Download presentation
Presentation is loading. Please wait.

1
THE IMMUNE RESPONSES TO VIRUSES

3
Replication of Ebola Virus
1. Attachment First of all, there is attachment of virus to host receptors through GP glycoprotein which is endocytosed into vesicles in the host cell. Host DC-SIGN and DC-SIGNR play a role in virion attachment. 2. Viral Entry (Penetration) The virion enters early endosomes by Macropinocytosis or clathrin-mediated endocytosis. A. Macropinocytosis In this process, ruffled segments of the host’s plasma membrane protrude outward from the cell and form invaginations where the virus utilizes glycoproteins in order to attach to the surface of the plasma membrane. Macropinocytosis is a process in which the Eukaryotic host cells form macropinosomes, segments of plasma membranes that extend out from the cell approximately µm, in order to incorporate the virus into the cell. The formation of macropinosomes occurs spontaneously, as a result of the activation of various growth factors, or simultaneously with the intake of cellular molecules or extracellular fluid. B. Clathrin-mediated endocytosis Clathrin-mediated endocytosis is the other means by which Ebolavirus enters the host cell. This process is very similar to macropinocytosis in that the plasma membrane forms invaginations that engulf the cell. However, clathrin-mediated endocytosis is different in that proteins on the surface of the host’s surface, and in particular clathrin, facilitate the attachment of the virus to the host’s cell surface. Glycoproteins are still used to attach the virus to the cell surface, and the NP-C1 cholesterol transporter still facilitates the fusion of the virus with endosomes and lysosomes and still allows the virus to escape into the cytoplasm. Without the NPC1 cholesterol transporter, Ebolavirus cannot leave the vesicle in order to replicate and cause infection in other cells. To penetrate the cell, the viral membrane fuses with vesicle membrane, and the nucleocapsid is released into the cytoplasm. In some culture cells, GP glycoprotein can be processed by host Cathepsin L andCathepsin B into 19kDa GP1. But this processing is not happening in all cells or for all ebolavirus. 19kDA GP1 interacts with host NPC1, which is highly expressed in dendritic cells. Fusion of virus membrane with the vesicle membrane is triggered by either low pH orNPC1 binding. 3. Sequential Transcription During transcription, the RNA genome is transcribed into seven monocistronic mRNAs whose length is determined by highly conserved start and stop signals. The transcription process begins with the binding of the polymerase complex to a single binding site located within the leader region of the genome. The complex then slides along the RNA template and sequentially transcribes the individual genes in their 3’ to 5’ order. Encapsidated, negative-sense genomic ssRNA is used as a template for the synthesis (3′-5′) of polyadenylated, monocistronic mRNAs and, using the host cell’s ribosomes, tRNA molecules, etc., the mRNA is translated into individual viral proteins. 4. Replication As viral protein levels rise, a switch occurs from translation to replication. Using the negative-sense genomic RNA as a template, a complementary +ssRNA is synthesized; this is then used as a template for the synthesis of new genomic (-)ssRNA, which is rapidly encapsidated Replication presumably starts when enough nucleoprotein is present to encapsidate neo-synthetized antigenomes and genomes. 5. Budding The newly formed nucleocapsids and envelope proteins associate at the host cell’s plasma membrane; budding occurs, destroying the cell. These viruses recruit components of the cellular ESCRT (endosomal sorting complex required for transport) system to mediate host-assisted viral budding. SCRT complexes are normally used by the cell for biological functions involving membrane remodeling, such as intraluminal vesicle formation, autophagy or terminal stages of cytokinesis. The ESCRT family consists of ESCRT-0, ESCRT-I, ESCRT-II which are primarily involved in cargo sorting and membrane deformation, and ESCRT-III which cleaves the bud neck from its cytosolic face . In the last step, vps4 disassembles the complex. The budding reaction catalyzed by the ESCRT machinery has reversed topology when compared with most other budding processes in the cell, such as endocytosis and formation of transport vesicles. 6. Release Finally, the virion is released.
 The virion enters early endosomes by Macropinocytosis or clathrin-mediated endocytosis. A. Macropinocytosis. In this process, ruffled segments of the host’s plasma membrane protrude outward from the cell and form invaginations where the virus utilizes glycoproteins in order to attach to the surface of the plasma membrane. Macropinocytosis is a process in which the Eukaryotic host cells form macropinosomes, segments of plasma membranes that extend out from the cell approximately µm, in order to incorporate the virus into the cell. The formation of macropinosomes occurs spontaneously, as a result of the activation of various growth factors, or simultaneously with the intake of cellular molecules or extracellular fluid. B. Clathrin-mediated endocytosis. Clathrin-mediated endocytosis is the other means by which Ebolavirus enters the host cell. This process is very similar to macropinocytosis in that the plasma membrane forms invaginations that engulf the cell. However, clathrin-mediated endocytosis is different in that proteins on the surface of the host’s surface, and in particular clathrin, facilitate the attachment of the virus to the host’s cell surface. Glycoproteins are still used to attach the virus to the cell surface, and the NP-C1 cholesterol transporter still facilitates the fusion of the virus with endosomes and lysosomes and still allows the virus to escape into the cytoplasm. Without the NPC1 cholesterol transporter, Ebolavirus cannot leave the vesicle in order to replicate and cause infection in other cells. To penetrate the cell, the viral membrane fuses with vesicle membrane, and the nucleocapsid is released into the cytoplasm. In some culture cells, GP glycoprotein can be processed by host Cathepsin L andCathepsin B into 19kDa GP1. But this processing is not happening in all cells or for all ebolavirus. 19kDA GP1 interacts with host NPC1, which is highly expressed in dendritic cells. Fusion of virus membrane with the vesicle membrane is triggered by either low pH orNPC1 binding. 3. Sequential Transcription. During transcription, the RNA genome is transcribed into seven monocistronic mRNAs whose length is determined by highly conserved start and stop signals. The transcription process begins with the binding of the polymerase complex to a single binding site located within the leader region of the genome. The complex then slides along the RNA template and sequentially transcribes the individual genes in their 3’ to 5’ order. Encapsidated, negative-sense genomic ssRNA is used as a template for the synthesis (3′-5′) of polyadenylated, monocistronic mRNAs and, using the host cell’s ribosomes, tRNA molecules, etc., the mRNA is translated into individual viral proteins. 4. Replication. As viral protein levels rise, a switch occurs from translation to replication. Using the negative-sense genomic RNA as a template, a complementary +ssRNA is synthesized; this is then used as a template for the synthesis of new genomic (-)ssRNA, which is rapidly encapsidated. Replication presumably starts when enough nucleoprotein is present to encapsidate neo-synthetized antigenomes and genomes. 5. Budding. The newly formed nucleocapsids and envelope proteins associate at the host cell’s plasma membrane; budding occurs, destroying the cell. These viruses recruit components of the cellular ESCRT (endosomal sorting complex required for transport) system to mediate host-assisted viral budding. SCRT complexes are normally used by the cell for biological functions involving membrane remodeling, such as intraluminal vesicle formation, autophagy or terminal stages of cytokinesis. The ESCRT family consists of ESCRT-0, ESCRT-I, ESCRT-II which are primarily involved in cargo sorting and membrane deformation, and ESCRT-III which cleaves the bud neck from its cytosolic face . In the last step, vps4 disassembles the complex. The budding reaction catalyzed by the ESCRT machinery has reversed topology when compared with most other budding processes in the cell, such as endocytosis and formation of transport vesicles. 6. Release. Finally, the virion is released.")
4
Mechanisms of induction of type I interferons by viruses
Viral nucleic acids and proteins are recognized by several cellular receptor families (TLRs and the family of cytosolic RIG-like receptors, or RLRs, which include MDA-5, RIG-I, DAI and others), which activate transcription factors (the IRF proteins) that stimulate the production of type I interferons, IFN-α and IFN-β. Cellular and Molecular Immunology, 7th ed., 2012 Elsevier
, which activate transcription factors (the IRF proteins) that stimulate the production of type I interferons, IFN-α and IFN-β. Cellular and Molecular Immunology, 7th ed., 2012 Elsevier.")
5
Major functions of the type I interferons
Interferon-α and interferon-β (IFN-α and IFN-β) have three major functions. First, they prevent viral replication by activating host genes that destroy viral mRNA and inhibit translation of viral proteins. Second, they increase the expression of ligands for NK-cell receptors. Third, they activate NK cells to kill virus-infected cells.
 have three major functions. First, they prevent viral replication by activating host genes that destroy viral mRNA and inhibit translation of viral proteins. Second, they increase the expression of ligands for NK-cell receptors. Third, they activate NK cells to kill virus-infected cells.")
6
Biologic actions of type I interferons
Type I interferons (IFN-α, IFN-β) are produced by virus-infected cells in response to intracellular TLR signaling and other sensors of viral RNA. Type I interferons bind to receptors on neighboring uninfected cells and activate JAKSTAT signaling pathways, which induce expression of genes whose products interfere with viral replication. Type I interferons also bind to receptors on infected cells and induce expression of genes whose products enhance the cell’s susceptibility to CTL-mediated killing. Cellular and Molecular Immunology, 7th ed., 2012 Elsevier
 are produced by virus-infected cells in response to intracellular TLR signaling and other sensors of viral RNA. Type I interferons bind to receptors on neighboring uninfected cells and activate JAKSTAT signaling pathways, which induce expression of genes whose products interfere with viral replication. Type I interferons also bind to receptors on infected cells and induce expression of genes whose products enhance the cell’s susceptibility to CTL-mediated killing. Cellular and Molecular Immunology, 7th ed., 2012 Elsevier.")
7
Activation of NK cells by receptors of innate and adaptive immunity
Left panel: activation of an NK cell by the 2B4 and NKG2D receptors of innate immunity. Both of these receptors must bind to their respective ligands on the target cell to produce signaling that will activate the NK cell. Right panel: an NK cell being activated by FcγRIIIA that has bound complexes of cell-surface antigens and antigen-specific IgG molecules. Signals from the Fc receptor alone are sufficient to activate the NK cell.

8
Antibody Dependent Cellular Cytotoxicity (ADCC)
When the antibodies bind to the antigen on virus infected cells, the FcγRIII receptors on the NK cell bind to the Fc regions of the cell-bound IgG. Multiple interactions between IgG and FcγRIII molecules establish stable binding between the NK cell and its target. Signals from FcγRIII activate the NK cell to form a conjugate pair and a synapse with the target cell. By secreting the contents of its lytic granules onto the surface of the target cell, the NK cell condemns the virus infected target cell to die by apoptosis.

9
Induction of T cell responses to virus-infected cells
Cells infected with intracellular microbes, such as viruses, are ingested by dendritic cells, and the antigens of the infectious microbes are processed and presented in association with class I MHC molecules to CD8+ T cells. Thus, dendritic cells are able to present endocytosed vesicular antigens by the class I pathway. Note that the same cross-presenting APCs may display class II MHC–associated antigens from the microbe for recognition by CD4+ helper T cells. Cellular and Molecular Immunology, 7th ed., 2012 Elsevier

10
Cytotoxic CD8 T cells can kill several infected target cells
in succession When a CD8 T cell recognizes a peptide:MHC class I complex on an infected cell (first panel), it programs the infected cell to die by apoptosis (second panel). The T cell then detaches from the first target cell, synthesizes a new set of lytic granules, and then seeks out a second target cell to attack (third panel). The cycle is repeated with an attack on a third target cell (fourth panel).
, it programs the infected cell to die by apoptosis (second panel). The T cell then detaches from the first target cell, synthesizes a new set of lytic granules, and then seeks out a second target cell to attack (third panel). The cycle is repeated with an attack on a third target cell (fourth panel).")
11
Viral infections are blocked by neutralizing antibodies
The effect of exposure to this year’s influenza virus is compared for a student who was vaccinated against the virus and made neutralizing anti-influenza IgA antibodies (left panels), and her identical twin sister at the same university who was too busy studying for a final exam to get vaccinated and lacks anti-influenza antibodies (right panels). To replicate itself, influenza must get inside human cells, which it does by using its hemagglutinin protein to bind to the sialic acid attached to human cell-surface proteins. Internalization of the virus, with subsequent fusion of the viral and endosomal membranes, releases the viral RNA into the cytoplasm, where replication occurs. This entire process can be halted at the very first step by the presence of neutralizing antibodies against the viral hemagglutinin that cover up its binding site for sialic acid. Because influenza infects epithelial cells of the respiratory tract, the effective antibodies are IgA dimers, as shown here.
, and her identical twin sister at the same university who was too busy studying for a final exam to get vaccinated and lacks anti-influenza antibodies (right panels). To replicate itself, influenza must get inside human cells, which it does by using its hemagglutinin protein to bind to the sialic acid attached to human cell-surface proteins. Internalization of the virus, with subsequent fusion of the viral and endosomal membranes, releases the viral RNA into the cytoplasm, where replication occurs. This entire process can be halted at the very first step by the presence of neutralizing antibodies against the viral hemagglutinin that cover up its binding site for sialic acid. Because influenza infects epithelial cells of the respiratory tract, the effective antibodies are IgA dimers, as shown here.")
12
How antibodies combat viruses
How antibodies combat viruses. (a) Neutralization of free virus by antibodies, (b) complement-mediated lysis of free virus and infected cells triggered by antibodies, (c) opsonization of virus particles by antibodies and phagocytosis of virus particles via Fc- or complement-receptors, (d) antibody-dependent cellular cytotoxicity (ADCC) against infected cells. Neutralizing antibodies (red), non-neutralizing (blue), Fc-receptors (violet), complement components (light-blue), complement-receptors (black).
 Neutralization of free virus by antibodies, (b) complement-mediated lysis of free virus and infected cells triggered by antibodies, (c) opsonization of virus particles by antibodies and phagocytosis of virus particles via Fc- or complement-receptors, (d) antibody-dependent cellular cytotoxicity (ADCC) against infected cells. Neutralizing antibodies (red), non-neutralizing (blue), Fc-receptors (violet), complement components (light-blue), complement-receptors (black).")
13
Kinetics of various anti-viral mechanisms
Kinetics of innate and adaptive immune responses to a virus infection. Cellular and Molecular Immunology, 7th ed., 2012 Elsevier

14
Innate and adaptive immune responses against viruses
Mechanisms by which innate and adaptive immunity prevent and eradicate virus infections. Innate immunity is mediated by type I interferons, which prevent infection, and NK cells, which eliminate infected cells. Adaptive immunity is mediated by antibodies and CTLs, which also block infection and kill infected cells, respectively. Cellular and Molecular Immunology, 7th ed., 2012 Elsevier

15
Evolution of new influenza variants by antigenic drift
Upon infection with influenza strain V, person P produced antibodies against various epitopes of the viral hemagglutinin. Some antibodies are neutralizing (green); others are not (blue). When person P is further exposed to strain V, the neutralizing antibodies prevent the virus from infecting cells (left panel). In the course of infecting person Q, viral strain V mutates to give strain V*, which differs from V by one amino acid substitution (yellow) in the hemagglutinin (center panel). This amino-acid difference eliminates the epitope recognized by neutralizing antibodies made against strain V. Consequently, strain V* influenza virus can infect cells of person P without interference from the antibodies made against strain V (right panel). To clear this second influenza infection, person P must mount a primary immune response that makes neutralizing antibodies against strain V*. The viral neuraminidase (not shown) undergoes antigenic drift in a similar manner.
; others are not (blue). When person P is further exposed to strain V, the neutralizing antibodies prevent the virus from infecting cells (left panel). In the course of infecting person Q, viral strain V mutates to give strain V*, which differs from V by one amino acid substitution (yellow) in the hemagglutinin (center panel). This amino-acid difference eliminates the epitope recognized by neutralizing antibodies made against strain V. Consequently, strain V* influenza virus can infect cells of person P without interference from the antibodies made against strain V (right panel). To clear this second influenza infection, person P must mount a primary immune response that makes neutralizing antibodies against strain V*. The viral neuraminidase (not shown) undergoes antigenic drift in a similar manner.")
16
Evolution of new influenza virus variants by antigenic shift
Human (red) and avian (blue) influenza viruses can simultaneously infect pigs, which in this context are called secondary hosts. In a pig cell that is infected by both viruses (left panel), their RNA segments become reassorted to produce a variety of recombinant viruses. One type of recombinant virus has a hemagglutinin of avian origin (center panel). The avian hemagglutinin is antigenically very different from the hemagglutinins of the influenza viruses currently infecting the human population, including the one that infected the pig cell. Individual humans are highly susceptible to the recombinant virus because they do not have antibodies that bind to the hemagglutinin and prevent the virus from infecting cells (right panel). Because the entire human population is vulnerable to the recombinant virus, the latter has the potential to produce a pandemic influenza.
 and avian (blue) influenza viruses can simultaneously infect pigs, which in this context are called secondary hosts. In a pig cell that is infected by both viruses (left panel), their RNA segments become reassorted to produce a variety of recombinant viruses. One type of recombinant virus has a hemagglutinin of avian origin (center panel). The avian hemagglutinin is antigenically very different from the hemagglutinins of the influenza viruses currently infecting the human population, including the one that infected the pig cell. Individual humans are highly susceptible to the recombinant virus because they do not have antibodies that bind to the hemagglutinin and prevent the virus from infecting cells (right panel). Because the entire human population is vulnerable to the recombinant virus, the latter has the potential to produce a pandemic influenza.")
17
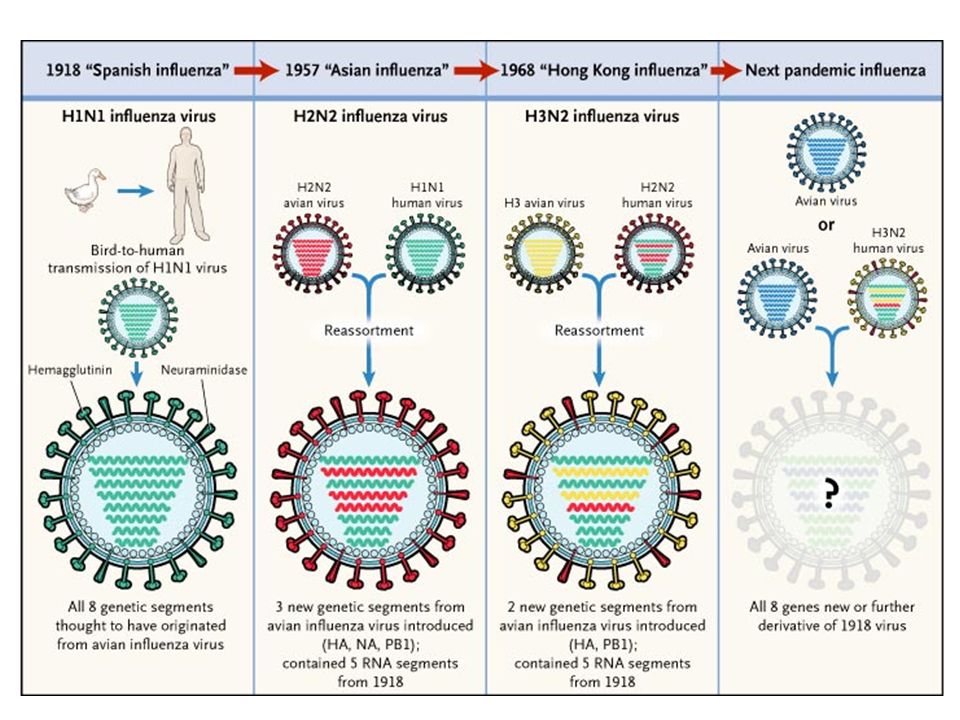
Generation of new influenza virus strains
by genetic recombination (antigenic shift) The genome of the influenza virus is composed of eight separate RNA strands, which allows genetic recombination by reassortment of the segments in various hosts, such as a pig, bird, or humans, that are simultaneously infected with two different strains. These genetic reassortments create new viruses that are antigenically distinct from their precursors and thus are able to evade immune detection in large numbers of newly infected hosts. In the example shown, H1N1 influenza virus, which was responsible for the pandemic of 2009, was generated by reassortment of swine, avian, and human viruses in pigs and then passed back to humans. Cellular and Molecular Immunology, 7th ed., 2012 Elsevier
 The genome of the influenza virus is composed of eight separate RNA strands, which allows genetic recombination by reassortment of the segments in various hosts, such as a pig, bird, or humans, that are simultaneously infected with two different strains. These genetic reassortments create new viruses that are antigenically distinct from their precursors and thus are able to evade immune detection in large numbers of newly infected hosts. In the example. shown, H1N1 influenza virus, which was responsible for the pandemic of 2009, was generated by reassortment of swine, avian, and human viruses in pigs and then passed back to humans. Cellular and Molecular Immunology, 7th ed., 2012 Elsevier.")
19
Mechanisms by which viruses inhibit antigen processing
and presentation The pathway of class I MHC–associated antigen presentation is shown, with examples of viruses that block different steps in this pathway. CMV, cytomegalovirus; EBV, Epstein-Barr virus; ER, endoplasmic reticulum; HSV, herpes simplex virus; TAP, transporter associated with antigen processing. Cellular and Molecular Immunology, 7th ed., 2012 Elsevier

21
The immunopathogenesis of HIV infection

24
The number of people living with HIV infection worldwide
is still increasing, but seems to be reaching a maximum

25
The virion of human immunodeficiency virus
The left panel is an electron micrograph showing three virions. The right panel is a diagram of a single virion. gp120 and gp41 are virally encoded envelope glycoproteins of molecular masses 120 kDa and 41 kDa that form the viral spike. Photograph courtesy of Hans Gelderblom.

26
The genes and proteins of HIV-1
HIV-1 has an RNA genome consisting of nine genes flanked by long terminal repeats (LTRs). The products of the nine genes and their known functions are tabulated. Several of the viral genes are overlapping and are read in different frames. Others encode large polyproteins that after translation are cleaved to produce several proteins having different activities. The gag, pol, and env genes are common to all retroviruses, and their protein products are all present in the virion.
. The products of the nine genes and their known functions are tabulated. Several of the viral genes are overlapping and are read in different frames. Others encode large polyproteins that after translation are cleaved to produce several proteins having different activities. The gag, pol, and env genes are common to all retroviruses, and their protein products are all present in the virion.")
27
The life cycle of HIV The gp120 envelope protein of the virus binds to CD4, enabling gp120 to also bind the chemokine co-receptor. This binding releases gp41, causing fusion of the viral envelope with the plasma membrane and release of the viral core into the cytoplasm. The RNA genome is released and reverse transcribed into double-stranded cDNA. This DNA migrates to the nucleus in association with the viral integrase, and becomes integrated into the cell genome, as a provirus (top panels). Activation of the T cell causes low-level transcription of the provirus that directs the synthesis of the early proteins Tat and Rev. These then expand and change the pattern of provirus transcription to produce mRNA encoding the protein constituents of the virion and RNA molecules corresponding to the HIV genome. Envelope proteins travel to the plasma membrane, whereas other viral proteins and viral genomic RNA assemble into nucleocapsids. New virus particles bud from the cell, acquiring their lipid envelope and envelope glycoproteins in the process (bottom panels).
. Activation of the T cell causes low-level transcription of the provirus that directs the synthesis of the early proteins Tat and Rev. These then expand and change the pattern of provirus transcription to produce mRNA encoding the protein constituents of the virion and RNA molecules corresponding to the HIV genome. Envelope proteins travel to the plasma membrane, whereas other viral proteins and viral genomic RNA assemble into nucleocapsids. New virus particles bud from the cell, acquiring their lipid envelope and envelope glycoproteins in the process (bottom panels).")
28
extinction of CD4 T cells
After infection with HIV there is a gradual extinction of CD4 T cells The number of CD4 T cells (green line) refers to those present in peripheral blood. Opportunistic infections and other symptoms become more frequent as the CD4 T-cell count falls, starting at around 500 cells/μl. The disease then enters the symptomatic phase. When CD4 T-cell counts fall below 200 cells/μl, the patient is said to have AIDS.
 refers to those present in peripheral blood. Opportunistic infections and other symptoms become more frequent as the CD4 T-cell count falls, starting at around 500 cells/μl. The disease then enters the symptomatic phase. When CD4 T-cell counts fall below 200 cells/μl, the patient is said to have AIDS.")
29
CD4+ T CELL DEPLETION IN HIV INFECTION
Direct cytopathic effect of HIV – lytic cycle in activated CD4+ T cells Killing by virus-specific CD8+ T lymphocytes – CD4+HIV+ targets Syncytia formation – gp120 of infected T cells binds to uninfected T cells fusion

30
The natural course of an HIV infection and the immune
response against it In the early phase of HIV infection, while the adaptive immune response is being activated, the virus reaches high levels (red line). With production of HIV-specific antibodies (blue lines) and cytotoxic T cells (yellow line). The virus is controlled but is not eliminated. When the destruction of CD4 T cells outstrips their renewal, adaptive immunity declines and levels of virus increase again.
. With production of HIV-specific antibodies (blue lines) and cytotoxic T cells (yellow line). The virus is controlled but is not eliminated. When the destruction of CD4 T cells outstrips their renewal, adaptive immunity declines and levels of virus increase again.")
31
Some opportunistic infections that kill patients with AIDS
Listed are the most common opportunistic infections that kill AIDS patients in developed countries. The malignancies are listed separately but they are also the result of impaired responses to infectious agents. Candida infection in oral cavity and esophagy

32
Receptors and co-receptors
All HIV strains can infect and replicate in freshly isolated human CD4+ T cells that are activated in vitro. In contrast, some strains will infect primary cultures of human macrophages but not continuous T cell lines (macrophagetropic, or M-tropic, virus), whereas other strains will infect T cell lines but not macrophages (T-tropic virus). Some virus strains also infect both T cell lines and macrophages (dual-tropic virus). Macrophage-tropic virus isolates express a gp120 that binds to CCR5, which is expressed on macrophages (and some memory T cells), whereas T cell–tropic viruses bind to CXCR4, which is expressed on T cell lines. HIV variants are described as X4 for CXCR4 binding, R5 for CCR5 binding, or R5X4 for the ability to bind to both chemokine receptors. In many HIV-infected individuals, there is a change from the production of virus that uses CCR5 and is predominantly macrophage tropic early in the disease to virus that binds to CXCR4 and is T cell line tropic late in the disease. The T-tropic strains tend to be more virulent, presumably because they infect and deplete T cells more than do M-tropic strains. CCR5 co-receptor on monocytes, macrophages and memory T cells CXCR4 co-receptor on activated T cells In initial infection In late infection
, whereas other strains will infect T cell lines but not macrophages (T-tropic virus). Some virus strains also infect both T cell lines and macrophages (dual-tropic virus). Macrophage-tropic virus isolates express a gp120 that binds to CCR5, which is expressed on macrophages (and some memory T cells), whereas T cell–tropic viruses bind to CXCR4, which is. expressed on T cell lines. HIV variants are described as X4 for CXCR4 binding, R5 for CCR5 binding, or R5X4 for the ability to bind to both chemokine receptors. In many HIV-infected individuals, there is a change from the production of virus that uses CCR5 and is predominantly macrophage tropic early in the disease to virus that binds to CXCR4 and is T cell line tropic late in the disease. The T-tropic strains tend to be more virulent, presumably because they infect and deplete T cells more than do M-tropic strains. CCR5 co-receptor on monocytes, macrophages and memory T cells. CXCR4 co-receptor on activated T cells. In initial infection. In late infection.")
33
of CCR5-Δ32 allele frequencies
Current world-wide frequency distribution of CCR5-Δ32 allele frequencies The delta 32 mutation is more prevalent in some racial backgrounds than in others. Population studies of the Caucasian population of western European ancestry revealed that approximately 1% of people were homozygous (2 copies) for the delta 32 mutation and up to 20% of people were heterozygous (1 copy). Within the United States, Canada and Australia, the frequency is 8% to 10% within the Caucasian background individuals, but less than 1% in the Afro-American populations. There is also a very low frequency of the mutation among the population in eastern Asia but much higher towards western Asia. 20% of the Caucasian population is heterozygous for a mutation in the CCR5 gene, which results in an unfunctional protein People homozygous for an inherited defect of the CCR5 gene are resistant to HIV infection – 1% of the Caucasian population
 for the delta 32 mutation and up to 20% of people were heterozygous (1 copy). Within the United States, Canada and Australia, the frequency is 8% to 10% within the Caucasian background individuals, but less than 1% in the Afro-American populations. There is also a very low frequency of the mutation among the population in eastern Asia but much higher towards western Asia. 20% of the Caucasian population is heterozygous for a mutation in the CCR5 gene, which results in an unfunctional protein. People homozygous for an inherited defect of the CCR5 gene are resistant to HIV infection – 1% of the Caucasian population.")
34
Three main transmission routes for HIV have been
identified Sexual The risk of female-to-male transmission is 0.04% per act and male-to-female transmission is 0.08% per act.

35
Blood products Blood transfusion, intravenous drug users, piercings, tattoos

36
Mother-to-child The risk of transmission is %. Perinatal > intrauterin > breast feeding

37
Antiretroviral therapy (ART) is the combination of several antiretroviral medicines used to slow the rate at which HIV makes copies of itself (multiplies) in the body. A combination of three or more antiretroviral medicines is more effective than using just one medicine (monotherapy) to treat HIV. The use of three or more antiretroviral medicines—sometimes referred to as an anti-HIV "cocktail"—is currently the standard treatment for HIV infection. So far, this treatment offers the best chance of preventing HIV from multiplying, which allows your immune system to stay healthy. The goal of antiretroviral therapy is to reduce the amount of virus in your body (viral load) to a level that can no longer be detected with current blood tests. Antiretroviral medicines that are often used to treat HIV include: Nucleoside/nucleotide reverse transcriptase inhibitors, also called nucleoside analogs, such as abacavir, emtricitabine, and tenofovir. These medicines are often combined for best results. Nonnucleoside reverse transcriptase inhibitors (NNRTIs), such as efavirenz, etravirine, and nevirapine. Protease inhibitors (PIs), such as atazanavir, darunavir, and ritonavir. Entry inhibitors, such as enfuvirtide and maraviroc. Integrase inhibitors, such as dolutegravir and raltegravir.
 to a level that can no longer be detected with current blood tests. Antiretroviral medicines that are often used to treat HIV include: Nucleoside/nucleotide reverse transcriptase inhibitors, also called nucleoside analogs, such as abacavir, emtricitabine, and tenofovir. These medicines are often combined for best results. Nonnucleoside reverse transcriptase inhibitors (NNRTIs), such as efavirenz, etravirine, and nevirapine. Protease inhibitors (PIs), such as atazanavir, darunavir, and ritonavir. Entry inhibitors, such as enfuvirtide and maraviroc. Integrase inhibitors, such as dolutegravir and raltegravir.")
38
Enzyme Mistakes The cause of HIV variability DNA-Polymerase
1 Mutation in bp Reverse Transcriptase 1 Mutation in bp THE VIRAL REVERSE TRANSCRIPTASE LACKS PROOF READING MECHANISMS OF TYPE POSSESSED BY CELLULAR DNA NEW VIRAL VARIANTS (quasi species) CO-EXIST IN AN INDIVIDUAL NEGATIVE SELECTION OF NEUTRALIZING AND CYTOTOXIC EPITOPES
 CO-EXIST IN AN INDIVIDUAL. NEGATIVE SELECTION OF NEUTRALIZING AND CYTOTOXIC EPITOPES.")
39
Antiviral drugs rapidly clear virus from the blood
and increase the number of circulating CD4 T cells The first and second panels show that maintenance of HIV levels in the blood depends on the continual infection of newly produced CD4 T cells. This is because cells live for only a few days once infected. The third and fourth panels show the effects of administering a drug (red squares) that blocks the viral life cycle. The existing virions in the blood are rapidly cleared by the actions of neutralizing antibody, complement, and phagocytes. Newly produced CD4 T cells are not infected, whereupon they live longer and accumulate in the circulation.
 that blocks the viral life cycle. The existing virions in the blood are rapidly cleared by the actions of neutralizing antibody, complement, and phagocytes. Newly produced CD4 T cells are not infected, whereupon they live longer and accumulate in the circulation.")
40
Broadly neutralizing antibodies recognize four epitopes
of the HIV envelope glycoprotein that trimerizes to form the viral spike Broadly neutralizing antibodies protect against many different strains of HIV. These antibodies interact with conserved parts of the envelope glycoprotein, which are important for its functional interactions. The orange-colored antibody binds to the CD4-binding site; the green-colored antibody binds to conserved elements of the V1 and V2 hypervariable loop of gp120; the purplecolored antibody binds to the conserved elements of the glycan shield and the V3 hypervariable loop of gp120; and the grey-colored antibody binds gp41. Image courtesy of Louise Scharf.

Similar presentations




>")


 –Phagocytic cells: WBC’s –Natural killer cells: perforins –Resident bacteria.>")

 2. Immunity to intracellular pathogens NK cells, control of Th1/Th2 responses 3.>")






 is the virus that causes Acquired Immunodeficiency Syndrome (AIDS).>")




 ◦ Pathogen: ◦ Reservoir: ◦ Portal of.>")

