Download presentation
Presentation is loading. Please wait.

1
The Urea Cycle

3
TCA cycle

4
HOW UREA CYCLE WORKS?

11
kidney urine carbamoyl phosphate synthetase-I ornithine transcarbamoylase.

12
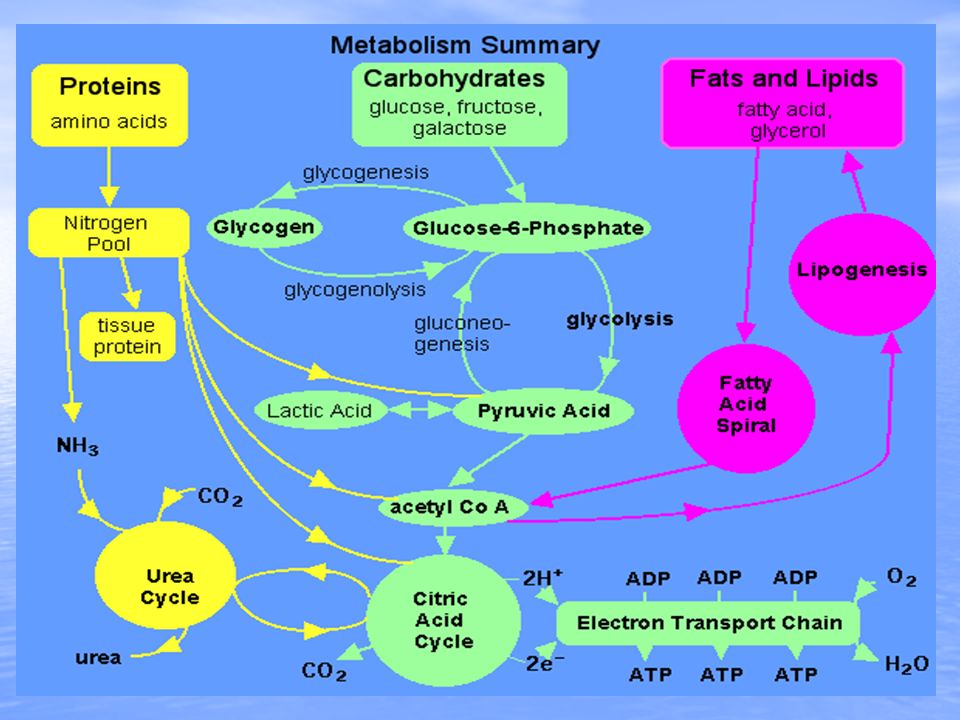
Urea is the major end product of nitrogen metabolism in humans and mammals. Ammonia, the product of oxidative deamination reactions, is toxic in even small amounts and must be removed from the body. The urea cycle or the ornithine cycle describes the conversion reactions of ammonia into urea. Since these reactions occur in the liver, the urea is then transported to the kidneys where it is excreted. The overall urea formation reaction is: 2 Ammonia + carbon dioxide + 3ATP ---> urea + water + 3 ADP

13
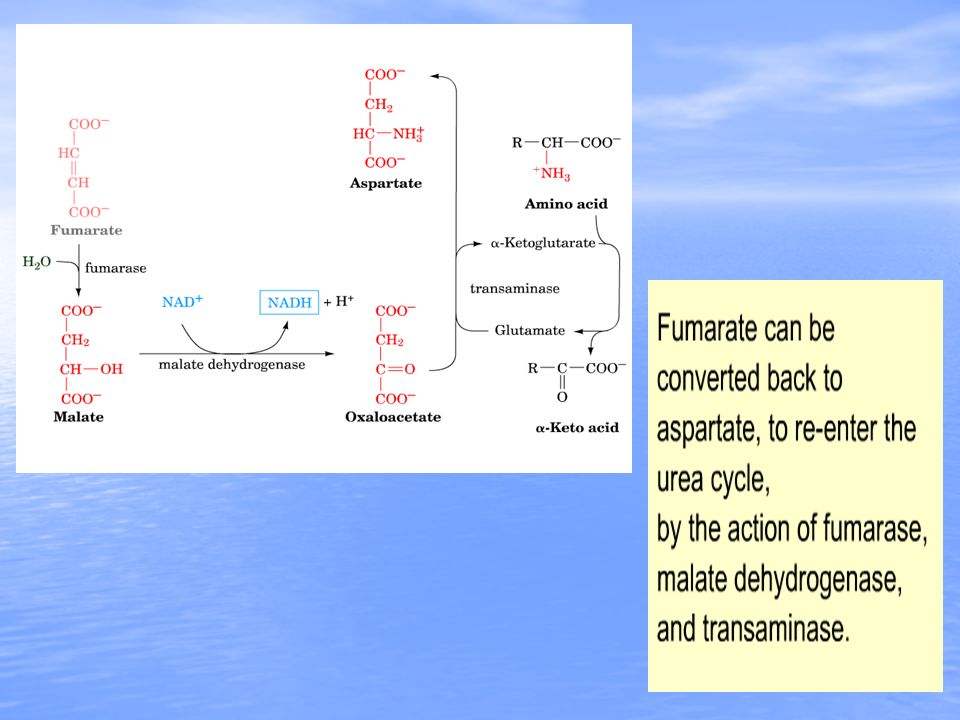
The step wise process of the urea cycle is summarized in the graphic. One amine group comes from oxidative deamination of glutamic acid while the other amine group comes from aspartic acid. Aspartic acid is regenerated from fumaric acid produced by the urea cycle. The fumaric acid first undergoes reactions through a portion of the citric acid cycle to produce oxaloacetic acid which is then changed by transamination into aspartic acid.

14
Urea is routinely measured in the blood as: Blood Urea Nitrogen (BUN). BUN levels may be elevated (a condition called uremia) in both acute and chronic renal (kidney) failure. Various diseases damage the kidney and cause faulty urine formation and excretion. Congestive heart failure leads to a low blood pressure and consequent reduced filtration rates through the kidneys, therefore, BUN may be elevated. Urinary tract obstructions can also lead to an increased BUN. In severe cases, hemodialysis is used to remove the soluble urea and other waste products from the blood. Waste products diffuse through the dialyzing membrane because their concentration is lower in the dialyzing solution. Ions, such as Na+ and Cl- which are to remain in the blood, are maintained at the same concentration in the dialyzing solution - no net diffusion occurs.
 in both acute and chronic renal (kidney) failure. Various diseases damage the kidney and cause faulty urine formation and excretion. Congestive heart failure leads to a low blood pressure and consequent reduced filtration rates through the kidneys, therefore, BUN may be elevated. Urinary tract obstructions can also lead to an increased BUN. In severe cases, hemodialysis is used to remove the soluble urea and other waste products from the blood. Waste products diffuse through the dialyzing membrane because their concentration is lower in the dialyzing solution. Ions, such as Na+ and Cl- which are to remain in the blood, are maintained at the same concentration in the dialyzing solution - no net diffusion occurs..")
15
As stated previously, high ammonia levels are toxic to humans. A complete block of any step in the urea cycle is fatal since there is no known alternative pathway for the synthesis of urea. Inherited disorders from defective enzymes may cause a partial block in some of the reactions and results in hyperammonemia which can lead to mental retardation. Extensive ammonia accumulation leads to extensive liver damage and death. Liver cirrhosis caused by alcoholism creates an interference in the enzymes which produce carbamyl phosphate in the first step on the cycle.

16
Ammonia Excretion Animals such as fish that live in the water excrete ammonia. However, even fish do not have large concentrations of ammonia in the blood because it is excreted as the amide in glutamine. Glutamine is carried to a membrane in the gills near the surrounding water where hydrolysis of the glutamine to glutamic acid releases the ammonia. Humans also excrete small amounts of NH 3 which is converted by the acid in the urine to NH 4 + (ammonium ions). Ammonia is excreted only as a defense against an acidosis condition because the neutralization of acid by ammonia has the effect of conserving bases in the blood.
. Ammonia is excreted only as a defense against an acidosis condition because the neutralization of acid by ammonia has the effect of conserving bases in the blood..")
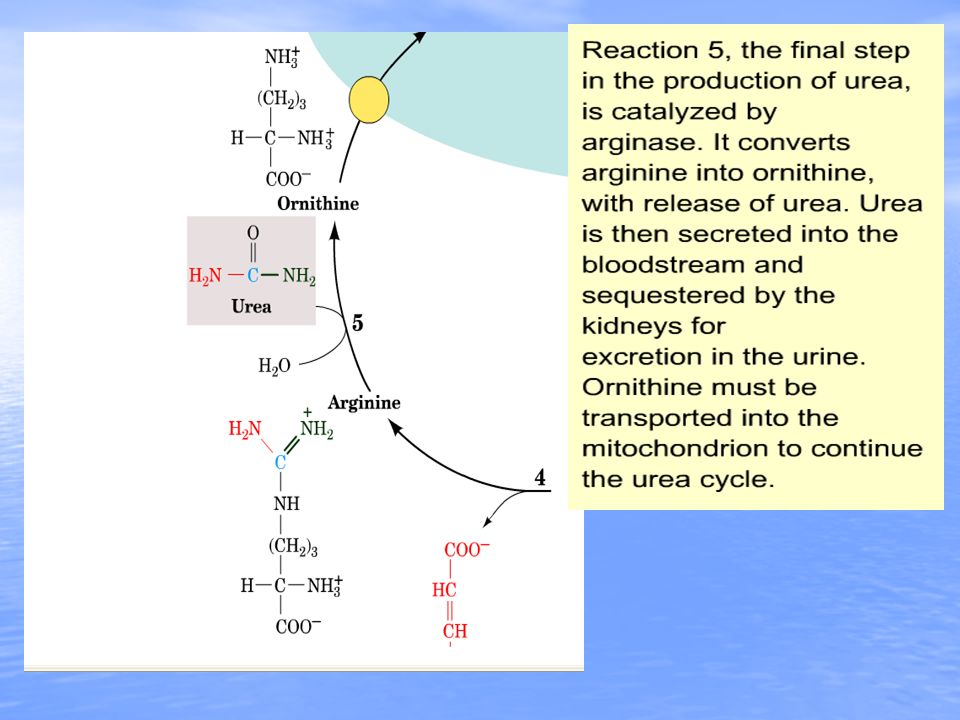
17
Uric Acid Excretion In animals that excrete urea, water is required to dissolve urea in urine. Actually, urea is a natural diuretic which insures the flow of urine. Animals such as birds and reptiles which must conserve water, excrete nitrogen as uric acid as a solid mixed with very small amounts of water. The conversion of ammonia and the synthesis of uric acid in birds and reptiles is a complicated energy requiring process. In humans, uric acid is not produced from ammonia but is synthesized from adenine and guanine found in various nucleotides.

18
Uric acid is normally excreted in small amounts in the urine of humans. Uric acid concentrations may become elevated in kidney diseases and leukemia. Uric acid salts maybe deposited in the form of kidney and bladder stones. A painful arthritic disorder called gout results from the deposition of uric acid salts in cartilage in the joints. Gout is probably caused by a variety of inborn errors of metabolism resulting in excessive synthesis of uric acid. Gout can be controlled by diet and/or a drug called allopurinol which inhibits the enzyme which produces uric acid.

20
Urea is the chief nitrogenous waste of mammals. Urea is the chief nitrogenous waste of mammals. Most of our nitrogenous waste comes from the breakdown of amino acids. Most of our nitrogenous waste comes from the breakdown of amino acids.amino acidsamino acids This occurs by deamination. Deamination of amino acids results in the production of ammonia (NH 3 ). This occurs by deamination. Deamination of amino acids results in the production of ammonia (NH 3 ). The Urea Cycle
. This occurs by deamination. Deamination of amino acids results in the production of ammonia (NH 3 ). The Urea Cycle.")
21
Ammonia is an extremely toxic base and its accumulation in the body would quickly be fatal.base However, the liver contains a system of carrier molecules and enzymes which quickly converts the ammonia (and carbon dioxide) into urea. This is called the urea cycle.

22
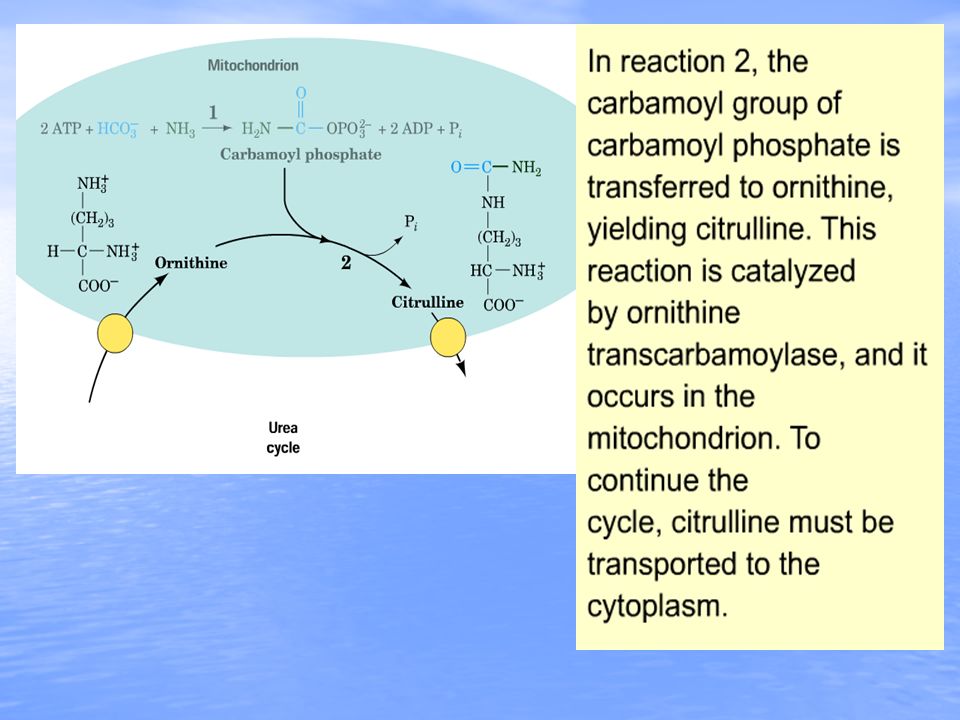
The essential features of the urea cycle reactions and their metabolic regulation are as follows: The essential features of the urea cycle reactions and their metabolic regulation are as follows: Arginine from the diet or from protein breakdown is cleaved by the cytosolic enzyme arginase, generating urea and ornithine. In subsequent reactions of the urea cycle a new urea residue is built on the ornithine, regenerating arginine and perpetuating the cycle. Arginine from the diet or from protein breakdown is cleaved by the cytosolic enzyme arginase, generating urea and ornithine. In subsequent reactions of the urea cycle a new urea residue is built on the ornithine, regenerating arginine and perpetuating the cycle. Ornithine arising in the cytosol is transported to the mitochondrial matrix, where ornithine transcabamoylase catalyzes the condensation of ornithine with carbamoyl phosphate, producing citrulline. Ornithine arising in the cytosol is transported to the mitochondrial matrix, where ornithine transcabamoylase catalyzes the condensation of ornithine with carbamoyl phosphate, producing citrulline. The energy for the reaction is provided by the high- energy anhydride of carbamoyl phosphate. The product, citrulline, is then transported to the cytosol, where the remaining reactions of the cycle take place. The energy for the reaction is provided by the high- energy anhydride of carbamoyl phosphate. The product, citrulline, is then transported to the cytosol, where the remaining reactions of the cycle take place.

23
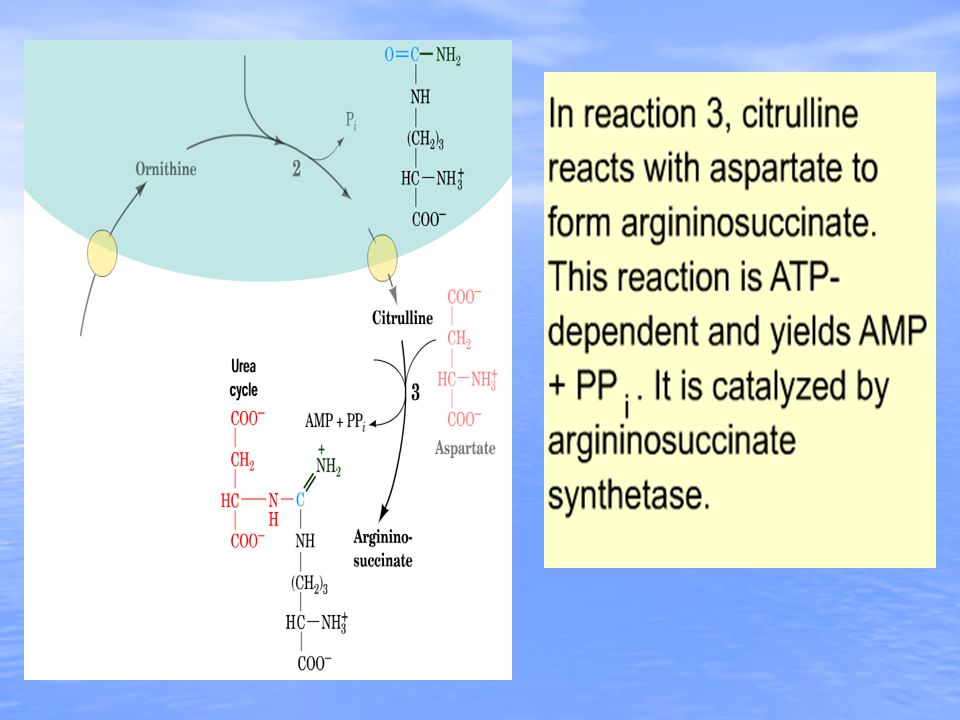
The synthesis of citrulline requires a prior activation of carbon and nitrogen as carbamoyl phosphate (CP). The activation step requires 2 equivalents of ATP and the mitochondrial matrix enzyme carbamoyl phosphate synthetase-I (CPS-I). There are two CP synthetases: a mitochondrial enzyme, CPS-I, which forms CP destined for inclusion in the urea cycle, and a cytosolic CP synthatase (CPS-II), which is involved in pyrimidine nucleotide biosynthesis. CPS-I is positively regulated by the allosteric effector N-acetyl-glutamate, while the cytosolic enzyme is acetylglutamate independent.pyrimidine nucleotide biosynthesis In a 2-step reaction, catalyzed by cytosolic argininosuccinate synthetase, citrulline and aspartate are condensed to form argininosuccinate. The reaction involves the addition of AMP (from ATP) to the amido carbonyl of citrulline, forming an activated intermediate on the enzyme surface (AMP-citrulline), and the subsequent addition of aspartate to form argininosuccinate.
. There are two CP synthetases: a mitochondrial enzyme, CPS-I, which forms CP destined for inclusion in the urea cycle, and a cytosolic CP synthatase (CPS-II), which is involved in pyrimidine nucleotide biosynthesis. CPS-I is positively regulated by the allosteric effector N-acetyl-glutamate, while the cytosolic enzyme is acetylglutamate independent.pyrimidine nucleotide biosynthesis In a 2-step reaction, catalyzed by cytosolic argininosuccinate synthetase, citrulline and aspartate are condensed to form argininosuccinate. The reaction involves the addition of AMP (from ATP) to the amido carbonyl of citrulline, forming an activated intermediate on the enzyme surface (AMP-citrulline), and the subsequent addition of aspartate to form argininosuccinate..")
24
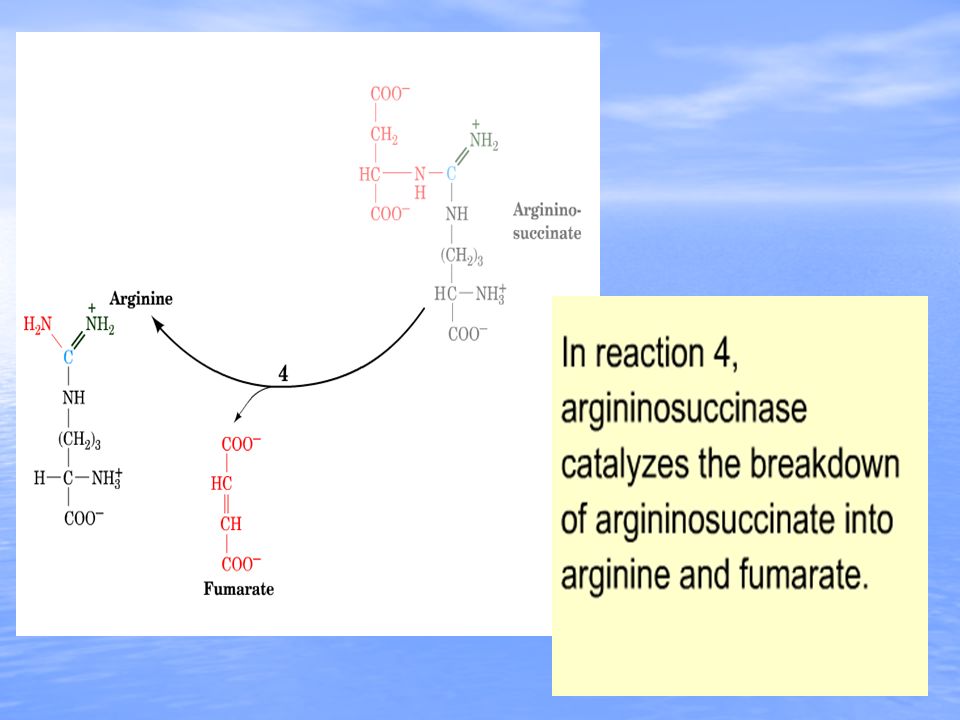
Arginine and fumarate are produced from argininosuccinate by the cytosolic enzyme argininosuccinate lyase (also called argininosuccinase). In the final step of the cycle arginase cleaves urea from aspartate, regenerating cytosolic ornithine, which can be transported to the mitochondrial matrix for another round of urea synthesis. The fumarate, generated via the action of arginiosuccinate lyase, is reconverted to aspartate for use in the argininosuccinate synthetase reaction. This occurs through the actions of cytosolic versions of the TCA cycle enzymes, fumarase (which yields malate) and malate dehydrogenase (which yields oxaloacetate). The oxaloacetate is then transaminated to aspartate by AST.TCA cycle Beginning and ending with ornithine, the reactions of the cycle consumes 3 equivalents of ATP and a total of 4 high-energy nucleotide phosphates. Urea is the only new compound generated by the cycle; all other intermediates and reactants are recycled. The energy consumed in the production of urea is more than recovered by the release of energy formed during the synthesis of the urea cycle intermediates. Ammonia released during the glutamate dehydrogenase reaction is coupled to the formation of NADH. In addition, when fumarate is converted back to aspartate, the malate dehydrogenase reaction used to convert malate to oxaloacetate generates a mole of NADH. These two moles of NADH, thus, are oxidized in the mitochondria yielding 6 moles of ATP.
 and malate dehydrogenase (which yields oxaloacetate). The oxaloacetate is then transaminated to aspartate by AST.TCA cycle Beginning and ending with ornithine, the reactions of the cycle consumes 3 equivalents of ATP and a total of 4 high-energy nucleotide phosphates. Urea is the only new compound generated by the cycle; all other intermediates and reactants are recycled. The energy consumed in the production of urea is more than recovered by the release of energy formed during the synthesis of the urea cycle intermediates. Ammonia released during the glutamate dehydrogenase reaction is coupled to the formation of NADH. In addition, when fumarate is converted back to aspartate, the malate dehydrogenase reaction used to convert malate to oxaloacetate generates a mole of NADH. These two moles of NADH, thus, are oxidized in the mitochondria yielding 6 moles of ATP..")
25
Regulation of the Urea Cycle The urea cycle operates only to eliminate excess nitrogen. On high-protein diets the carbon skeletons of the amino acids are oxidized for energy or stored as fat and glycogen, but the amino nitrogen must be excreted. To facilitate this process, enzymes of the urea cycle are controlled at the gene level. With long-term changes in the quantity of dietary protein, changes of 20-fold or greater in the concentration of cycle enzymes are observed. When dietary proteins increase significantly, enzyme concentrations rise. On return to a balanced diet, enzyme levels decline. Under conditions of starvation, enzyme levels rise as proteins are degraded and amino acid carbon skeletons are used to provide energy, thus increasing the quantity of nitrogen that must be excreted. Short-term regulation of the cycle occurs principally at CPS-I, which is relatively inactive in the absence of its allosteric activator N- acetylglutamate. The steady-state concentration of N-acetylglutamate is set by the concentration of its components acetyl-CoA and glutamate and by arginine, which is a positive allosteric effector of N-acetylglutamate synthetase. acetyl-CoA + glutamate ----> N-acetylglutamate + CoA

26
Neurotoxicity Associated with Ammonia Earlier it was noted that ammonia was neurotoxic. Marked brain damage is seen in cases of failure to make urea via the urea cycle or to eliminate urea through the kidneys. The result of either of these events is a buildup of circulating levels of ammonium ion. Aside from its effect on blood pH, ammonia readily traverses the brain blood barrier and in the brain is converted to glutamate via glutamate dehydrogenase, depleting the brain of α-KG. As the α-KG is depleted, oxaloacetate falls correspondingly, and ultimately TCA cycle activity comes to a halt. In the absence of aerobic oxidative phosphorylation and TCA cycle activity, irreparable cell damage and neural cell death ensue. In addition, the increased glutamate leads to glutamine formation. This depletes glutamate stores which are needed in neural tissue since glutamate is both a neurotransmitter and a precursor for the synthesis of γ-aminobutyrate: GABA, another neurotransmitter. Therefore, reductions in brain glutamate affect energy production as well as neurotransmission.γ-aminobutyrate: GABA Additional untoward consequences are the result of elevations in neural glutamine concentration. Glial cell (astrocytes) volume is controlled by intracellular organic osmolyte metabolism. The organic osmolyte is glutamine. As glutamine levels rise in the brain the volume of fluid within glial cells increases resulting in the cerebral edema seen in infants with hyperammonemia caused by urea cycle defects.
 volume is controlled by intracellular organic osmolyte metabolism. The organic osmolyte is glutamine. As glutamine levels rise in the brain the volume of fluid within glial cells increases resulting in the cerebral edema seen in infants with hyperammonemia caused by urea cycle defects..")
Similar presentations






>")



In stomach: passage of food into stomach stimulates gastric mucosa to secret a polypeptide hormone called: Gastrin.>")


 about glutamine and alanine? A) They are nitrogen donors in.>")







