Download presentation
Presentation is loading. Please wait.

1
Using the genome Studying expression of all genes simultaneously 1.Microarrays: “reverse Northerns” 2.High-throughput sequencing 3. Bisulfite sequencing to detect C methylation

2
Using the genome Bisulfite sequencing to detect C methylation ChIP-chip or ChIP-seq to detect chromatin modifications: 17 mods are associated with active genes in CD-4 T cells

3
Generating the histone code Histone acetyltransferases add acetic acid Deacetylases “reset” by removing the acetate

4
Generating the histone code CDK8 kinases histones to repress transcription Appears to interact with mediator to block transcription Phosphorylation of Histone H3 correlates with activation of heat shock genes! Phosphatases reset the genes

5
Generating the histone code Rad6 proteins ubiquitinate histone H2B to repress transcription Polycomb proteins ubiquitinate histone H2A to silence genes

6
Generating the histone code Rad6 proteins ubiquitinate histone H2B to repress transcription Polycomb proteins ubiquitinate histone H2A to silence genes A TFTC/STAGA module mediates histone H2A and H2B deubiquitination, coactivates nuclear receptors, and counteracts heterochromatin silencing

7
Generating the histone code Many proteins methylate histones: highly regulated!

8
Generating the histone code Many proteins methylate histones: highly regulated! Methylation status determines gene activity

9
Generating the histone code Many proteins methylate histones: highly regulated! Methylation status determines gene activity Mutants (eg Curly leaf) are unhappy!
 are unhappy!.")
10
Generating the histone code Many proteins methylate histones: highly regulated! Methylation status determines gene activity Mutants (eg Curly leaf) are unhappy! Chromodomain protein HP1 can tell the difference between H3K9me2 (yellow) & H3K9me3 (red)
 are unhappy. Chromodomain protein HP1 can tell the difference between H3K9me2 (yellow) & H3K9me3 (red).")
11
Generating the histone code Chromodomain protein HP1 can tell the difference between H3K9me2 (yellow) & H3K9me3 (red) Histone demethylases have been recently discovered
 & H3K9me3 (red) Histone demethylases have been recently discovered")
12
Generating methylated DNA Si RNA are key: RNA Pol IV generates antisense or foldback RNA, often from TE

13
Generating methylated DNA Si RNA are key: RNA Pol IV generates antisense or foldback RNA, often from TE RDR2 makes it DS, 24 nt siRNA are generated by DCL3

14
Generating methylated DNA RDR2 makes it DS, 24 nt siRNA are generated by DCL3 AGO4 binds siRNA, complex binds target & Pol V

15
Generating methylated DNA RDR2 makes it DS, 24 nt siRNA are generated by DCL3 AGO4 binds siRNA, complex binds target & Pol V Pol V makes intergenic RNA, associates with AGO4- siRNA to recruit “silencing Complex” to target site

16
Generating methylated DNA RDR2 makes it DS, 24 nt siRNA are generated by DCL3 AGO4 binds siRNA, complex binds target & Pol V Pol V makes intergenic RNA, associates with AGO4-siRNA to recruit “silencing Complex” to target site Amplifies signal! extends meth- ylated region

17
Using the genome Many sites provide gene expression data online NIH Gene expression omnibus http://www.ncbi.nlm.nih.gov/geo/ provides access to many different types of gene expression data http://www.ncbi.nlm.nih.gov/geo/

18
Using the genome Many sites provide gene expression data online NIH Gene expression omnibus http://www.ncbi.nlm.nih.gov/geo/ provides access to many different types of gene expression data http://www.ncbi.nlm.nih.gov/geo/ Many different sites provide “digital Northerns” or other comparative analyses of gene expression http://cgap.nci.nih.gov/SAGE http://www.weigelworld.org/research/projects/geneexpr essionatlas http://www.weigelworld.org/research/projects/geneexpr essionatlas

19
Using the genome Many sites provide gene expression data online NIH Gene expression omnibus http://www.ncbi.nlm.nih.gov/geo/ provides access to many different types of gene expression data http://www.ncbi.nlm.nih.gov/geo/ Many different sites provide “digital Northerns” or other comparative analyses of gene expression http://cgap.nci.nih.gov/SAGE http://www.weigelworld.org/research/projects/geneexpr essionatlas http://www.weigelworld.org/research/projects/geneexpr essionatlas MPSS (massively-parallel signature sequencing) http://mpss.udel.edu/
")
20
Using the genome Many sites provide gene expression data online NIH Gene expression omnibus http://www.ncbi.nlm.nih.gov/geo/ provides access to many different types of gene expression data http://www.ncbi.nlm.nih.gov/geo/ Many different sites provide “digital Northerns” or other comparative analyses of gene expression http://cgap.nci.nih.gov/SAGE http://www.weigelworld.org/research/projects/geneexpr essionatlas http://www.weigelworld.org/research/projects/geneexpr essionatlas MPSS (massively-parallel signature sequencing) http://mpss.udel.edu/ http://mpss.udel.edu/ Use it to decide which tissues to extract our RNA from
 Use it to decide which tissues to extract our RNA from")
21
Using the genome Many sites provide gene expression data online Many sites provide other kinds of genomic data online http://encodeproject.org/ENCODE/

22
Post-transcriptional regulation Nearly ½ of human genome is transcribed, only 1% is coding 98% of RNA made is non-coding

23
Post-transcriptional regulation Nearly ½ of human genome is transcribed, only 1% is coding 98% of RNA made is non-coding Fraction increases with organism’s complexity

24
Known NcRNAs classes and functions

25
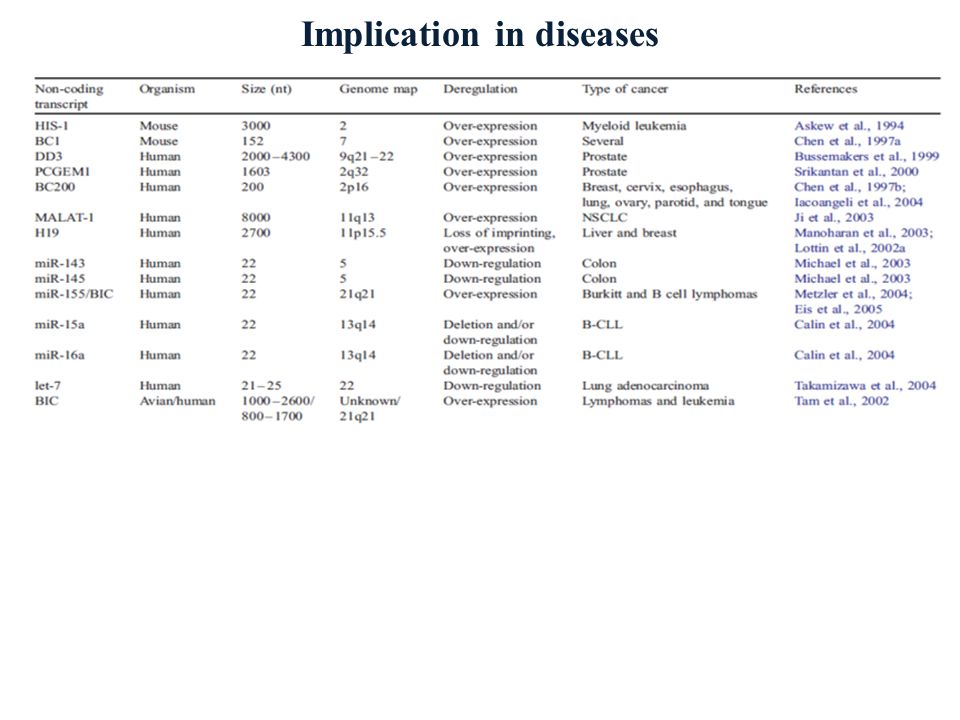
Implication in diseases

27
Transcription in Eukaryotes 3 RNA polymerases all are multi-subunit complexes 5 in common 3 very similar variable # unique ones Plants also have Pols IV & V make siRNA

28
Transcription in Eukaryotes RNA polymerase I: 13 subunits (5 + 3 + 5 unique) acts exclusively in nucleolus to make 45S-rRNA precursor
 acts exclusively in nucleolus to make 45S-rRNA precursor")
29
Transcription in Eukaryotes Pol I: acts exclusively in nucleolus to make 45S-rRNA precursor accounts for 50% of total RNA synthesis

30
Transcription in Eukaryotes Pol I: acts exclusively in nucleolus to make 45S-rRNA precursor accounts for 50% of total RNA synthesis insensitive to -aminitin

31
Transcription in Eukaryotes Pol I: only makes 45S-rRNA precursor 50 % of total RNA synthesis insensitive to -aminitin Mg 2+ cofactor Regulated @ initiation frequency

32
Processing rRNA 1)~ 100 bases are methylated C/D box snoRNA pick sites One for each!
~ 100 bases are methylated C/D box snoRNA pick sites One for each!")
33
Processing rRNA 1)~ 100 bases are methylated C/D box snoRNA pick sites One for each! 2)~ 100 Us are changed to PseudoU H/ACA box snoRNA pick sites One for each!
~ 100 Us are changed to PseudoU H/ACA box snoRNA pick sites One for each!.")
34
Processing rRNA 1)~ 100 bases are methylated C/D box snoRNA pick sites 2)~ 100 Us are changed to PseudoU H/ACA box snoRNA pick sites 3) Some snoRNA direct modification of tRNA and snRNA
~ 100 bases are methylated C/D box snoRNA pick sites 2)~ 100 Us are changed to PseudoU H/ACA box snoRNA pick sites 3) Some snoRNA direct modification of tRNA and snRNA")
35
Processing rRNA 1)~ 200 bases are modified 2) 45S pre-rRNA is cut into 28S, 18S and 5.8S products by ribozymes RNase MRP cuts between 18S & 5.8S U3, U8, U14, U22, snR10 and snR30 also guide cleavage
~ 200 bases are modified 2) 45S pre-rRNA is cut into 28S, 18S and 5.8S products by ribozymes RNase MRP cuts between 18S & 5.8S U3, U8, U14, U22, snR10 and snR30 also guide cleavage")
36
Processing rRNA 1)~ 200 bases are methylated 2) 45S pre-rRNA is cut into 28S, 18S and 5.8S products 3) Ribosomes are assembled w/in nucleolus
~ 200 bases are methylated 2) 45S pre-rRNA is cut into 28S, 18S and 5.8S products 3) Ribosomes are assembled w/in nucleolus")
37
RNA Polymerase III makes ribosomal 5S and tRNA (+ some snRNA, scRNA, etc) >100 different kinds of ncRNA ~10% of all RNA synthesis Cofactor = Mn 2+ cf Mg 2+ sensitive to high [ -aminitin]
![RNA Polymerase III makes ribosomal 5S and tRNA (+ some snRNA, scRNA, etc) >100 different kinds of ncRNA ~10% of all RNA synthesis Cofactor = Mn 2+ cf Mg 2+ sensitive to high [ -aminitin]](http://images.slideplayer.com/25/7959195/slides/slide_37.jpg "RNA Polymerase III makes ribosomal 5S and tRNA (+ some snRNA, scRNA, etc) >100 different kinds of ncRNA ~10% of all RNA synthesis Cofactor = Mn 2+ cf Mg 2+ sensitive to high [ -aminitin]")
38
Processing tRNA 1)tRNA is trimmed 5’ end by RNAse P (1 RNA, 10 proteins)
tRNA is trimmed 5’ end by RNAse P (1 RNA, 10 proteins)")
39
Processing tRNA 1)tRNA is trimmed 2)Transcript is spliced Some tRNAs are assembled from 2 transcripts
tRNA is trimmed 2)Transcript is spliced Some tRNAs are assembled from 2 transcripts")
40
Processing tRNA 1)tRNA is trimmed 2)Transcript is spliced 3)CCA is added to 3’ end By tRNA nucleotidyl transferase (no template) tRNA +CTP -> tRNA-C + PPi tRNA-C +CTP--> tRNA-C-C + PPi tRNA-C-C +ATP -> tRNA-C-C-A + PPi
tRNA is trimmed 2)Transcript is spliced 3)CCA is added to 3’ end By tRNA nucleotidyl transferase (no template) tRNA +CTP -> tRNA-C + PPi tRNA-C +CTP--> tRNA-C-C + PPi tRNA-C-C +ATP -> tRNA-C-C-A + PPi")
41
Processing tRNA 1)tRNA is trimmed 2)Transcript is spliced 3)CCA is added to 3’ end 4)Many bases are modified Protects tRNA Tweaks protein synthesis
tRNA is trimmed 2)Transcript is spliced 3)CCA is added to 3’ end 4)Many bases are modified Protects tRNA Tweaks protein synthesis")
42
Processing tRNA 1)tRNA is trimmed 2)Transcript is spliced 3)CCA is added to 3’ end 4)Many bases are modified 5)No cap! -> 5’ P (due to 5’ RNAse P cut)
.")
43
Splicing: the spliceosome cycle 1) U1 snRNP (RNA/protein complex) binds 5’ splice site
 U1 snRNP (RNA/protein complex) binds 5’ splice site")
44
Splicing:The spliceosome cycle 1) U1 snRNP binds 5’ splice site 2) U2 snRNP binds “branchpoint” -> displaces A at branchpoint
 U1 snRNP binds 5’ splice site 2) U2 snRNP binds branchpoint -> displaces A at branchpoint")
45
Splicing:The spliceosome cycle 1) U1 snRNP binds 5’ splice site 2) U2 snRNP binds “branchpoint” -> displaces A at branchpoint 3) U4/U5/U6 complex binds intron displace U1 spliceosome has now assembled
 U1 snRNP binds 5’ splice site 2) U2 snRNP binds branchpoint -> displaces A at branchpoint 3) U4/U5/U6 complex binds intron displace U1 spliceosome has now assembled")
46
Splicing: RNA is cut at 5’ splice site cut end is trans-esterified to branchpoint A

47
Splicing: 5) RNA is cut at 3’ splice site 6) 5’ end of exon 2 is ligated to 3’ end of exon 1 7) everything disassembles -> “lariat intron” is degraded
 RNA is cut at 3’ splice site 6) 5’ end of exon 2 is ligated to 3’ end of exon 1 7) everything disassembles -> lariat intron is degraded")
48
Splicing:The spliceosome cycle

49
Splicing: Some RNAs can self-splice! role of snRNPs is to increase rate! Why splice?

50
Splicing: Why splice? 1) Generate diversity exons often encode protein domains
 Generate diversity exons often encode protein domains")
51
Splicing: Why splice? 1) Generate diversity exons often encode protein domains Introns = larger target for insertions, recombination
 Generate diversity exons often encode protein domains Introns = larger target for insertions, recombination.")
52
Why splice? 1) Generate diversity >94% of human genes show alternate splicing
 Generate diversity >94% of human genes show alternate splicing")
53
Why splice? 1) Generate diversity >94% of human genes show alternate splicing same gene encodes different protein in different tissues
 Generate diversity >94% of human genes show alternate splicing same gene encodes different protein in different tissues.")
54
Why splice? 1) Generate diversity >94% of human genes show alternate splicing same gene encodes different protein in different tissues Stressed plants use AS to make variant stress-response proteins
 Generate diversity >94% of human genes show alternate splicing same gene encodes different protein in different tissues Stressed plants use AS to make variant stress-response proteins.")
55
Why splice? 1) Generate diversity >94% of human genes show alternate splicing same gene encodes different protein in different tissues Stressed plants use AS to make variant Stress-response proteins Splice-regulator proteins control AS: regulated by cell-specific expression and phosphorylation
 Generate diversity >94% of human genes show alternate splicing same gene encodes different protein in different tissues Stressed plants use AS to make variant Stress-response proteins Splice-regulator proteins control AS: regulated by cell-specific expression and phosphorylation.")
56
Why splice? 1)Generate diversity Trabzuni D, et al (2013)Nat Commun. 22:2771. Found 448 genes that were expressed differently by gender in human brains (2.6% of all genes expressed in the CNS). All major brain regions showed some gender variation, and 85% of these variations were due to RNA splicing differences
. All major brain regions showed some gender variation, and 85% of these variations were due to RNA splicing differences.")
57
Why splice? 1)Generate diversity Wilson LOW, Spriggs A, Taylor JM, Fahrer AM. (2014). A novel splicing outcome reveals more than 2000 new mammalian protein isoforms. Bioinformatics 30: 151-156 Splicing created a frameshift, so was annotated as “nonsense-mediated decay” an alternate start codon rescued the protein, which was expressed
. A novel splicing outcome reveals more than 2000 new mammalian protein isoforms. Bioinformatics 30: Splicing created a frameshift, so was annotated as nonsense-mediated decay an alternate start codon rescued the protein, which was expressed.")
58
Why splice? Splicing created a frameshift, so was annotated as “nonsense-mediated decay” an alternate start codon rescued the protein, which was expressed Found 1849 human & 733 mouse mRNA that could encode alternate protein isoforms the same way So far 64 have been validated by mass spec

59
Regulatory ncRNA 1.SiRNA direct DNA-methylation via RNA-dependent DNA-methyltansferase 2. In other cases direct RNA degradation

60
mRNA degradation lifespan varies 100x Sometimes due to AU-rich 3' UTR sequences Defective mRNA may be targeted by NMD, NSD, NGD Other RNA are targeted by small interfering RNA

61
Other mRNA are targeted by small interfering RNA defense against RNA viruses DICERs cut dsRNA into 21-28 bp

62
Other mRNA are targeted by small interfering RNA defense against RNA viruses DICERs cut dsRNA into 21-28 bp helicase melts dsRNA

63
Other mRNA are targeted by small interfering RNA defense against RNA viruses DICERs cut dsRNA into 21-28 bp helicase melts dsRNA - RNA binds RISC

64
Other mRNA are targeted by small interfering RNA defense against RNA viruses DICERs cut dsRNA into 21-28 bp helicase melts dsRNA - RNA binds RISC complex binds target

65
Other mRNA are targeted by small interfering RNA defense against RNA viruses DICERs cut dsRNA into 21-28 bp helicase melts dsRNA - RNA binds RISC complex binds target target is cut

66
Small RNA regulation siRNA: target RNA viruses (& transgenes) miRNA: arrest translation of targets created by digestion of foldback Pol II RNA with mismatch loop
 miRNA: arrest translation of targets created by digestion of foldback Pol II RNA with mismatch loop")
67
Small RNA regulation siRNA: target RNA viruses (& transgenes) miRNA: arrest translation of targets created by digestion of foldback Pol II RNA with mismatch loop Mismatch is key difference: generated by different Dicer
 miRNA: arrest translation of targets created by digestion of foldback Pol II RNA with mismatch loop Mismatch is key difference: generated by different Dicer")
68
Small RNA regulation siRNA: target RNA viruses (& transgenes) miRNA: arrest translation of targets created by digestion of foldback Pol II RNA with mismatch loop Mismatch is key difference: generated by different Dicer Arrest translation in animals, target degradation in plants
 miRNA: arrest translation of targets created by digestion of foldback Pol II RNA with mismatch loop Mismatch is key difference: generated by different Dicer Arrest translation in animals, target degradation in plants")
69
small interfering RNA mark specific targets once cut they are removed by endonuclease-mediated decay

71
Most RNA degradation occurs in P bodies recently identified cytoplasmic sites where exosomes & XRN1 accumulate when cells are stressed

72
Most RNA degradation occurs in P bodies recently identified cytoplasmic sites where exosomes & XRN1 accumulate when cells are stressed Also where AGO & miRNAs accumulate

73
Most RNA degradation occurs in P bodies recently identified cytoplasmic sites where exosomes & XRN1 accumulate when cells are stressed Also where AGO & miRNAs accumulate w/o miRNA P bodies dissolve!

74
Thousands of antisense transcripts in plants 1.Overlapping genes

75
Thousands of antisense transcripts in plants 1.Overlapping genes 2.Non-coding RNAs

76
Thousands of antisense transcripts in plants 1.Overlapping genes 2.Non-coding RNAs 3.cDNA pairs

77
Thousands of antisense transcripts in plants 1.Overlapping genes 2.Non-coding RNAs 3.cDNA pairs 4.MPSS

78
Thousands of antisense transcripts in plants 1.Overlapping genes 2.Non-coding RNAs 3.cDNA pairs 4.MPSS 5.TARs

79
Thousands of antisense transcripts in plants Hypotheses 1.Accident: transcription unveils “cryptic promoters” on opposite strand (Zilberman et al)
")
80
Hypotheses 1. Accident: transcription unveils “cryptic promoters” on opposite strand (Zilberman et al) 2. Functional a.siRNA b.miRNA c.Silencing
 2. Functional a.siRNA b.miRNA c.Silencing.")
81
Hypotheses 1. Accident: transcription unveils “cryptic promoters” on opposite strand (Zilberman et al) 2. Functional a.siRNA b.miRNA c.Silencing d.Priming: chromatin remodeling requires transcription!
 2. Functional a.siRNA b.miRNA c.Silencing d.Priming: chromatin remodeling requires transcription!.")
Similar presentations





>")

 Map the genome 2) Prepare an AC library 3) Order the library 4) Subdivide each AC into lambda contigs 5) Subdivide each lambda into.>")









>")


