Download presentation
Presentation is loading. Please wait.

1
Proteomics and posttranslational modifications Xiaozhong Peng Department of Molecular Biology and Biochemistry National Laboratory of Medical Molecular Biology CAMS & PUMC

2
Protein Translation

3
Ribosome Structure

4
The Initiation of Translation

5
Mechanism of translation initiation. Preinitiation complex Initiation complex

6
Eukaryotic initiation complex CBP First AUG ( 5’ cap) 7-methyl- guanosine cap
 7-methyl- guanosine cap")
7
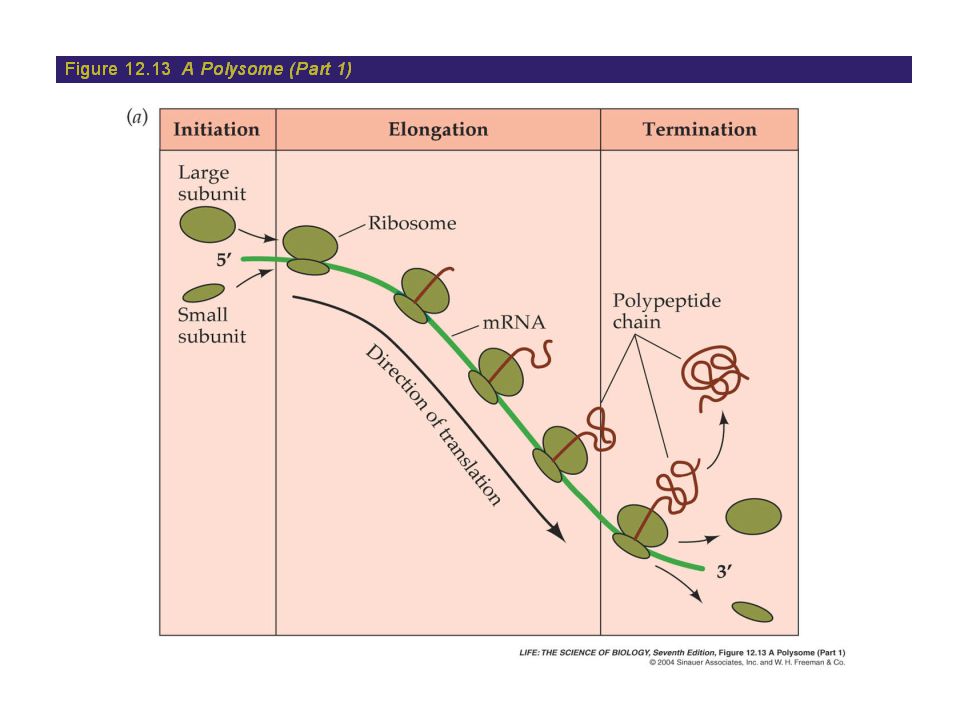
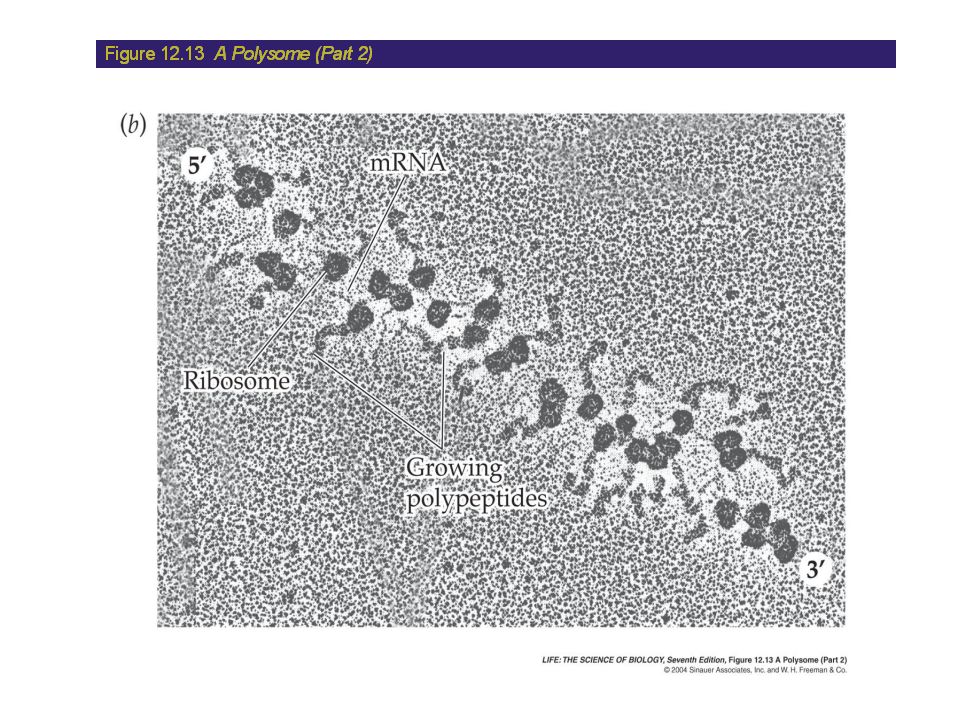
Translation: The Elongation Stage

8
The Termination of Translation

13
Pre-translation: take place at the level of amino acyl-tRNA prior to polymerization. Co-translation: take place during polymerization. Post-translation: take place after the completed protein has been released from the polysome.

14
Post-translational modifications

15
● N-terminal or C-terminal modification –Removal of N-formylmethionine –N-acetylation (50% of eucaryotic proteins) ● N-terminal and C-terminal processing –Maturation, proteolytic processing ● Modification of individual amino acids –Phosphorylation –Glycosylation –Methylation –Farnesylation ● Protein splicin: Intein
 ● N-terminal and C-terminal processing –Maturation, proteolytic processing ● Modification of individual amino acids –Phosphorylation –Glycosylation –Methylation –Farnesylation ● Protein splicin: Intein")
17
● More than 200 known posttranslational Modifications have been reported. --Gudepu, R.G.& Wold,F.(1998)in Proteins:Analysis and Design,ed.Angeletti, R.H.(Academic, San Diego),pp.121-207. ● More than 300 different types of PTMs are currently known and new ones are regularly discovered. --Ole Norregaard Jensen. Current Opinion in Chemical Biology 2004,8:33-41.
in Proteins:Analysis and Design,ed.Angeletti, R.H.(Academic, San Diego),pp ● More than 300 different types of PTMs are currently known and new ones are regularly discovered. --Ole Norregaard Jensen. Current Opinion in Chemical Biology 2004,8:")
18
● Protein Ubiquitination

19
The Nobel Prize in Chemistry 2004 "for the discovery of ubiquitin-mediated protein degradation" Aaron Ciechanover Technion – Israel Institute of Technology Haifa, Israel Avram Hershko Technion – Israel Institute of Technology Haifa, Israel Irwin Rose University of California Irvine, CA, USA

20
Proteins build up all living things: plants, animals and therefore us humans. In the past few decades biochemistry has come a long way towards explaining how the cell produces all its various proteins(at least five Nobel Prizes have been awarded in this area). But as to the breaking down of proteins, not so many researchers were interested.Aaron Ciechanover,Avram Hershko and Irwin Rose went against the stream and at the beginning of the 1980s discovered one of the cell's most important cyclical processes, regulated protein degradation. For this, they are being rewarded with this year's Nobel Prize in Chemistry. This year's Nobel Laureates in chemistry, Aaron Ciechanover, Avram Hershko and Irwin Rose, have contributed ground-breaking chemical knowledge of how the cell can regulate the presence of a certain protein by marking unwanted proteins with a label consisting of the polypeptide ubiquitin. Proteins so labelled are then broken down – degraded – rapidly in cellular "waste disposers" called proteasomes.
. But as to the breaking down of proteins, not so many researchers were interested.Aaron Ciechanover,Avram Hershko and Irwin Rose went against the stream and at the beginning of the 1980s discovered one of the cell s most important cyclical processes, regulated protein degradation. For this, they are being rewarded with this year s Nobel Prize in Chemistry. This year s Nobel Laureates in chemistry, Aaron Ciechanover, Avram Hershko and Irwin Rose, have contributed ground-breaking chemical knowledge of how the cell can regulate the presence of a certain protein by marking unwanted proteins with a label consisting of the polypeptide ubiquitin. Proteins so labelled are then broken down – degraded – rapidly in cellular waste disposers called proteasomes..")
21
Protein Degradation: Schoenheimer : a pioneer in this field! 1942--isotope tracer techniques—indicated that proteins in animals are continuously synthesized and degraded and therefore are in a Dynamic state. Degradation needs no energy-or does it? It doesn’t: Trypsin: a type of cell organelle: Lysosome It does: 1.Simpson, 1953: release of amino acids from cultured liver slices was energy-dependent. 2.Hershko and Tomkin, 1971: energy-dependent degradation of the Enzyme tyrosine aminotransferase in cultured hepatoma cells. 3.Ciechanover, 1997: tyrosine aminotransferase degradation is indeed ubiquitin-mediated.

22
Fig 1. Ubiquitin - a common polypeptide that represents the "kiss of death ". The Label is ubiquitin: ● Was first isolated from bovine thymus (calf sweetbread) by Goldstein in 1975. ● Busch found “protein A24”-histone H2A+ubiquitin.??? ● Hunt and Dayhoff found in 1977.Named from Latin ubique, “everywhere”. ● 76 amino acids peptide.found in numerous different tissues and organisms-but not in bacteria.
 by Goldstein in ● Busch found protein A24 -histone H2A+ubiquitin. . ● Hunt and Dayhoff found in 1977.Named from Latin ubique, everywhere . ● 76 amino acids peptide.found in numerous different tissues and organisms-but not in bacteria..")
23
The discovery of ubiquitin-mediated protein degradation: ● A major part of the work was done during a series of sabbatical leaves when Hershko and Ciechanover worked in Rose’s laboratory at the Fox Chase Cancer Center in Philadephia. Two surprising discoveries: ● in 1978, when Reticulocyte lysate system was passed over a DEAE cellulose column to remove the hemmoglobin, two fractions one contains APF-1(active principle of fraction1)-ubiquitin. ● in 1979,the second fractions subdivided by salt precipitation into two :one contains 450kDa protein-proteasome, and another contains E1-E3 enzymes. The Breakthrough in 1980: 125 I-Labeled APF-1 125 I-Labeled lysozyme, a-lactalbumin and globin
-ubiquitin. ● in 1979,the second fractions subdivided by salt precipitation into two :one contains 450kDa protein-proteasome, and another contains E1-E3 enzymes. The Breakthrough in 1980: 125 I-Labeled APF I-Labeled lysozyme, a-lactalbumin and globin.")
24
Two novel enzymatic activities:

25
Fig 2. Ubiquitin-mediated protein degradation 1. The E1 enzyme activates the ubiquitin molecule. This reaction requires energy in the form of ATP. 2. The ubiquitin molecule is transferred to a different enzyme, E2. 3. The E3 enzyme can recognise the protein target which is to be destroyed. The E2-ubiquitin complex binds so near to the protein target that the actual ubiquitin label can be transferred from E2 to the target. 4. The E3 enzyme now releases the ubiquitin-labelled protein. 5. This last step is repeated until the protein has a short chain of ubiquitin molecules attached to itself. 6. This ubiquitin chain is recognised in the opening of the proteasome. The ubiquitin label is disconnected and the protein is admitted and chopped into small pieces. Multi-step ubiquitin-tagging hypothesis:

26
Fig 3. The cell's waste disposer, the proteasome. The black spots indicate active, protein-degrading surfaces. The Proteasome-the cell’s waste disposer ● A human cell contains about 30,000 proteasomes, can break down practically all proteins to 7-9-aa-long peptides.

27
More recent research:

28
● Regulation of the cell cycle( for example) APC: anaphase-promoting complex
 APC: anaphase-promoting complex")
30
To identify the E3 ligase responsible for H2A ubiquitination, They Developed an assay: in the presence of E1,E2,ATP and Flag-Ubiquitin(FUb), Hela nuclear protein fractions were tested for E3 Ligase activity with core histone or nucleosomal histone substrates.
, Hela nuclear protein fractions were tested for E3 Ligase activity with core histone or nucleosomal histone substrates.")
34
Chromatin Immunoprecipitation Immunoprecipitation PCR Identification The schematic principle of ChIP DNA-Protein Interaction Histone Modification

35
Summary: ● Purified an E3 ubiquitin ligase complex that is specific for histone H2A. ● The complex, termed hPRC1L(human Polycomb repressive complex1-like), is composed of several Polycomb-group proteins including Ring1, Ring2, Bmi1and HPH2. ● hPRC1L monoubiquitinates nucleosomal histone H2A at lysine 119. ● Reducing the expression of Ring2 results in a dramatic decrease in the level of ubiquitinated H2A in HeLa cells. ● Chromatin immunoprecipitation analysis demonstrated colocalization of dRing with ubiquitinated H2A at the PRE and promoter regions of the Drosophila Ubx gene in wing imaginal discs. ● Removal of dRing in SL2 tissue culture cells by RNA interference resulted in loss of H2A ubiquitination concomitant with derepression of Ubx. Thus, their studies identify the H2A ubiquitin ligase, and link H2A ubiquitination to Polycomb silencing.
, is composed of several Polycomb-group proteins including Ring1, Ring2, Bmi1and HPH2. ● hPRC1L monoubiquitinates nucleosomal histone H2A at lysine 119. ● Reducing the expression of Ring2 results in a dramatic decrease in the level of ubiquitinated H2A in HeLa cells. ● Chromatin immunoprecipitation analysis demonstrated colocalization of dRing with ubiquitinated H2A at the PRE and promoter regions of the Drosophila Ubx gene in wing imaginal discs. ● Removal of dRing in SL2 tissue culture cells by RNA interference resulted in loss of H2A ubiquitination concomitant with derepression of Ubx. Thus, their studies identify the H2A ubiquitin ligase, and link H2A ubiquitination to Polycomb silencing..")
36
Ubiquitin like molecule:

37
● Protein Glycosylations

39
Figure 12.15 A Signal Sequence Moves a Polypeptide into the ER (Part 1)
")
40
Figure 12.15 A Signal Sequence Moves a Polypeptide into the ER (Part 2)
")
Similar presentations














 Control of Protein Function Spectroscopy of Biopolymers.>")












 Genes.>")