Download presentation
Presentation is loading. Please wait.

1
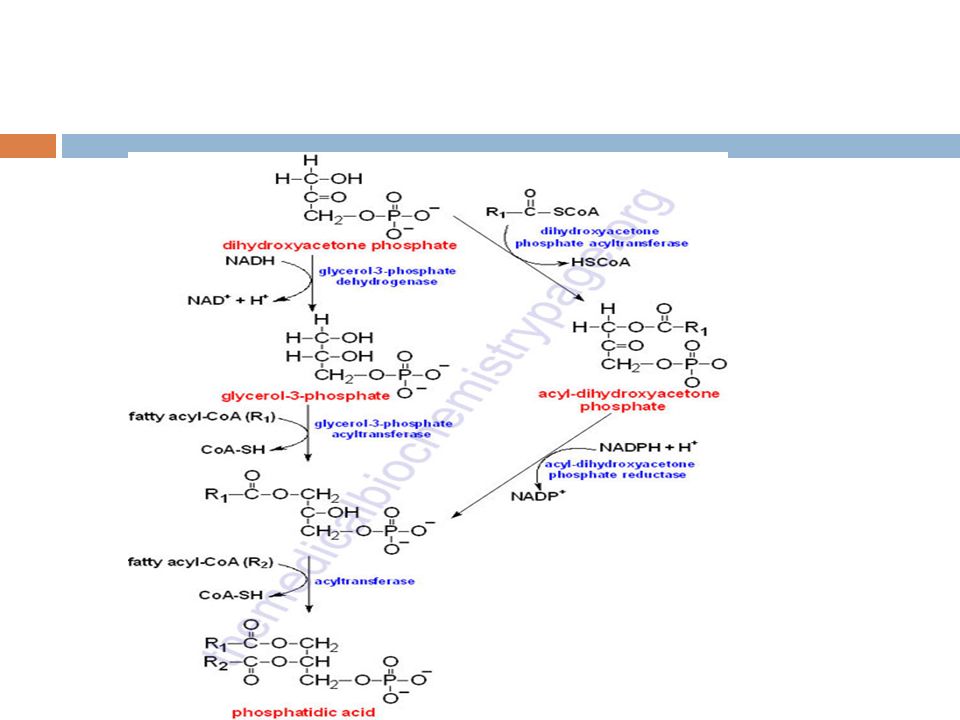
Lipidosis Prof. S. Kajuna Synthesis of Triglycerides Fatty acids are stored for future use as triacylglycerols (TAGs) in all cells, but primarily in adipocytes of adipose tissue. TAGs constitute molecules of glycerol to which three fatty acids have been esterified. The fatty acids present in TAGs are predominantly saturated. The major building block for the synthesis of TAGs, in tissues other than adipose tissue, is glycerol. Adipocytes lack glycerol kinase, therefore, dihydroxyacetone phosphate (DHAP), produced during glycolysis, is the precursor for TAG synthesis in adipose tissue. This means that adipocytes must have glucose to oxidize in order to store fatty acids in the form of TAGs. DHAP can also serve as a backbone precursor for TAG synthesis in tissues other than adipose, but does so to a much lesser extent than glycerol.
 in all cells, but primarily in adipocytes of adipose tissue. TAGs constitute molecules of glycerol to which three fatty acids have been esterified. The fatty acids present in TAGs are predominantly saturated. The major building block for the synthesis of TAGs, in tissues other than adipose tissue, is glycerol. Adipocytes lack glycerol kinase, therefore, dihydroxyacetone phosphate (DHAP), produced during glycolysis, is the precursor for TAG synthesis in adipose tissue. This means that adipocytes must have glucose to oxidize in order to store fatty acids in the form of TAGs. DHAP can also serve as a backbone precursor for TAG synthesis in tissues other than adipose, but does so to a much lesser extent than glycerol..")
4
The glycerol backbone of TAGs is activated by phosphorylation at the C-3 position by glycerol kinase. The utilization of DHAP for the backbone is carried out through either of two pathways depending upon whether the synthesis of triglycerides is carried out in the mitochondria and ER or the ER and the peroxisomes. In the former case the action of glycerol-3-phosphate dehydrogenase, a reaction that requires NADH (the same reaction as that used in the glycerol-phosphate shuttle), converts DHAP to glycerol-3-phosphate.
, converts DHAP to glycerol-3-phosphate..")
5
Glycerol-3-phosphate acyltransferase (GPAT) then esterifies a fatty acid to glycerol-3-phosphate generating the monoacylglycerol phosphate structure called lysophosphatidic acid. The second reaction pathway utilizes the peroxisomal enzyme DHAP acyltransferase to fatty acylate DHAP to acyl-DHAP which is then reduced by the NADPH-requiring enzyme acyl-DHAP reductase.

6
The fatty acids incorporated into TAGs are activated to acyl-CoAs through the action of acyl-CoA synthetases. Two molecules of acyl-CoA are esterified to glycerol-3- phosphate to yield 1,2-diacylglycerol phosphate (commonly identified as phosphatidic acid). The phosphate is then removed, by phosphatidic acid phosphatase (PAP1), to yield 1,2-diacylglycerol, the substrate for addition of the third fatty acid. Intestinal monoacylglycerols, derived from the hydrolysis of dietary fats, can also serve as substrates for the synthesis of 1,2-diacylglycerols.
. The phosphate is then removed, by phosphatidic acid phosphatase (PAP1), to yield 1,2-diacylglycerol, the substrate for addition of the third fatty acid. Intestinal monoacylglycerols, derived from the hydrolysis of dietary fats, can also serve as substrates for the synthesis of 1,2-diacylglycerols..")
7
Phospholipid synthesis acid (1,2-diacylglycerol 3-phosphate). Most phospholipids have a saturated fatty acid on C-1 and an unsaturated fatty acid on C-2 of the glycerol backbone. The most commonly added alcohols (serine, ethanolamine and choline) also contain nitrogen that may be positively charged, whereas, glycerol and inositol do not.
 also contain nitrogen that may be positively charged, whereas, glycerol and inositol do not..")
8
The major classifications of phospholipids are:Phosphatidylcholine (PC)
")
9
Phosphatidylethanolamine (PE)
")
10
Phosphatidylserine (PS)
")
11
Phosphatidylinositol (PI)
")
12
Phosphatidylglycerol (PG)
")
13
Cardiolipin (diphosphatidylglycerol, DPG)
")
14
PC Phospholipids can be synthesized by two mechanisms. One utilizes a CDP-activated polar head group for attachment to the phosphate of phosphatidic acid. The other utilizes CDP-activated 1,2-diacylglycerol and an inactivated polar head group. This class of phospholipids is also called the lecithins. At physiological pH, phosphatidylcholines are neutral zwitterions. They contain primarily palmitic or stearic acid at carbon 1 and primarily oleic, linoleic or linolenic acid at carbon 2. The lecithin dipalmitoyllecithin is a component of lung or pulmonary surfactant.

15
It contains palmitate at both carbon 1 and 2 of glycerol and is the major (80%) phospholipid found in the extracellular lipid layer lining the pulmonary alveoli. Choline is activated first by phosphorylation and then by coupling to CDP prior to attachment to phosphatidic acid. PC is also synthesized by the addition of choline to CDP-activated 1,2-diacylglycerol. A third pathway to PC synthesis, involves the conversion of either PS or PE to PC. The conversion of PS to PC first requires decarboxylation of PS to yield PE; this then undergoes a series of three methylation reactions utilizing S- adenosylmethionine (SAM) as methyl group donor.
 as methyl group donor..")
16
PE These molecules are neutral zwitterions at physiological pH. They contain primarily palmitic or stearic acid on carbon 1 and a long chain unsaturated fatty acid (e.g. 18:2, 20:4 and 22:6) on carbon 2. Synthesis of PE can occur by two pathways. The first requires that ethanolamine be activated by phosphorylation and then by coupling to CDP. The ethanolamine is then transferred from CDP-ethanolamine to phosphatidic acid to yield PE. The second involves the decarboxylation of PS.
 on carbon 2. Synthesis of PE can occur by two pathways. The first requires that ethanolamine be activated by phosphorylation and then by coupling to CDP. The ethanolamine is then transferred from CDP-ethanolamine to phosphatidic acid to yield PE. The second involves the decarboxylation of PS..")
17
PS Phosphatidylserines will carry a net charge of –1 at physiological pH and are composed of fatty acids similar to the phosphatidylethanolamines. The pathway for PS synthesis involves an exchange reaction of serine for ethanolamine in PE. This exchange occurs when PE is in the lipid bilayer of the a membrane. As indicated above, PS can serve as a source of PE through a decarboxylation reaction.

18
PI: These molecules contain almost exclusively stearic acid at carbon 1 and arachidonic acid at carbon 2. Phosphatidylinositols composed exclusively of non- phosphorylated inositol exhibit a net charge of –1 at physiological pH. These molecules exist in membranes with various levels of phosphate esterified to the hydroxyls of the inositol. Molecules with phosphorylated inositol are termed polyphosphoinositides. The polyphosphoinositides are important intracellular transducers of signals emanating from the plasma membrane. The synthesis of PI involves CDP-activated 1,2-diacylglycerol condensation with myo-inositol

19
PI subsequently undergoes a series of phosphorylations of the hydroxyls of inositol leading to the production of polyphosphoinositides. One polyphosphoinositide (phosphatidylinositol 4,5- bisphosphate, PIP 2 ) is a critically important membrane phospholipid involved in the transmission of signals for cell growth and differentiation from outside the cell to inside.
 is a critically important membrane phospholipid involved in the transmission of signals for cell growth and differentiation from outside the cell to inside..")
20
PG: Phosphatidylglycerols exhibit a net charge of –1 at physiological pH. These molecules are found in high concentration in mitochondrial membranes and as components of pulmonary surfactant. Phosphatidylglycerol also is a precursor for the synthesis of cardiolipin. PG is synthesized from CDP- diacylglycerol and glycerol-3-phosphate. The vital role of PG is to serve as the precursor for the synthesis of diphosphatidylglycerols (DPGs).
..")
21
DPG These molecules are very acidic, exhibiting a net charge of –2 at physiological pH. They are found primarily in the inner mitochondrial membrane and also as components of pulmonary surfactant. One important class of diphosphatidylglycerols is the cardiolipins. These molecules are synthesized by the condensation of CDP-diacylglycerol with PG.

22
The fatty acid distribution at the C–1 and C–2 positions of glycerol within phospholipids is continually in flux, owing to phospholipid degradation and the continuous phospholipid remodeling that occurs while these molecules are in membranes. Phospholipid degradation results from the action of phospholipases. There are various phospholipases that exhibit substrate specificities for different positions in phospholipids.

23
In many cases the acyl group which was initially transferred to glycerol, by the action of the acyl transferases, is not the same acyl group present in the phospholipid when it resides within a membrane. The remodeling of acyl groups in phospholipids is the result of the action of phospholipase A 1 (PLA 1 ) and phospholipase A 2 (PLA 2).
 and phospholipase A 2 (PLA 2)..")
24
Sites of action of the phospholipases A 1, A 2, C and D.

25
The products of these phospholipases are called lysophospholipids and can be substrates for acyl transferases utilizing different acyl-CoA groups. Lysophospholipids can also accept acyl groups from other phospholipids in an exchange reaction catalyzed by lysolecithin:lecithin acyltransferase (LLAT). PLA 2 is also an important enzyme, whose activity is responsible for the release of arachidonic acid from the C–2 position of membrane phospholipids. The released arachidonate is then a substrate for the synthesis of the eicosanoids.
. PLA 2 is also an important enzyme, whose activity is responsible for the release of arachidonic acid from the C–2 position of membrane phospholipids. The released arachidonate is then a substrate for the synthesis of the eicosanoids..")
26
Plasmalogens Plasmalogens are glycerol ether phospholipids. They are of two types, alkyl ether (–O–CH 2 –) and alkenyl ether (–O–CH=CH–). Dihydroxyacetone phosphate serves as the glycerol precursor for the synthesis of glycerol ether phospholipids. Three major classes of plasmalogens have been identified: choline, ethanolamine and serine plasmalogens. Ethanolamine plasmalogen is prevalent in myelin. Choline plasmalogen is abundant in cardiac tissue.
 and alkenyl ether (–O–CH=CH–). Dihydroxyacetone phosphate serves as the glycerol precursor for the synthesis of glycerol ether phospholipids. Three major classes of plasmalogens have been identified: choline, ethanolamine and serine plasmalogens. Ethanolamine plasmalogen is prevalent in myelin. Choline plasmalogen is abundant in cardiac tissue..")
27
One particular choline plasmalogen (1-O-1'-enyl-2- acetyl-sn-glycero-3-phosphocholine) has been identified as an extremely powerful biological mediator, capable of inducing cellular responses at concentrations as low as 10 –11 M. This molecule is called platelet activating factor, PAF.

28
PAF functions as a mediator of hypersensitivity, acute inflammatory reactions and anaphylactic shock. PAF is synthesized in response to the formation of antigen-IgE complexes on the surfaces of basophils, neutrophils, eosinophils, macrophages and monocytes. The synthesis and release of PAF from cells leads to platelet aggregation and the release of serotonin from platelets. PAF also produces responses in liver, heart, smooth muscle, and uterine and lung tissues.

29
Platelet activating factor

30
Sphingolipids Sphingolipids, or glycosylceramides, are a class of lipids containing a backbone of sphingoid bases, a set of aliphatic amino alcohols that includes sphingosine. lipidsaliphaticaminoalcoholssphingosine A sphingolipid with an R group consisting of a hydrogen atom only is a ceramide. Other common R groups include phosphocholine, yielding a sphingomyelin, and various sugar monomers or dimers, yielding cerebrosides and globosides, respectively. Cerebrosides and globosides are collectively known as glycosphingolipids.ceramidephosphocholinesphingomyelin cerebrosidesglobosides glycosphingolipids

31
General structures of sphingolipids

32
The long-chain bases, sometimes simply known as sphingoid bases, are the first non-transient products of de novo sphingolipid synthesis in both yeast and mammals. These compounds, specifically known as phytosphingosine and dihydrosphingosine (also known as sphinganine, although this term is less common), are mainly C 18 compounds, with somewhat lower levels of C 20 bases. Ceramides and glycosphingolipids are N-acyl derivatives of these compounds.de novo The sphingosine backbone is O-linked to a (usually) charged head group such as ethanolamine, serine, or choline.ethanolamineserinecholine The backbone is also amide-linked to an acyl group, such as a fatty acid.acyl groupfatty acid
, are mainly C 18 compounds, with somewhat lower levels of C 20 bases. Ceramides and glycosphingolipids are N-acyl derivatives of these compounds.de novo The sphingosine backbone is O-linked to a (usually) charged head group such as ethanolamine, serine, or choline.ethanolamineserinecholine The backbone is also amide-linked to an acyl group, such as a fatty acid.acyl groupfatty acid.")
33
Simple sphingolipids, which include the sphingoid bases and ceramides, make up the early products of the sphingolipid synthetic pathways. Ceramides, as a general class, are N-acylated sphingoid bases lacking additional head groups. Ceramides Dihydroceramide is produced by N-acylation of dihydrosphingosine. Dihydroceramide is found in both yeast and mammalian systems. Ceramide is produced in mammalian systems by desaturation of dihydroceramide by dihydroceramide desaturase 1 (DES1). This highly bioactive molecule may also be phosphorylated to form ceramide-1-phosphate. Ceramideceramide-1-phosphate
. This highly bioactive molecule may also be phosphorylated to form ceramide-1-phosphate. Ceramideceramide-1-phosphate.")
34
Complex sphingolipids may be formed by addition of head groups to ceramide or phytoceramide: Sphingomyelins have a phosphocholine or phosphoethanolamine molecule with an ester linkage to the 1-hydroxy group of a ceramide. Sphingomyelinsphosphocholinephosphoethanolamineester linkage Glycosphingolipids are ceramides with one or more sugar residues joined in a β -glycosidic linkage at the 1-hydroxyl position (see image). Glycosphingolipidssugarglycosidic Cerebrosides have a single glucose or galactose at the 1-hydroxy position. Cerebrosidesglucosegalactose Sulfatides are sulfated cerebrosides. Sulfatides Gangliosides have at least three sugars, one of which must be sialic acid. Gangliosidessialic acid Inositol-containing ceramides, which are derived from phytoceramide, are produced in yeast. These include inositol phosphorylceramide, mannose inositol phosphorylceramide, and mannose diinositol phosphorylceramide. Inositolmannose
. Glycosphingolipidssugarglycosidic Cerebrosides have a single glucose or galactose at the 1-hydroxy position. Cerebrosidesglucosegalactose Sulfatides are sulfated cerebrosides. Sulfatides Gangliosides have at least three sugars, one of which must be sialic acid. Gangliosidessialic acid Inositol-containing ceramides, which are derived from phytoceramide, are produced in yeast. These include inositol phosphorylceramide, mannose inositol phosphorylceramide, and mannose diinositol phosphorylceramide. Inositolmannose.")
35
Mammalian sphingolipid metabolism De novo sphingolipid synthesis begins with formation of 3-keto-dihydrosphingosine by serine palmitoyltransferase. The preferred substrates for this reaction are palmitoyl-CoA and serine. However, studies have demonstrated that serine palmitoyltransferase has some activity toward other species of fatty acyl-CoA and alternative amino acids.serine palmitoyltransferasepalmitoyl-CoAserineamino acids

36
Ceramide may subsequently have several fates. It may be phosphorylated by ceramide kinase to form ceramide-1-phosphate. Alternatively, it may be glycosylated by glucosylceramide synthase or galactosylceramide synthase. Additionally, it can be converted to sphingomyelin by the addition of a phosphorylcholine headgroup by sphingomyelin synthase. Diacylglycerol is generated by this process. Finally, ceramide may be broken down by a ceramidase to form sphingosine. Sphingosine may be phosphorylated to form sphingosine-1-phosphate. This may be dephosphorylated to reform sphingosine. [ Ceramideceramide kinaseglucosylceramide synthase galactosylceramide synthasesphingomyelin phosphorylcholinesphingomyelin synthaseDiacylglycerol ceramidasesphingosine [

37
Functions of mammalian sphingolipids Sphingolipids are commonly believed to protect the cell surface against harmful environmental factors by forming a mechanically stable and chemically resistant outer leaflet of the plasma membrane lipid bilayer. Certain complex glycosphingolipids were found to be involved in specific functions, such as cell recognition and signaling. Cell recognition depends mainly on the physical properties of the sphingolipids, whereas signaling involves specific interactions of the glycan structures of glycosphingolipids with similar lipids present on neighboring cells or with proteins.plasma membranelipid bilayerglycosphingolipidscell recognition and signalingproteins

38
Recently, simple sphingolipid metabolites, such as ceramide and sphingosine-1-phosphate, have been shown to be important mediators in the signaling cascades involved in apoptosis, proliferation, stress responses, necrosis, inflammation, autophagy, senescence, and differentiation. Ceramide-based lipids self-aggregate in cell membranes and form separate phases less fluid than the bulk phospholipids. These sphingolipid-based microdomains, or "lipid rafts" were originally proposed to sort membrane proteins along the cellular pathways of membrane transport. At present, most research focuses on the organizing function during signal transduction.metabolitessphingosine-1-phosphateapoptosisproliferationnecrosisinflammationautophagy senescencedifferentiationcell membranes phaseslipid rafts

39
Sphingolipids are synthesized in a pathway that begins in the ER and is completed in the Golgi apparatus, but these lipids are enriched in the plasma membrane and in endosomes, where they perform many of their functions. Transport occurs via vesicles and monomeric transport in the cytosol. Sphingolipids are virtually absent from mitochondria and the ER, but constitute a 20-35 molar fraction of plasma membrane lipids.ERGolgi apparatusplasma membraneendosomescytosolmitochondriaER In experimental animals, feeding sphingolipids inhibits colon carcinogenesis, reduces (bad) LDL cholesterol and elevates (good) HDL cholesterol. colon carcinogenesisLDL cholesterolHDL cholesterol
 LDL cholesterol and elevates (good) HDL cholesterol. colon carcinogenesisLDL cholesterolHDL cholesterol.")
40
Sphingolipidoses There are several disorders of sphingolipid metabolism, known as sphingolipidoses. The main members of this group are Niemann-Pick disease, Fabry disease, Krabbe disease, Gaucher disease, Tay- Sachs disease and Metachromatic leukodystrophy. They are generally inherited in an autosomal recessive fashion, but notably Fabry disease is X-linked. Taken together, sphingolipidoses have an incidence of approximately 1 in 10.000, but substantially more in certain populations such as Ashkenazi Jews. Enzyme replacement therapy is available to treat mainly Fabry disease and Gaucher disease, and people with these types of sphingolipidoses may live well into adulthood. The other types are generally fatal by age 1 to 5 years for infantile forms, but progression may be mild for juvenile- or adult-onset forms. sphingolipidosesNiemann-Pick diseaseFabry diseaseKrabbe diseaseGaucher diseaseTay- Sachs diseaseMetachromatic leukodystrophyautosomal recessive Fabry diseaseX-linked incidenceAshkenazi JewsEnzyme replacement therapyFabry diseaseGaucher disease

Similar presentations

















 –But typically amphipathic Lipophilic (hydrophobic) chain Polar/charged (hydrophilic)>")


