Download presentation
Presentation is loading. Please wait.

1
These 2D methods work for proteins up to about 100 amino acids, and even here, anything from 50-100 amino acids is difficult. We need to reduce the complexity of these 2D spectra. We can start by replacing 14 N with 15 N, a spin 1/2 nucleus.

2
1 H- 15 N HSQC of rat FAS ACP 1 H Chemical Shift 15N shift of nitrogen of amide bond X 89!

3
Simplifying the fingerprint region with 15 N edited NOESY and TOCSY spectra

4
These methods take advantage of large 1 J coupling constants

5
HNCA Backbone assignment via 1 J couplings

6
HN(CO)CA
CA")
7
Slice from HNCA (at the 15 N shift of I44, T14, R74..). Each pair of peaks correlates a C i) and C i-1) with the 1 H and 15 N shift of residue i. Slice from HN(CO)CA (at the 15 N shift of I44, T14, R74..). Each pair of peaks correlates the C i-1) with the 1 H and 15 N shift of residue i.
 and C i-1) with the 1 H and 15 N shift of residue i. Slice from HN(CO)CA (at the 15 N shift of I44, T14, R74..). Each pair of peaks correlates the C i-1) with the 1 H and 15 N shift of residue i..")
8
An example. 13 C shifts of Isoleucine We know the 13 C shifts from the backbone assignment

9
Stage 2. Sidechain assignments completed with HCCH-COSY and HCCH-TOCSY for example. The HCCH experiments provide connectivities of the aliphatic side chains of individual amino acid residues. Complete assignments can be obtained if the backbone assignments and the side-chain assignments can be connected via the 13 C shifts.

10
Attempt to gain complete 1 H, 15 N and 13 C chemical shift assignments. We can now resolve uncertainty in NOEs we observe. These 4 methyls would give an ambiguous network of possible NOEs. But suppose we knew that the 13 C shift of the CH3 of Ile 1 was 9.3ppm and the CH3 of Ile 2 was 13 ppm.

11
Far larger proteins can now be tackled…44kDa Simian immuodeficiency virus (SIV) ectodomain used to fuse with host white blood cells
 ectodomain used to fuse with host white blood cells")
12
Types of Spin Relaxation Longitudinal or spin-lattice relaxation (T 1 ) - recovery of longitudinal magnetization - establishment of thermal equilibrium populations - exchange of energy Transverse or spin-spin relaxation (T 2 ) -decay of transverse magnetization - no exchange of energy - increase of entropy
 - recovery of longitudinal magnetization - establishment of thermal equilibrium populations - exchange of energy Transverse or spin-spin relaxation (T 2 ) -decay of transverse magnetization - no exchange of energy - increase of entropy")
13
Precession of Transverse Magnetization The transverse magnetization components oscillate and decay MxMx MyMy Time x y z x y z x y z BoBo xy plane M y (t) = -M z eq cos( t) exp{-t / T 2 } M x (t) = M z eq sin( t) exp{-t / T 2 } oscillation at the Larmor frequency decay time constant = spin-spin relaxation time OR transverse relaxation time
 = -M z eq cos( t) exp{-t / T 2 } M x (t) = M z eq sin( t) exp{-t / T 2 } oscillation at the Larmor frequency decay time constant = spin-spin relaxation time OR transverse relaxation time")
14
Transverse relaxation or T 2 decay transverse magnetization is excited by first pulse along –y-axis transverse magnetization dephases due to field inhomogeneity during the interval /2. “Black” vectors rotate faster than “grey” vectors

15
T 1. Build up of longitudinal magnetization when field is switched on M z (t) = M z eq [1- exp{- (t-t on ) / T 1 }] Equilibrium longitudinal magnetization Spin-lattice relaxation time OR longitudinal relaxation time
 = M z eq [1- exp{- (t-t on ) / T 1 }] Equilibrium longitudinal magnetization Spin-lattice relaxation time OR longitudinal relaxation time.")
16
Inversion of longitudinal magnetization by π pulse 180 o rotation about x-axis Recovery of longitudinal magnetization after π pulse 1 2

17
Simple theory of T 1 rotational correlation time mean square amplitude of fluctuating fields spin-lattice relaxation rate constant Larmor frequency rotational correlation time [in ns] approx. equal to 0.5 molecular mass [in kDa] 1 kDa = 1000 atomic mass units large molecules tumble more slowly small molecules tumble more quickly Rotational correlation time c
![Simple theory of T 1 rotational correlation time mean square amplitude of fluctuating fields spin-lattice relaxation rate constant Larmor frequency rotational correlation time [in ns] approx.](http://images.slideplayer.com/37/10684335/slides/slide_17.jpg "equal to 0.5 molecular mass [in kDa] 1 kDa = 1000 atomic mass units large molecules tumble more slowly small molecules tumble more quickly Rotational correlation time c.")
18
Comparison of T 1 and T 2 rapid motion (small molecule non-viscous liquids), T 1 and T 2 are equal Slow motion (large molecules, viscous liquids): T 2 is shorter than T 1.
, T 1 and T 2 are equal Slow motion (large molecules, viscous liquids): T 2 is shorter than T 1.")
19
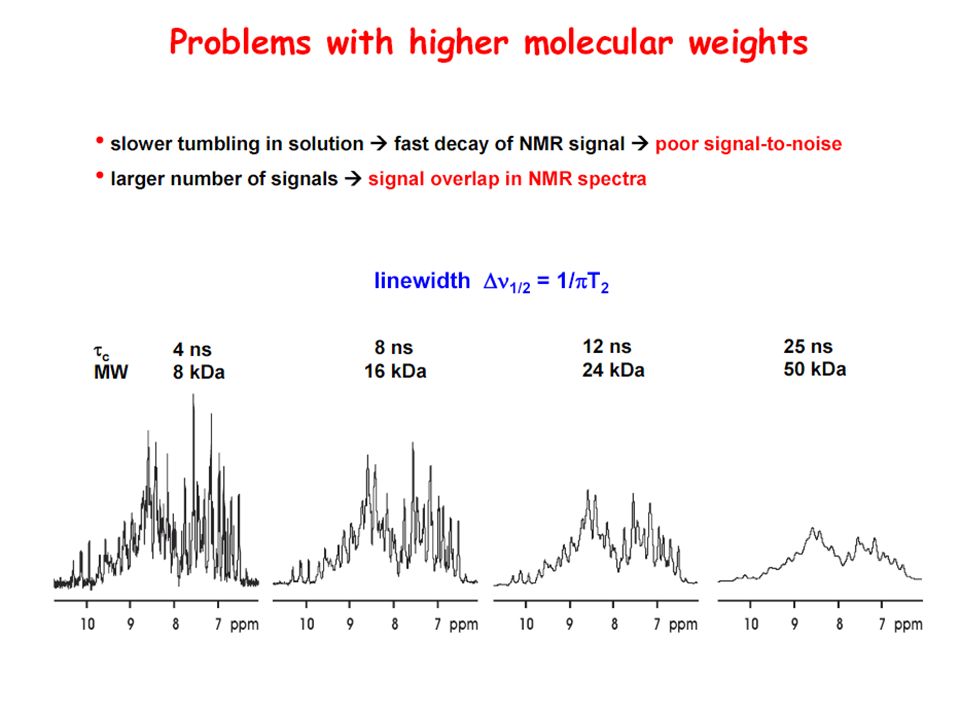
Problems with higher molecular weights and how to overcome them is the line-width in Hz at half peak height

20
Pg 46 & 47 of Rattle

Similar presentations




 (Fall Term, 2005) Department of Chemistry National Sun Yat-sen University 無機物理方法(核磁共振部分)>")





>")








