease, but may be either homozygous or heterozygous for a given disease allele. Different disease alleles should be in Hardy-Weinberg equilibrium.

53
Jaeken syndrome Researchers studied 54 patients and identified most common mutation as R141H. Dividing population into R141H and other alleles [Other]. Allele frequencies are: Other: 0.6 and R141H: 0.4.

54
Jaeken syndrome If disease alleles are in H-W equilibrium then we predict genotype frequencies of Other/other: 0.36 Other/R141H: 0.48 R141H/R141H: 0.16

55
Jaeken syndrome Observed frequencies are: Other/Other: 0.2
Other/R141H: 0.8 R141H/R141H : 0 Clearly population not in H-W equilibrium.

56
Jaeken syndrome Researchers concluded that R141H is an especially severe mutation and homozygotes die before or just after birth. Thus, there is selection so H-W assumption is violated.

57
Using H-W to predict potential spread of CCR5-32 allele
Will AIDS epidemic cause CCR5-32 (delta32) allele to spread? Offers protection against HIV infection. In principle it could, but models suggest it probably will not in any real population.
 allele to spread Offers protection against HIV infection. In principle it could, but models suggest it probably will not in any real population.")
58
Spread of CCR5-32 allele Scenario 1. Initial allele frequency 20%. 25% of heterozygotes and those homozygous for normal allele die of AIDS. Homozygous 32 individuals do not die of AIDS. Over 40 generations allele increases to almost 100%.

59
FIG 5.15a

60
Spread of CCR5-32 allele In human population with high HIV infection rate and high frequencies of 32 allele, 32 could spread rapidly.

61
Spread of CCR5-32 allele Scenario 2. In areas with low HIV infection rates (under 1%), but high levels of 32 (as found in Europe), selection too weak to raise delta 32 frequency much.
, but high levels of 32 (as found in Europe), selection too weak to raise delta 32 frequency much.")
62
FIG 5.15b

63
Spread of CCR5-32 allele Scenario 3. In sub-Saharan Africa HIV infection rate about 25%. However, 32 allele almost absent Under these conditions 32 frequency will hardly change because most copies of allele are in heterozygotes, which are not protected from HIV.

64
FIG 5.15c

65
Testing predictions of population genetics theory
Theory predicts that if an individual carrying an allele has higher than average fitness then the frequency of that allele will increase from one generation to the next. Obviously, the converse should be true and a deleterious allele should decrease in frequency if its bearers have lower fitness.

66
Testing predictions of population genetics theory
Mathematical treatment of effect of selection on gene frequencies is given in Box 6.3 (page 186) of your text. Main point is that if the average fitness of an allele A when paired at random with other alleles in the population is higher than the average fitness of the population, then it will increase in frequency.
 of your text. Main point is that if the average fitness of an allele A when paired at random with other alleles in the population is higher than the average fitness of the population, then it will increase in frequency.")
67
Tests of theory Dawson (1970). Flour beetles. Two alleles at locus: + and l. +/+ and +/l phenotypically normal. l/l lethal.

68
Dawson’s flour beetles
Dawson founded two populations with heterozygotes (frequency of + and l alleles thus 0.5). Expected + allele to increase in frequency and l allele to decline over time.
. Expected + allele to increase in frequency and l allele to decline over time.")
69
Fig 5.16a

70
Dawson’s flour beetles
Predicted and observed allele frequencies matched very closely. l allele declined rapidly at first, but rate of decline slowed.

71
Dawson’s flour beetles
Dawson’s results show that when a recessive allele is common, evolution by natural selection is rapid, but slows as recessive allele becomes rarer. Hardy-Weinberg explains why.

72
Dawson’s flour beetles
When recessive allele (a) common e.g. freq a=0.95 genotype frequencies are: AA (0.05)2 Aa (2 (0.05)(0.95) aa (0.95)2 0.0025AA Aa aa With more than 90% of phenotypes being recessive, if aa is selected against we expect rapid population change.
 common e.g. freq a=0.95 genotype frequencies are: AA (0.05)2 Aa (2 (0.05)(0.95) aa (0.95) AA 0.095Aa aa. With more than 90% of phenotypes being recessive, if aa is selected against we expect rapid population change.")
73
Dawson’s flour beetles
When recessive allele (a) rare [e.g. freq a=0.05] genotype frequencies are: AA (0.95)2 Aa 2(0.95)(0.05) aa (0.05)2 0.9025AA Aa aa Fewer than 0.25% of phenotypes are aa recessive. Most “a” alleles (>97%) are hidden from selection as heterozygotes. Expect only slow change in frequency of a.
 rare [e.g. freq a=0.05] genotype frequencies are: AA (0.95)2 Aa 2(0.95)(0.05) aa (0.05) AA 0.095Aa aa. Fewer than 0.25% of phenotypes are aa recessive. Most a alleles (>97%) are hidden from selection as heterozygotes. Expect only slow change in frequency of a.")
74
Maintaining multiple alleles in gene pool
Dawson’s beetle work shows that deleterious rare alleles may be very hard to eliminate from a gene pool because they remain hidden from selection as heterozygotes. (this only applies if the allele is not dominant.)
")
75
Maintaining multiple alleles in gene pool
There are several different ways in which multiple alleles may be maintained in populations even, as we saw, if one allele is deleterious. 1. Deleterious recessive allele can hide in heterozygote 2. There may be heterozygote advantage 3. Frequency-dependent selection

76
Maintaining multiple alleles in gene pool-2. Heterozygote advantage
Another way in which multiple alleles may be maintained in a population is through heterozygote advantage. Classic example is sickle cell allele.

77
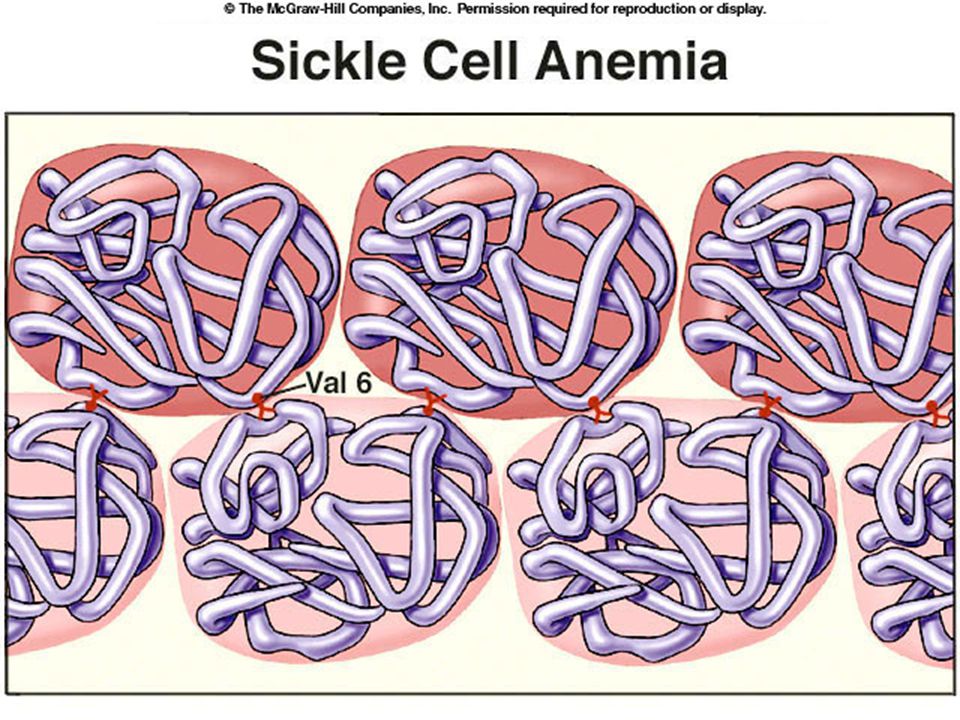
Sickle Cell Anemia Sickle cell anemia is a condition common among West Africans (and African Americans of West African ancestry). In sickle cell anemia red blood cells are sickle shaped. Usually fatal by about age 10.

79
About 1% of West Africans have sickle
cell anemia. A single mutation that causes a valine amino acid to replace a glutamine in an alpha chain of the hemoglobin molecule. Mutation causes molecules to stick together.

81
Why isn’t mutant sickle cell gene
eliminated by natural selection?

82
Only individuals homozygous for
sickle cell gene get sickle cell anemia. Individuals with one copy of sickle cell gene (heterozygotes) get sickle cell trait (mild form of disease). Individuals with sickle cell allele (one or two copies) don’t get malaria.
 get sickle cell trait. (mild form of disease). Individuals with sickle cell allele (one or two. copies) don’t get malaria.")
85
Heterozygotes have higher survival than
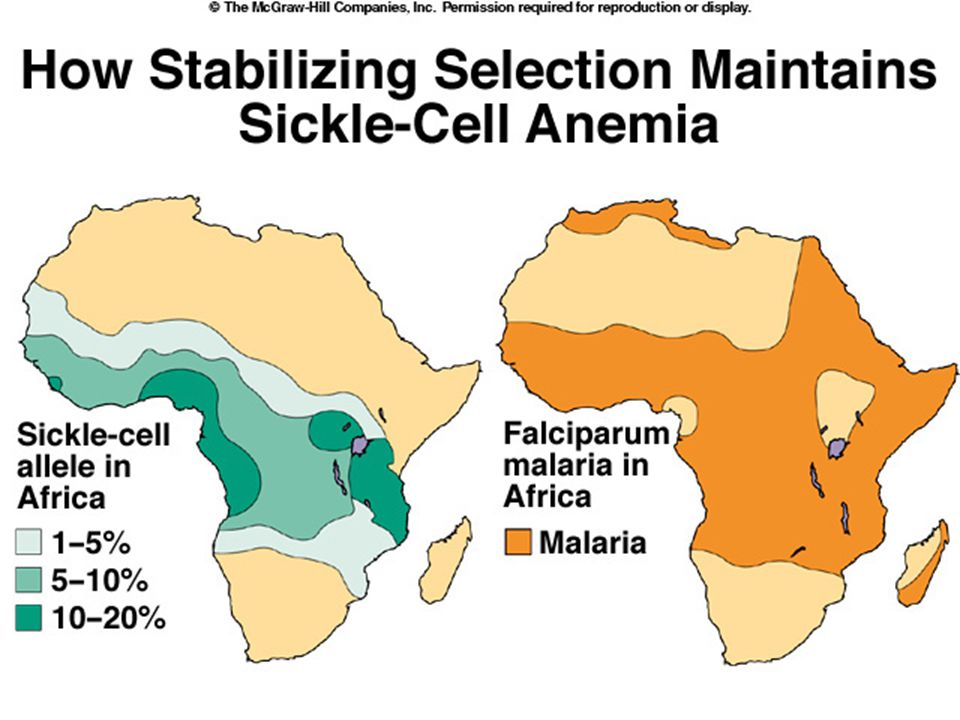
either homozygote. Heterozygote advantage. Sickle cell homozygotes die of sickle cell anemia. “Normal” homozygotes more likely to die of malaria. Stabilizing selection for sickle cell allele.

86
Maintaining multiple alleles in gene pool- 3
Maintaining multiple alleles in gene pool- 3. Frequency-dependent selection Another way in which multiple alleles are maintained is frequency-dependent selection. Frequency-dependent selection occurs when rare alleles have a selective advantage because they are rare.

87
Frequency-dependent selection
Color polymorphism in Elderflower Orchid Two flower colors: yellow and purple. Offer no food reward to bees. Bees alternate visits to colors. How are two colors maintained in the population?

88
Frequency-dependent selection
Gigord et al. hypothesis: Bees tend to visit equal numbers of each flower color. As a result the rarer color will have an advantage (because each individual rarer color flower will get more visits from pollinators).
.")
89
Frequency-dependent selection
Experiment: provided five arrays of potted orchids with different frequencies of yellow orchids in each. Monitored orchids for fruit set and removal of pollinaria (pollen bearing structures)
")
90
Frequency-dependent selection
As predicted, reproductive success of yellow varied with frequency.

91
5.21 a

92
Mutation as an evolutionary force
It is obvious that selection is a very powerful evolutionary force but how strong is mutation alone as an evolutionary force? To check: Two alleles A and a. Frequency of A = 0.9, a = 0.1.

93
Mutation as an evolutionary force
Assume A mutates to a at rate of 1 copy per 10,000 per generation (high rate, but within observed range) and all mutations occur in gametes. How much does this change gene pool in next generation?
 and all mutations occur in gametes. How much does this change gene pool in next generation")
94
Mutation as an evolutionary force
Hardy Weinberg genotypes in current generation: 0.81 AA, 0.18 Aa, 0.01 aa With no mutation allele frequency in gene pool 0.9 A, 0.1 a

95
Mutation as an evolutionary force
But mutation reduces frequency of A and increases frequency of a A a 0.9 - (0.0001)(0.9) (0.0001)(0.9) A a
(0.9) (0.0001)(0.9) A a.")
96
5.23

97
Mutation as an evolutionary force
Not a big change. After 1000 generations frequency of A = 0.81.

98
5.24

99
Mutation as an evolutionary force
Mutation alone clearly not a powerful evolutionary force. But mutation AND selection together make a very powerful evolutionary force. Remember mutation provides the raw material for selection to work with.

100
Lenski’s E. coli work Lenski et al. studied mutation and selection together in an E. coli strain that did not exchange DNA (hence mutation only source of new variation). Bacteria grown in challenging environment (low salts and low glucose medium) so selection would be strong.
. Bacteria grown in challenging environment (low salts and low glucose medium) so selection would be strong.")
101
Lenski’s E. coli work 12 replicate populations tracked over about 10,000 generations. Fitness and cell size of populations increased over time. Pattern of change interesting: steplike. Why is it steplike?

102
5.25

103
Lenski’s E. coli work Step-like pattern results when a new mutation occurs and sweeps through the population as mutant bacteria out-reproduce competitors. Remember, without mutation evolution would eventually cease. Mutation is the ultimate source of genetic variation.

104
Mutation-selection balance
Most mutations are deleterious and natural selection acts to remove them from population. Deleterious alleles persist, however, because mutation continually produces them.

105
Mutation-selection balance
When rate at which deleterious alleles are being eliminated is equal to their rate of production by mutation we have mutation-selection balance.

106
Mutation-selection balance
Equilibrium frequency of deleterious allele q = square root of µ/s where µ is mutation rate and s is the selection coefficient (measure of strength of selection against allele; ranges from 0 to 1). See Box 6.10 page 215 for derivation of equation.
. See Box 6.10 page 215 for derivation of equation.")
107
Mutation-selection balance
Equation makes intuitive sense. If s is small (mutation only mildly deleterious) and µ (mutation rate) is high than q (allele frequency) will also be relatively high. If s is large and µ is low, than q will be low too.
 and µ (mutation rate) is high than q (allele frequency) will also be relatively high. If s is large and µ is low, than q will be low too.")
108
Mutation-selection balance
Spinal muscular atrophy is a generally lethal condition caused by a mutation on chromosome 5. Selection coefficient estimated at Deleterious allele frequency about 0.01 in Caucasians. Inserting above numbers into equation and solving for µ get estimated mutation rate of 0.9 X 10-4

109
Mutation-selection balance
Observed mutation rate is about 1.1 X10-4, very close agreement in estimates. High frequency of allele is accounted for by observed mutation rate.

110
Is frequency of Cystic fibrosis maintained by mutation selection balance?
Cystic fibrosis is caused by a loss of function mutation at locus on chromosome 7 that codes for CFTR protein (cell surface protein in lungs and intestines). Major function of protein is to destroy Pseudomonas aeruginosa bacteria. Bacterium causes severe lung infections in CF patients.
. Major function of protein is to destroy Pseudomonas aeruginosa bacteria. Bacterium causes severe lung infections in CF patients.")
111
Cystic fibrosis Very strong selection against CF alleles, but CF frequency about 0.02 in Europeans. Can mutation rate account for high frequency?

112
Cystic fibrosis Assume selection coefficient (s) of 1 and q = 0.02.
Estimate mutation rate µ is 4.0 X 10-4 But actual mutation rate is only 6.7 X 10-7 Is there an alternative explanation?

113
Cystic fibrosis May be heterozygote advantage.
Pier et al. (1998) hypothesized CF heterozygotes may be resistant to typhoid fever. Typhoid fever caused by Salmonella typhi bacteria. Bacteria infiltrate gut by crossing epithelial cells.
 hypothesized CF heterozygotes may be resistant to typhoid fever. Typhoid fever caused by Salmonella typhi bacteria. Bacteria infiltrate gut by crossing epithelial cells.")
114
Cystic fibrosis Hypothesized that S. typhi bacteria may use CFTR protein to enter cells. If so, CF-heterozygotes should be less vulnerable to S. typhi because their gut epithilial cells have fewer CFTR proteins on cell surface.

115
Cystic fibrosis Experimental test.
Produced mouse cells with three different CFTR genotypes CFTR homozygote (wild type) CFTR/F508 heterozygote (F508 most common CF mutant allele) F508/F508 homozygote
 CFTR/F508 heterozygote (F508 most common CF mutant allele) F508/F508 homozygote.")
116
Cystic fibrosis Exposed cells to S. typhi bacteria.
Measured number of bacteria that entered cells. Clear results

117
Fig 5.27a

118
Cystic fibrosis F508/F508 homozygote almost totally resistant to S. typhi. Wild type homozygote highly vulnerable Heterozygote contained 86% fewer bacteria than wild type.

119
Cystic fibrosis Further support for idea F508 provides resistance to typhoid provided by positive relationship between F508 allele frequency in generation after typhoid outbreak and severity of the outbreak.

120
Fig 5.27b Data from 11 European countries

Similar presentations







 that mendel described.>")












