Download presentation
Presentation is loading. Please wait.

2
Cell biology (72373) 2011 Amir Eden Department of Cell & Developmental Biology Institute of Life Sciences Room: 3-564 Email: eden@vms.huji.ac.ileden@vms.huji.ac.il The Cell Cycle Ubiquitin-Proteasome system Apoptosis
 2011 Amir Eden Department of Cell & Developmental Biology Institute of Life Sciences Room: The Cell Cycle Ubiquitin-Proteasome system Apoptosis")
3
DNA (gene) mRNA Protein Transcription RNA processing (splicing etc) Translation Folding Post translational modifications Peptides/amino acids Proteolysis
 mRNA Protein Transcription RNA processing (splicing etc) Translation Folding Post translational modifications Peptides/amino acids Proteolysis")
4
3 Cellular proteins are degraded at different rates. –Ornithine decarboxylase has a half-life of 11 minutes. –Hemoglobin lasts as long as a red blood cell. – Υ-Crystallin (eye lens protein) lasts as long as the organism does. Cellular Protein Degradation
 lasts as long as the organism does. Cellular Protein Degradation.")
5
15N labeled amino acids are incorporated into protein and then show up as amino acids

6
Over 50 hydrolytic enzymes. Common to the processes of degradation in the Intestine and lysosome that they do not require energy.

7
Lysosomes are dynamic organelles that receive and degrade macromolecules from the secretory, endocytic, autophagic and phagocytic membrane- trafficking pathways.

9
(Reticulocyte) Add (3H) labeled protein and monitor degraded fraction
 Add (3H) labeled protein and monitor degraded fraction")
10
The lysosome can’t be the only system that degrades proteins in the cell 1.Different proteins have different half lives - from minutes to days. 2.The degradation requires energy. 3.Distinct response of different proteins to lysosomal inhibitors.

11
Ubiquitin mediated degradation Avram Hershko Aaron Ciechanover 2004 Nobel prize in Chemistry Irwin rose

12
The breakthrough Establishment of in vitro system for proteolysis - enabled biochemical fractionation

13
(Reticulocyte) Add (3H) labeled protein and monitor degraded fraction
 Add (3H) labeled protein and monitor degraded fraction")
17
Multiple Ubiquitin genes encode Ub multimers or fusions with ribosomal proteins

22
K48

25
Procesive cycle to achieve poly-ubiquitination

26
Types of E3 ligases

29
Why have a 3-step ubiquitination process? Ubiquitin E1 (1) E2 (12-30) E3 (>200?) –HECT-type –RING-type –PHD-type –U-box containing
 E2 (12-30) E3 (>200 ) –HECT-type –RING-type –PHD-type –U-box containing.")
30
26S proteasome

31
30 The proteasome – the cell's waste disposer Proteasomes are barrel-formed structures. The black spots indicate active, protein-degrading surfaces within the barrel where it is shielded from the rest of the cell. The only way in to the active surface is via the “gate", which recognizes polyubiquitinated proteins. The proteosomes can break down practically all proteins to 7-9-amino- acid-long peptides. The peptides are released from the other end of the proteasome. A human cell contains about 30,000 proteasomes The proteasome itself cannot choose proteins, only ubiquitin-labelled proteins.

32
31 6) Proteasome-degradation. In eukaryotes 20S proteasome is able to break down unfolded target polypeptide. In eukaryotes 26S proteasome is required to attack a folded protein. Groll, M. et al. (1999) The catalytic sites of 20S proteasomes and their role in subunit maturation. A mutational and crystallographic study. PNAS 96: 10976
 The catalytic sites of 20S proteasomes and their role in subunit maturation. A mutational and crystallographic study. PNAS 96:")
33
Core: Processive! A collection of proteolytic enzymes: Chymotrypsin-like activity (preference for tyrosine or phenylalanine at the P1 (peptide cabonyl) position). Trypsin-like activity (preference for arginine or lysine at the P1 position). Post-glutamyl activity (preference for glutamate or other acidic residue at the P1 position)... Cap: multisubunits of which at least 6 Hydrolyze ATP to denature the protein and direct it to the core after removal of poly- Ub.
 position). Trypsin-like activity (preference for arginine or lysine at the P1 position). Post-glutamyl activity (preference for glutamate or other acidic residue at the P1 position)... Cap: multisubunits of which at least 6 Hydrolyze ATP to denature the protein and direct it to the core after removal of poly- Ub..")
34
Altogether: 26S proteasome

37
Why break down cellular proteins ? (Proteins that are imported via the plasma membrane are broken down (mostly lysosome)). Amino acids must be recycled. Regulation of protein level Proteins get denatured at some stage and must be disposed off Denatured proteins are poisonous for the cell. Proteins are often misfolded when synthesized - indeed a large fraction (~30%) of newly made proteins don’t pass the quality control and are degraded. 1.6 million peptides are broken down in each cell every minute !!
). Amino acids must be recycled. Regulation of protein level Proteins get denatured at some stage and must be disposed off Denatured proteins are poisonous for the cell. Proteins are often misfolded when synthesized - indeed a large fraction (~30%) of newly made proteins don’t pass the quality control and are degraded. 1.6 million peptides are broken down in each cell every minute !!.")
38
Why break down proteins ? Proteins get denatured at some stage and must be disposed of - denatured proteins are poisonous for the cell. Amino acids must be recycled.

39
How not to store proteins: 37oC (heat denatures) Small reactive molecules (They cause Oxidation, Deamination, Glycation) Other enzymes (proteases, phosphatases) Salts (denature), Fatty acids (Detergents) Other unfolded proteins (sticky) Intracellular environment is hostile to proteins
 Small reactive molecules (They cause Oxidation, Deamination, Glycation) Other enzymes (proteases, phosphatases) Salts (denature), Fatty acids (Detergents) Other unfolded proteins (sticky) Intracellular environment is hostile to proteins")
40
Why break down proteins ?

41
Mis-folded proteins are recognized, ubiquitinated and hence targeted to the Proteasome

43
ER associated degradation -ERAD Misfolded ER proteins are retrotranlocated to the cytoplams for proteasomal degradation

44
Abnormal proteins Protein Misfolding Incomplete Proteins Nonsense Mutations, Incorporation of Puromycin, Premature Termination Missense Proteins Mutations, Incorporation of Amino Acid Analogs, Biosynthetic Errors Free Subunits of Multimeric Complexes Excess Components Post-synthet ic Damage Oxygen Radicals, Intracellular Denaturation Genetic Engineering Gene Fusions, Frame-Shifts, Incorrect Localization

45
Why break down proteins ? As method for regulating various cellular processes. Cell cycle and division Differentiation and development Response to stress and extracellular modulators Morphogenesis of neuronal networks Modulation of cell surface receptors, ion channels and the secretory pathway, DNA repair Immune and inflammatory responses Biogenesis of organelles and apoptosis.

46
Specificity, specificity, specificity Regulation must be specific! Ubiquitin E1 (1) E2 (12-30) E3 (>200?) –HECT-type –RING-type –PHD-type –U-box containing
 E2 (12-30) E3 (>200 ) –HECT-type –RING-type –PHD-type –U-box containing.")
49
Signals for degradation (degrons) PEST sequences (Pro, Glu, Ser, Thr) FREQK nonessential under starvation conditions DUBS (de-ubiquinating enzymes) provide additional regulation
 PEST sequences (Pro, Glu, Ser, Thr) FREQK nonessential under starvation conditions DUBS (de-ubiquinating enzymes) provide additional regulation")
52
The N-end rule

53
Ubiquitin is involved in various neurodegenerative diseases like Alzheimer’s and Parkinson’s diseases

56
Aggresomes

59
Notch signaling CF- quality control ERAD

60
CFTR 508 CF- quality control ERAD

62
Antigen Presentation Peptide epitopes presented to cytotoxic T cells (CTLs) on class I MHC molecules are generated in the cytosol from limited processing of antigenic proteins. It is generally accepted that processing of most known MHC class I antigens is mediated by the ubiquitin–proteasome pathway

63
Antigen presentation

65
Ubiquitination is not only a degradation signal - mono-ubiquitination plays a role in regulation of various cellular processes from the histone code to trafficking. Ubiquitin binding protein domains: at least sixteen domains: UBA, UIM, MIU, DUIM, CUE, GAT, NZF, A20 ZnF, UBP ZnF, UBZ, Ubc, UEV, UBM, GLUE, Jab1/MPN and PFU.

69
UB ligase K63 E2 Substrates Recruitment of IKK (kinase)
")
70
And that’s only part of the story - there are ubiquitin-like proteins which also tag lysines of proteins and play a role in their regulation - SUMO, NEDD8

71
Ubiquitin-fold structures ubiquitin, Nedd8, UBX and ThiS all have ubiquitin folds (i.e., similar structures) ThiS is the most divergent, with only 14% sequence identity with ubiquitin; in comparison, Nedd8 is >50% identical all of the sequences contain C-terminal Gly-Gly residues that is used for conjugation; this Gly-Gly terminus is perfectly conserved ubiquitin Nedd8 UBX ThiS 18-3
 ThiS is the most divergent, with only 14% sequence identity with ubiquitin; in comparison, Nedd8 is >50% identical all of the sequences contain C-terminal Gly-Gly residues that is used for conjugation; this Gly-Gly terminus is perfectly conserved ubiquitin Nedd8 UBX ThiS 18-3")
77
Pathogenic aberrations in ubiquitiination

Similar presentations




























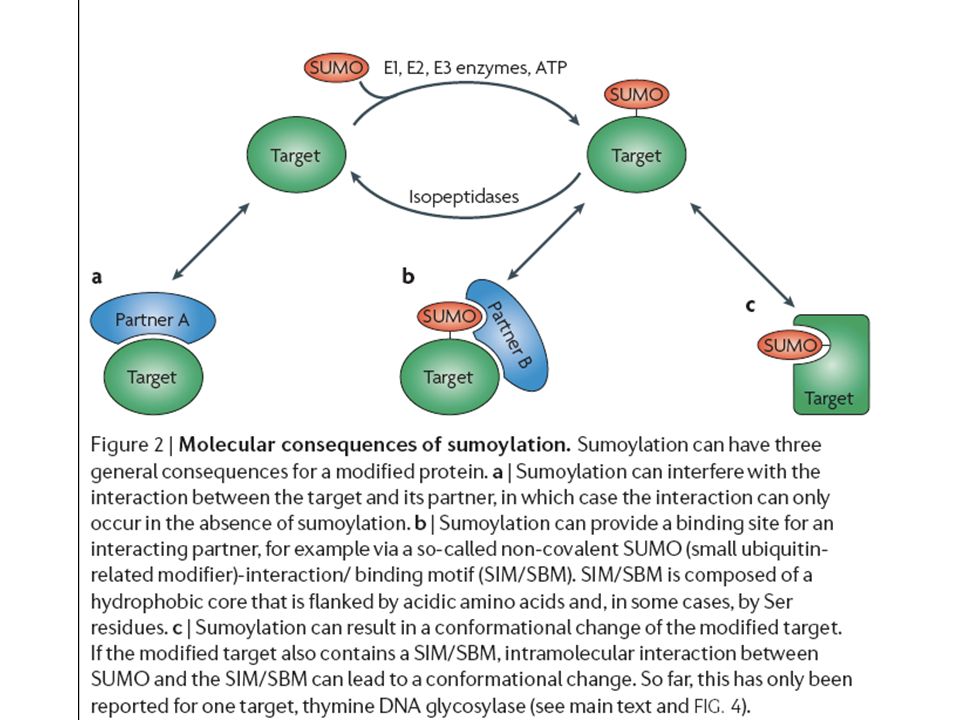

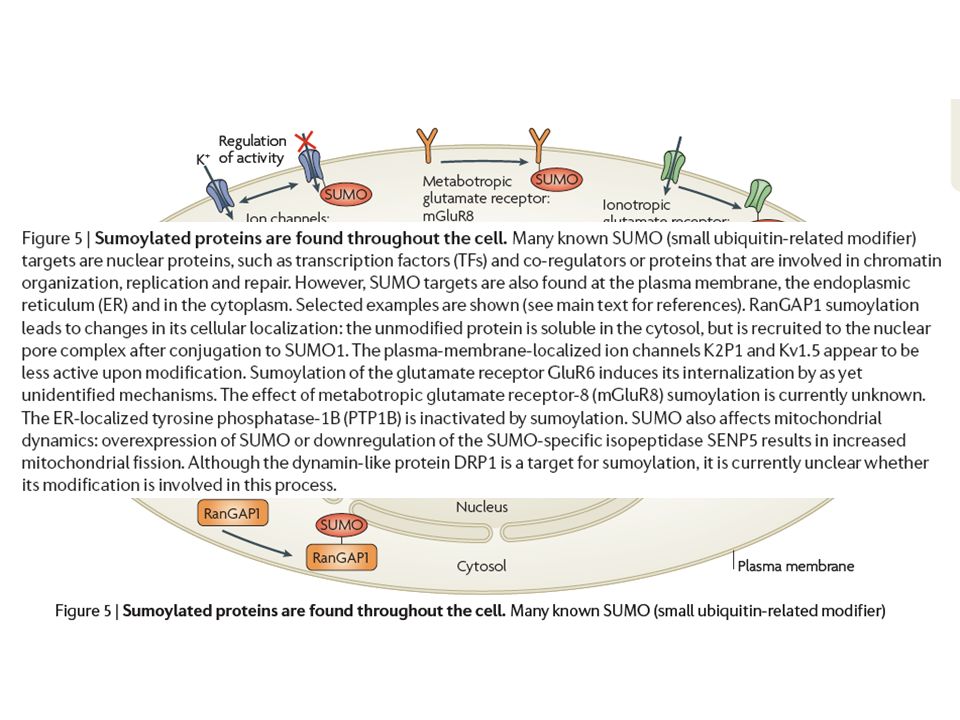






 Send the protein to a degradative compartment 2) selective degradation of individual molecules.>")








 Control of Protein Function Spectroscopy of Biopolymers.>")






