Download presentation
Presentation is loading. Please wait.

1
Chapter 14: Signal-Transduction Pathways Copyright © 2007 by W. H. Freeman and Company Berg Tymoczko Stryer Biochemistry Sixth Edition

2
Signal Transduction Signal transduction is the transmission of a message across a cell membrane initiated by binding of a signal molecule to the outer surface of the cell. Agonist – molecule that binds to the receptor protein on outside of a cell. Receptor protein – a serpentine integral protein that communicates with transducer on the inside of the cell. The hydrophobic membrane spanning segments are highly conserved. The agonist induces a conformational change in the receptor protein as long as it is bound. An antagonist can bind and prevent agonist binding.

3
Signal Transduction Transducer – carries a message from the receptor protein to an effector protein on the inside of the cell. The transducer is activated by the change in conformation of that portion of the receptor protein protruding inside the cell and begins a second messenger system inside the cell. G-proteins are a family of transducers. Effector protein – produces a mobile second messenger such as cAMP. In the case of cAMP, the effector protein is adenylate cyclase (also called adenylyl cyclase).
..")
4
Signal Flow

5
Agonists and Effects

6
Second Messengers

8
Rhodopsin Example of a 7TM receptor protein. (7 transmembrane helices) G-proteins bind to 7TM receptors.
 G-proteins bind to 7TM receptors..")
9
G-Proteins G-proteins: These transducers are heterotrimeric proteins (three subunits).G = αβγ α = 45000 D; different for each G protein. The N-term of this subunit is attached to a myristoyl or palmitoyl group as a fatty acid anchor to the membrane. γ = 7-9000 D; several are known. The C-term of this subunit is attached to a farnesyl or geranyl group as a prenylated anchor to the membrane. β = 35000 D; several are known. β is associated strongly with γ. Note: α and βγ are anchored in the membrane.

10
GDPαβγ

11
G-Proteins Several different α, β and γ subunits have been identified. These form a family of G-proteins, such as: G s stimulates response in adenylyl cyclase G i inhibits response in adenylyl cyclase G t activates c-AMP phosphodiesterase (one in cones, and one in rods) G p or G q a phosphoinositide G-protein that activates phospholipase C G r transducin, transduces visual stimuli G o unknown, primary G-protein in brain
 G p or G q a phosphoinositide G-protein that activates phospholipase C G r transducin, transduces visual stimuli G o unknown, primary G-protein in brain.")
12
G-Protein Function G-protein forms a stable, inactive complex with GDP: GDPαβγ A specific GDPαβγ binds to a specific receptor protein at the inside cell face. A G-protein coupled receptor is a serpentine integral protein that has seven transmembrane segments (7TM proteins). The agonist-receptor protein complex induces a conformation change in αβγ and GDP dissociates from GDPαβγ, dissociation is normally slow. The agonist-receptor protein complex can also be considered to be a signal amplifier, activating 10- 1000 G-protein molecules depending upon the specific GDPαβγ.
. The agonist-receptor protein complex induces a conformation change in αβγ and GDP dissociates from GDPαβγ, dissociation is normally slow. The agonist-receptor protein complex can also be considered to be a signal amplifier, activating G-protein molecules depending upon the specific GDPαβγ..")
13
G-Protein Events Dissociation of GDP is slow: GDPαβγ ----> GDP + αβγ GTP replaces GDP in a fast step: GTP + αβγ ----> GTPαβγ GTP binding decreases the affinity of βγ to α: GTPαβγ ----> GTPα + βγ GTPα is active and free to move along the membrane with the anchor to an effector protein. Receptor proteins and effector proteins are separated in the membrane. GTPα binds to the effector protein and elicits a response (activation or inhibition). As long as GTPα is present, the response continues.
. As long as GTPα is present, the response continues..")
14
G-Protein Events GTPα is also a GTPase (it slowly hydrolyzes GTP to GDP and Pi) GTPα ----> GDPα + Pi GDPα is inactive and dissociates from effector protein. GDPα migrates back, binds to βγ and to the receptor protein:GDPαβγRP If agonist is still bound to the receptor protein, GDP dissociates, reactivation occurs, and another G-protein cycle begins.

15
G-Protein Events 1. When the agonist dissociates from the receptor protein, the transduction process stops. This will depend in part on production of the hormone and its concentration. 2. Phosphorylation of Ser and Thr residues at the C-term (on the cytosolic face) of the receptor protein can also arrest the process. G-protein receptor kinase 2 (GRK-2) is responsible and attacks only the agonistRP and not the unbound receptor protein. The protein β-arrestin binds to agonist RP-P 2 to prevent G-protein action.
 of the receptor protein can also arrest the process. G-protein receptor kinase 2 (GRK-2) is responsible and attacks only the agonistRP and not the unbound receptor protein. The protein β-arrestin binds to agonist RP-P 2 to prevent G-protein action..")
16
G-protein switch At Receptor Protein At Adenylate cyclase

17
Adenylate Cyclase Effector Adenylyl cyclase (effector protein) is activated by GTPα and catalyzes formation of c-AMP. ATP ----> 3',5'-c-AMP + PPi Pyrophosphatase hydrolyzes PPi and pulls the reaction to completion.PPi ----> 2 Pi c-AMP is a second messenger that activates c-AMP dependent protein kinase (Protein Kinase A). Phosphodiesterase hydrolyzes c-AMP to 5'-AMP (destroys the second messenger). AMP has no second messenger capabilities.
. Phosphodiesterase hydrolyzes c-AMP to 5 -AMP (destroys the second messenger). AMP has no second messenger capabilities..")
18
An Agonist

19
12 Transmembrane Helices

20
G-Protein function

21
GTP hydrolysis & reform GDPαβγ

22
Deactivation of RP

23
Turning-on G-Protein Note: several types of hormone receptors may turn-on the same G-protein. For instance, in liver, there is a receptor site for glucagon (a polypeptide hormone) and another for epinephrine (a β-andrenergic or catecholamine hormone) these both activate G s, which stimulates adenylyl cyclase. A different receptor protein and agonist may turn-on another G-protein, G i, which inhibits adenylyl cyclase.
 and another for epinephrine (a β-andrenergic or catecholamine hormone) these both activate G s, which stimulates adenylyl cyclase. A different receptor protein and agonist may turn-on another G-protein, G i, which inhibits adenylyl cyclase..")
24
Flow diagram for G-protein mediated transduction

25
G-Protein System

26
P-Inositol-bisP and Diacylglycerol Phosphorylation of phosphatidyl inositol (cytosolic face) is catalyzed by PI kinase giving PI-4-P (PIP), followed a second time by PIP kinase to form PI-4,5-bisP (PIP 2 ). PI + ATP —> PI-4-P + ADP PI kinase PI-4-P + ATP —> PI-4,5-bisP + ADP PIP kinase The agonist, angiotensin II, binding to an ATII receptor initiates the G-protein process with G q. This also occurs with epinephrine and an α 1 andrenergic receptor.

27
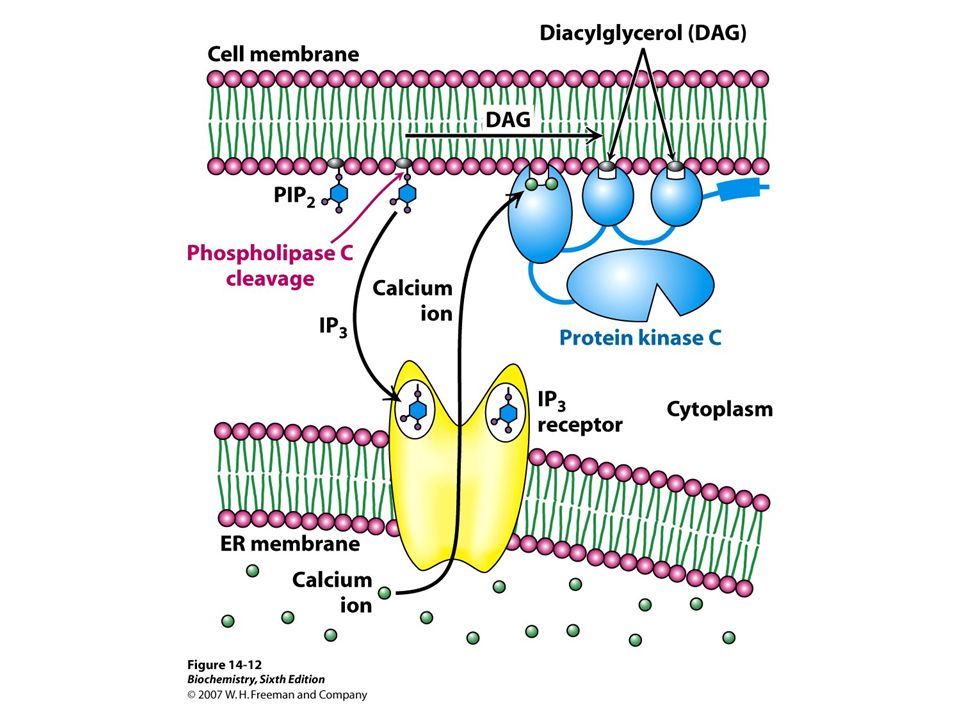
PIP 2 and DAG G q (GTPα q ) activates phospholipase C-β which cleaves PIP 2 to diacylglycerol (DAG) and inositol- 1,4,5-triphosphate both of which are second messengers. PIP 2 —> DAG + IP 3 DAG = 1-steroyl-2-arachidonyl-sn-glycerol The non-polar DAG remains membrane bound but moves and activates the effector protein kinase C. Protein kinase C phosphorylates Ser or Thr in a number of regulatory proteins and requires that DAG bind Ca ++ for activity.

28
PIP 2 and DAG

29
DAG is degraded in some cells to provide arachidonic acid for eicosanoid synthesis. The polar IP 3 diffuses into the cytosol and stimulates Ca ++ release from the EPR through a calcium channel by binding to an IP3 receptor. Ca ++ then activates calcium dependent protein kinase and the Ca ++ ATPase acting through calmodulin. Ca ++ serves other functions as well. Activation of effector ceases when GTPα q hydrolyzes to GDPα q.

31
PIP 2 and DAG IP 3 and DAG are rapidly recycled to PIP 2, not directly, but through a sequence of metabolic reactions. Phospholipase C-γ is activated through the inositol phosphate pathway via tyrosine kinase receptors (RTKs).Norepinephrine, vasopressin and acetylcholine have receptors that lead to the inositol-P signaling pathway. Calmodulin (17 kD) has 4 Ca ++ binding sites. It binds well when [Ca] > 500 mM; the normal steady state cytosolic [Ca] = 100 mM. Cells are sensitive to changes in low [Ca]. Ca ++ binding to calmodulin is in an EF Hand (helix-loop-helix) structure.
.Norepinephrine, vasopressin and acetylcholine have receptors that lead to the inositol-P signaling pathway. Calmodulin (17 kD) has 4 Ca ++ binding sites. It binds well when [Ca] > 500 mM; the normal steady state cytosolic [Ca] = 100 mM. Cells are sensitive to changes in low [Ca]. Ca ++ binding to calmodulin is in an EF Hand (helix-loop-helix) structure..")
33
Ca ++ Spreading in a cell

34
Ca ++ binding in Calmodulin Ca ++ binding. Six coordination sites with the protein and one with water.

37
Inositol-P System

38
Tyrosine Kinases (TK or RTK) Tyrosine kinase is a receptor protein that exists as β transmembrane chain attached through a disulfide bond to an α chain on the outside of the membrane. Each αβ complex is referred to as a monomeric subunit. Binding of insulin to the RP promotes dimerization of the αβ subunits and TK becomes active. One insulin molecule is bound between the two subunits.

39
Insulin Structure

40
Receptor Tyrosine Kinase

41
Tyrosine Kinases Active TK subunits phosphorylate each other at Tyr in both β chains on the cytosolic side of the αβαβ dimer. This process uses ATP and is known as autophosphorylation. Autophosphorylation induces a conformation change, activating the kinase form of TK. TK can now phosphorylate certain cytosolic proteins at tyrosine. So, this receptor protein is also a kinase (an effector protein).
..")
42
Autophosphorylation

43
TK Phosphorylation

44
Tyrosine Kinases TK acts on a group of small proteins called insulin receptor substrates (IRS) that are phosphorylated at Tyr in 4 Tyr-X-X-Met sequences. IRS then coordinates association among TK, PIP 2 and phosphoinositide-3 kinase (PI-3K). PI-3K has a SH2 domain used to bind Tyr-P. PI 3-kinase uses PIP 2 as substrate and phosphorylates the 3 position of inositol. PIP 2 + ATP ----> PI-3,4,5-trisP (PIP 3 ) + ADP
. PI-3K has a SH2 domain used to bind Tyr-P. PI 3-kinase uses PIP 2 as substrate and phosphorylates the 3 position of inositol. PIP 2 + ATP ----> PI-3,4,5-trisP (PIP 3 ) + ADP.")
46
PIP 3

47
Tyrosine Kinase System

48
Tyrosine Kinases PIP 3 is a membrane bound second messenger that activates certain protein kinases, such as 3-phosphoinositide-dependent protein kinase 1 (PDK1) which is a membrane bound Ser/Thr kinase. PDK-1 phosphorylates other mobile kinases in the cytosol such as Akt (protein kinase B) which is also a Ser/Thr kinase.
 which is also a Ser/Thr kinase..")
49
Tyrosine Kinases Insulin signaling is terminated by dissociation of insulin from the receptor. The activation process that has been initiated is deactivated via dephosphorylation of the various sites to which phosphate has been attached. Several types of phosphatases are important here: Protein tyrosine phosphatase for Tyr-P, Lipid phosphatases for PIP 3 and Protein serine phosphatases for Ser-P / Thr-P.

50
Tyrosine Kinase System

52
End of Chapter 14 Copyright © 2007 by W. H. Freeman and Company Berg Tymoczko Stryer Biochemistry Sixth Edition

Similar presentations




















 Membranes. Membrane transport Membranes are selectively permeable barriers Hydrophobic uncharged small molecules can freely diffuse.>")




