Download presentation
Presentation is loading. Please wait.

1
GROWTH FACTOR SIGNALING, HOX GENES AND THE BODY PLAN
MOLECULAR REGULATION OF DEVELOPMENT GROWTH FACTOR SIGNALING, HOX GENES AND THE BODY PLAN

2
Overview Pattern formation by morphogen gradients of growth factors and the dorsal-ventral (D-V) axis. B) Hox genes and the antero-posterior (A-P) axis. - colinearity - activation by retinoic acid - Hox genes in Evolution and Development
 Hox genes and the antero-posterior (A-P) axis. - colinearity. - activation by retinoic acid. - Hox genes in Evolution and Development.")
4
During development groups of inducing cells called organizing centers secrete graded growth factor signals. The concentration gradient of a morphogen can induce multiple cell differentiation choices.

5
D-V patterning can be best studied in the frog egg.

6
D-V cell differentiation depends of a gradient of BMP signaling.

7
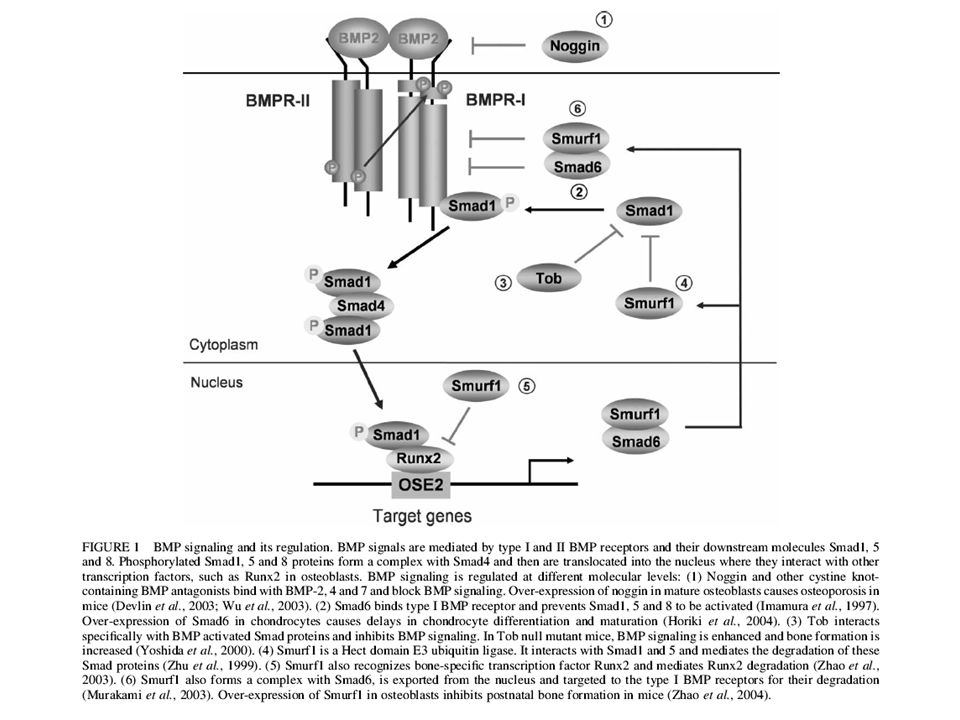
Secreted antagonists such as Chordin and Noggin can bind to growth factors in the extracellular space and prevent binding to cell surface receptors. This inhibitory mechanism is used in development to generate morphogen gradients. Chordin establishes a BMP4 gradient at gastrula. Chordin inhibits

8
Signal transduction: membrane receptors transduce the signal so that transcription factors are activated through phosphorylation. TGFβ family members (30 different ligands in humans) activate cell surface receptors (serine-threonine kinases). Xnr BMP4

9
Bone morphogenetic proteins (BMPs) are multi-functional growth factors that belong to the transforming growth factor b (TGFb) superfamily. To date, around 20 BMP family members have been identified and characterized. BMPs signal through serine/threonine kinase receptors, composed of type I and II subtypes. Three type I receptors have been shown to bind BMP ligands, type IA and IB BMP receptors (BMPR-IA or ALK-3 and BMPR-IB or ALK-6) and type IA activin receptor (ActR-IA or ALK-2) (Koenig, B.B. et al. (1994)
 (Koenig, B.B. et al. (1994)")
10
secreted BMP antagonists produced in the dorsal organizing center.
Ventral Spemann’s Organizer Neural ectoderm Epidermal ectoderm Mesoderm Endoderm At gastrula a gradient of BMP4 (a TGFβ-like growth factor) is established by a ventral source of BMP4, and a dorsal source of Chordin and Noggin, two secreted BMP antagonists produced in the dorsal organizing center.
 is established by. a ventral source of BMP4, and a dorsal source of Chordin and Noggin, two. secreted BMP antagonists produced in the dorsal organizing center.")
12
This is how organizing centers work in embryonic induction.
a morphogen gradient can be generated by a source of growth factor (such as BMP) or by a localized source of inhibitor (such as Chordin). This is how organizing centers work in embryonic induction.
 or by a localized source of inhibitor (such as Chordin). This is how organizing centers work in embryonic induction.")
13
Embryogenesis is controlled by surprisingly different signal transduction pathways:
TGFβ/BMP Serine/Threonine kinase receptors Receptor Tyrosine kinases such as FGF, EGF, IGF, Insulin Wnts Sonic Hedgehog Notch G protein-coupled receptors (7-transmembrane receptors) Nuclear hormone receptors Different signaling pathways pattern are active in the embryo, the same signals can trigger different types of cell differentiation responses in cells of different developmental history thus generating hundreds of differentiated cell types in the human body.
 Nuclear hormone receptors. Different signaling pathways pattern are active in the embryo, the same signals can trigger different types of cell differentiation responses in cells of different developmental history thus generating hundreds of differentiated cell types in the human body.")
14
A-P patterning outline:
2a) Hox genes: colinearity between the body plan and gene order in genomic DNA 2b) Hox genes and Retinoic acid 2c) Hox genes in the Evolution of Development
 Hox genes: colinearity between the body plan and gene order in genomic DNA. 2b) Hox genes and Retinoic acid. 2c) Hox genes in the Evolution of Development.")
15
Homeotic Mutations

16
Cloning of the homeobox (nucleic acid). Homeodomain refers to protein.
The homeodomain is a 60 aa DNA-binding domain that is very conserved during evolution. It fits into the major groove of the DNA. Define Hox, homeobox The term homeobox is reserved for the nucleic acid sequences that encode homeodomains. Since they are highly conserved, they can be detected by low-stringency hybridization across species.

17
Homeotic genes specify body segment identity in Drosophila.

18
Hox complexes are conserved between Drosophila and mammals (from De Robertis et al.,
Scientific American, 1990)
")
20
Vertebrates have four Hox complexes, with about 10 genes each
Vertebrates have four Hox complexes, with about 10 genes each. They can be aligned in 13 groups of paralogues. They display colinearity: Temporal colinearity: genes on one end of the complex are expressed first, those on the other (posterior) end are turned on last. Spatial colinearity: the more anteriorly expressed genes are in one end, the more posterior ones at the other end of the gene complex. Anterior Hox genes are activated sequentially by retinoic acid.
 end are turned on last. Spatial colinearity: the more anteriorly expressed genes are in one end, the more posterior ones at the other end of the gene complex. Anterior Hox genes are activated sequentially by retinoic acid.")
21
Homeobox (Hox) genes were originally discovered in the fruit fly Drosophila, where they function through a conserved omeodomain as transcriptional regulators to control embryonic morphogenesis. Since then over 1000 homeodomain proteins have been identified in several species. In vertebrates, 39 Hox genes have been identified as homologs of the original Drosophila complex, and like their Drosophila counterparts they are organized within chromosomal clusters. Vertebrate Hox genes have also been shown to play a critical role in embryonic development as transcriptional regulators. In vertebrates, more than 200 homeobox-containing genes have been identified

22
In Drosophila, a single homeotic complex (HOM-C) comprised of two separate clusters [Bithorax complex (BX-C) and Antennapedia complex (ANT-C)] is located on chromosome 3 In mice the Hox complex is comprised of 39 genes that are arranged into four separate chromosomal clusters designated Hox A, B, C, and D Vertebrate Hox and Drosophila HOM-C homeobox clusters is spatial colinearity, which is the expression of the Hox genes along the anteroposterior axis of the embryo in congruence with their arrangement along the chromosome . Thus, the 3’ genes within a chromosomal cluster are expressed more anteriorly than the 5’ genes with respect to the rostrocaudal axis of the embryo. In general, vertebrate Hox genes also display temporal colinearity and colinear sensitivity to RA, a known inducer of Hox gene expression
![In Drosophila, a single homeotic complex (HOM-C) comprised of two separate clusters [Bithorax complex (BX-C) and Antennapedia complex (ANT-C)] is located on chromosome 3](http://slideplayer.com/slide/3318165/11/images/22/In+Drosophila%2C+a+single+homeotic+complex+%28HOM-C%29+comprised+of+two+separate+clusters+%5BBithorax+complex+%28BX-C%29+and+Antennapedia+complex+%28ANT-C%29%5D+is+located+on+chromosome+3.jpg "In mice the Hox complex is comprised of 39 genes that are arranged into four separate chromosomal clusters designated Hox A, B, C, and D. Vertebrate Hox and Drosophila HOM-C homeobox clusters is spatial colinearity, which is the expression of the Hox genes along the anteroposterior axis of the embryo in congruence with their arrangement along the chromosome . Thus, the 3’ genes within a chromosomal cluster are expressed more anteriorly than the 5’ genes with respect to the rostrocaudal axis of the embryo. In. general, vertebrate Hox genes also display temporal colinearity and colinear sensitivity to RA, a known inducer of Hox gene expression.")
23
Ems empty spiracles (Emx in mammalian), transcription factors belonging to the orthodenticle family (Otx1, Otx2) play an important role during early and later events required for proper brain development. Otx1 is involved in corticogenesis, sense organ development and pituitary functions, while Otx2 is necessary earlier in development, for the correct anterior neural plate specification and organisation of the primitive streak. The first few processes that vertebrate embryos undergo - the division of the egg cytoplasm into cells (cleavage), the formation of the primary tissue layers (gastrulation) and the designation of embryonic coordinates (axis formation) do not seem to require the involvement of Hox genes and other anterior-posterior patterning genes. However, after the initiation of gastrulation, the subdivision of the embryo along the anterior-posterior axis begins. Concurrently, genes like the Hox genes, orthodenticle-related genes, and other homeobox-containing genes, start to be expressed in specific anterior-posterior domains. The Hox genes are expressed from the hindbrain posteriorly and turn on in an ordered sequential fashion, posterior to the domainof Otx2, an orthodenticle homolog.
, the formation of the primary tissue layers (gastrulation) and the designation of embryonic coordinates (axis formation) do not seem to require the involvement of Hox genes and other anterior-posterior patterning genes. However, after the initiation of gastrulation, the subdivision of the embryo along the anterior-posterior axis begins. Concurrently, genes like the Hox genes, orthodenticle-related genes, and other homeobox-containing genes, start to be expressed in specific anterior-posterior domains. The Hox genes are expressed from the hindbrain posteriorly and turn on in an ordered sequential fashion, posterior to the domainof Otx2, an orthodenticle homolog.")
25
Hox-C6 protein is seen in eight thoracic segments of the mouse embryo
Hox-C6 protein is seen in eight thoracic segments of the mouse embryo. Translation is blocked in the tail region, probably through the action of microRNAs. The inset shows that Hox-C6 mRNA is expressed all the way to the tip of the tail. From De Robertis, Cell 132, (2008) Fig. 18
 Fig. 18.")
26
Conservations between Drosophila and the four human Hox complexes
High RA response Low RA response From De Robertis, E.M. Evo-Devo: Variations on Ancestral themes. Cell 132, (2008)
")
27
Temporal and spatial colinearity: order of Hox genes in DNA follows the antero-posterior body axis.
Why have Hox genes stayed together in a complex?

28
2b) Retinoic acid activates HOX genes sequentially in cultured human teratocarcinoma cells
 Retinoic acid activates HOX genes sequentially in cultured human teratocarcinoma cells")
29
Retinoic acid receptor is a DNA-binding protein that works as a ligand-activated transcription factor. Many hydrophobic hormone receptors work in this way. Nuclear receptors work very differently from cell surface receptors. (RA) RA
 RA.")
30
Hox complexes have a retinoic acid receptor response element (RARE) in the DNA before paralogue 1. This DNA enhancer element controls expression of many genes in the complex. In retinoic acid teratogenesis, Hox gene expression borders move into more anterior regions. RARE

31
Hox genes and the development of body plans
Homeotic transformations in humans. A cervical vertebra transformed into a thoracic one with ribs. Hox knockouts in mice cause homeotic transformations, in this case an extra rib in the lumbar region (HoxC-8 mutant). Treatment with retinoic acid can also cause lumbar ribs. Fig. 11
. Treatment with retinoic acid can also cause lumbar ribs. Fig. 11.")
35
transcription factors of the
basic helix-loop-helix (bHLH) type.
 type.")
39
Numb vs Notch: drosophila vs Uomo

Similar presentations