Download presentation
Presentation is loading. Please wait.

1
A survey of what has been learned from individual-based models Don DeAngelis, USGS and University of Miami

2
What do we learn from models? Every important modeling approach, even if initially having practical aims, eventually leads to major changes in thinking: Dynamic simulation models - used to model urban, global, and ecosystem dynamics. They revealed the importance of feedback and unexpected indirect effects (e.g., processes are usually more important than initial conditions). Non-linear differential and difference equation models - revealed the possibility of limit cycles, chaotic behavior (which stress importance of initial conditions – butterfly effect!), mathematical catastrophes, etc. Cellular automata models - revealed that complexity may be generated by very simple rules; perhaps even that “will make it possible to construct a single truly fundamental theory of physics... (Wolfram 2002).
. Non-linear differential and difference equation models - revealed the possibility of limit cycles, chaotic behavior (which stress importance of initial conditions – butterfly effect!), mathematical catastrophes, etc. Cellular automata models - revealed that complexity may be generated by very simple rules; perhaps even that will make it possible to construct a single truly fundamental theory of physics... (Wolfram 2002)..")
3
What do we learn from models? Besides these broad conceptual advances, one can also point to many specific theoretical results that emerged from the ‘classical’ models of ecology: Reaction-diffusion models - traveling waves, diffusional instabilities Matrix age-structure models - existence of asymptotic stable age distributions (inherent in eigenvalues/eigenvectors) Game theory models - evolutionarily stable strategies
 Game theory models - evolutionarily stable strategies.")
4
The ultimate importance of any new approach may be unanticipated Perhaps the real importance of a modeling approach may eventually be gauged more by its revealing general and unexpected properties of systems (theoretical advances), and changing our way of thinking, than simply by its ability to provide good descriptions or predictions for systems. A modeling approach may be developed with rather limited pragmatic objectives to begin with, but may lead us in ‘its own’ directions towards broader theoretical insights. Does individual-based modeling (IBM) have this sort of importance?
 have this sort of importance .")
5
IBM as a distinct method of inquiry in some sciences In some fields shifts in modeling perspective from macro to micro (individual-based) scale, have led to new ideas and progress. To cite an early exponent of the micro approach (at least conceptually) in physics: “... the physics of low density plasmas has been to me a science in which all phenomena should be described from the point of view of individual particles. Instead of treating hydromagnetic equations, I prefer to sit and ride on each electron and ion and try to describe what the world is like from their point of view and what forces push them to the left or to the right.” Hannes Alfvén, “Memoirs of a dissident scientist”, American Scientist 76, page 250 The use of IBM in physics is now fundamental, from electrons to galaxies.
 in physics: ... the physics of low density plasmas has been to me a science in which all phenomena should be described from the point of view of individual particles. Instead of treating hydromagnetic equations, I prefer to sit and ride on each electron and ion and try to describe what the world is like from their point of view and what forces push them to the left or to the right. Hannes Alfvén, Memoirs of a dissident scientist , American Scientist 76, page 250 The use of IBM in physics is now fundamental, from electrons to galaxies..")
6
IBM as a distinct method of inquiry in some sciences This is also true in economics, although the emphasis on the individual per se is not new: “... [Up until the 1930s] it was widely believed that economics should be built on a structure resting on a few simple premises about the behavior of microcomponents [individuals]” “Because the demands on data and analytic ability were high, economists turned to macro time series.” “However, the micro-macro pendulum gradually started to swing back again.... the data and tools now available make possible the microanalytic paradigm for modeling a social system.” Orcutt et al. (1976)
.")
7
What overall significance does IBM represent in ecology? Uchmanski and Grimm (1996) argued that IBM (in the narrowly defined sense - see later) by its nature constitutes a significantly different class of models (or a different paradigm) than the classical models of ecology (ode, pde, and matrix models). Bolker et al. (1997) replied that “separating IBM from the rest of classical model is unnecessary and interferes with the insightful process of comparing models with different levels of detail.” In the sense that such models are affecting our fundamental theoretical views, the answer is perhaps not certain yet and will depend on whether this type of modeling affects our conceptual views and the type of theory that we develop.
 argued that IBM (in the narrowly defined sense - see later) by its nature constitutes a significantly different class of models (or a different paradigm) than the classical models of ecology (ode, pde, and matrix models). Bolker et al. (1997) replied that separating IBM from the rest of classical model is unnecessary and interferes with the insightful process of comparing models with different levels of detail. In the sense that such models are affecting our fundamental theoretical views, the answer is perhaps not certain yet and will depend on whether this type of modeling affects our conceptual views and the type of theory that we develop..")
8
What overall significance does IBM represent in ecology? That does not seem to be happening rapidly. Grimm’s (1999) survey of IBM noted: the use of IBM has been motivated during the past 10 years primarily on the pragmatic basis of providing a better description of ecological systems. there has been little attention to its potential theoretical advances; i.e., addressing paradigmatic questions. Similar points were made by Railsback (2001); i.e., that IBM has focused on providing detailed empirical descriptions, but not innovative theory. If IBM is new in a deep sense, it should lead to more than a better description of ecological systems.
 survey of IBM noted: the use of IBM has been motivated during the past 10 years primarily on the pragmatic basis of providing a better description of ecological systems. there has been little attention to its potential theoretical advances; i.e., addressing paradigmatic questions. Similar points were made by Railsback (2001); i.e., that IBM has focused on providing detailed empirical descriptions, but not innovative theory. If IBM is new in a deep sense, it should lead to more than a better description of ecological systems..")
9
What overall significance does IBM represent in ecology? In ecology, theory at the macro and micro levels has been largely separate: During most of the last 50 years, at least, the emphasis in population theory has been on analytic models at the macro level and top-down explanations. At the same time rule-based, optimization or game theory modeling has been applied to individual behaviors and social organization. Much of the significance of IBM lies in its potential to develop micro scale explanations for phenomena at the macro level.

10
What overall significance does IBM represent in ecology? This is far from a reality now. However it is possible to outline the progress in this direction. Generally, following Uchmanski and Grimm’s (1996), one can say that individual-based models are arrayed along a path from ‘broad’ to ‘narrow’ individual-based models, or towards greater complexity with which the individual’s life cycle is represented greater degree of modeling the dynamics of the resources the use of integer numbers (rather than number density) in representing population size Inclusion of variability of individuals of the same age.
, one can say that individual-based models are arrayed along a path from ‘broad’ to ‘narrow’ individual-based models, or towards greater complexity with which the individual’s life cycle is represented greater degree of modeling the dynamics of the resources the use of integer numbers (rather than number density) in representing population size Inclusion of variability of individuals of the same age..")
11
Does use of IBM represent a significant new approach in ecology? We can think of an ‘axis’ from broad to narrow IBM. This is not necessarily the same thing as an axis from ‘simple’ to ‘complex’, or from ‘better’ to ‘worse’ or vice versa. Me might think of it as an axis leading in the direction of greater individual structure or autonomy. The next graph attempts to organize the ways in which IBM has, at least modestly, provided insights and maybe modified our theory and thinking about ecological systems. Perhaps the direction of this axis points towards major changes in theory, or paradigm shifts.

12
Schematic of increasing individual autonomy in IBM

13
Level 1of IBM continuum: Discreteness and local interactions These are models that take into account the discreteness of individuals in a spatial lattice or continuum. No or limited individual variation. Examples: Theoretical models to explore effects of discreteness, local interaction, clustering, temporal forcing on the equilibrium values, stability, and emergence of spatial patterns of interacting populations. Often inspired by physical models, such as Ising model for phase transitions; e.g., - Breckling 1990, de Roos et al. 1991, Smith 1991, Durrett and Levin 1994, Pascual and Levin 1999, Gandhi et al. 1999, Cuddington and Yodzis 2000

14
Level 1of IBM continuum: Discreteness and local interactions Example: de Roos et al. (1991) Question: How does individual mobility affect dynamics and patterns of predator and prey populations? Model system: 128 x 128 position grid, empty or with one prey and or predator Mechanisms: Prey individuals reproduce probabilistically, but only if unoccupied site exist. Predators can eat prey if they land on same cell and have not eaten for number of time steps (handling time). Predators can reproduce after F meals. Results: Limited individual mobility greatly reduces the fluctuation in total density, though does not affect average densities. Suggested that ephemeral local concentrations of predators and prey are generated in the model, and that these fluctuate out of phase, leading to greater stability over the whole region.
 Question: How does individual mobility affect dynamics and patterns of predator and prey populations. Model system: 128 x 128 position grid, empty or with one prey and or predator Mechanisms: Prey individuals reproduce probabilistically, but only if unoccupied site exist. Predators can eat prey if they land on same cell and have not eaten for number of time steps (handling time). Predators can reproduce after F meals. Results: Limited individual mobility greatly reduces the fluctuation in total density, though does not affect average densities. Suggested that ephemeral local concentrations of predators and prey are generated in the model, and that these fluctuate out of phase, leading to greater stability over the whole region..")
15
Level 1of IBM continuum: Discreteness and local interactions What insights, or generalities, have been gained from these models? Because individuals are discrete and interactions are local, the mobility and movement rules of individual animals can affect population level properties (stability, population sizes). Diffusion limitation (or local negative correlations of predators and prey) may affect population sizes and, along with spatial pattern formation (out- of-phase oscillations, ‘ephemeral refuges’), stability.
. Diffusion limitation (or local negative correlations of predators and prey) may affect population sizes and, along with spatial pattern formation (out- of-phase oscillations, ‘ephemeral refuges’), stability..")
16
Level 2 of IBM continuum: Finite numbers of individuals Because of the discreteness of individuals, absolute numbers of individuals, not just densities, are conveniently modeled. Examples: Simulation of community of competing trees shows the strong effect of size of area on the time to extinction or dominance of populations; - Hubbell and Foster 1986 Simulation of discrete prey and predators in space, where the mean population size can be varied, shows that the time to extinction in the system increases as mean population size increases, but this happens in a complex way due to the nature of the effects of space - not explicable, say, from a classical birth-and-death model; - Donalson and Nisbet 1999

17
Level 2 of IBM continuum: Finite numbers of individuals What insights have been gained from these models? These models have led to a number of generalizations regarding the effects of system size and spatial aspects on extinction. Classical models cannot deal as effectively with such issues.

18
Level 3 of IBM continuum: Variation among individuals Individuals within a population differ, not only as a function of age, but within age classes as well. Examples: Simulation of a population of individuals in non-overlapping generations. The growth parameters vary among the individuals such that a size distribution is produced. - Grimm and Uchmanski (2002)
.")
19
Level 3 of IBM continuum: Variation among individuals What insights have been gained from this model? Intra-generational dynamics may be important in populations of non- overlapping generations. If fast, intra-generational dynamics dominate, this can help stabilize a consumer-resource system, as the consumer population number will ‘adjust’ to the resource and not overexploit it. Increased internal variance in individual variables (e.g.,weight) can help stabilize a population and increase its persistence.
 can help stabilize a population and increase its persistence..")
20
Level 3 to 4 of IBM continuum: Moderately complex life histories and behaviors The addition of complex life cycles and behaviors can be done in a simple way, ignoring bioenergetics, learning, etc., but including rules for behavior. Spatio-temporal environmental variation can generate variation within a population, but it is not self-ampifying. Examples: Simple types of population models that do not have bioenergetics or growth models; e.g., - Cape Sable Sparrow model (Nott 1998), snail kite model (Mooij et al., 2002) Within-population variation (successful vs. unsuccessful nesters) occurs, but does not create amplifying feedbacks.
, snail kite model (Mooij et al., 2002) Within-population variation (successful vs. unsuccessful nesters) occurs, but does not create amplifying feedbacks..")
21
Level 3 to 4 of IBM continuum: Moderately complex life histories and behaviors What insights can be gained from these models? These are models in which individuals have some autonomy, through rules for dealing with certain situations. The individuals can, for example, employ movement or other strategies. These behaviors can have major effects on the population dynamics.

22
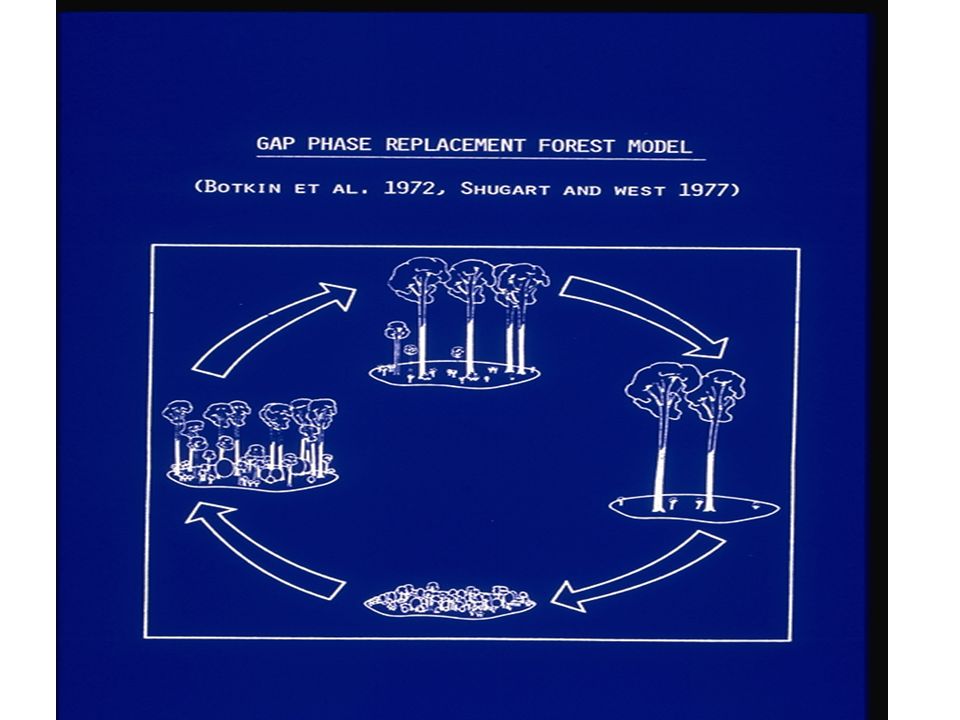
Level 4 of IBM continuum: Addition of growth and size dependence to life histories, and interactions among individuals When growth and size-dependent interactions are added to life cycle models, the models can be used to simulate competition resulting from local interactions. The individuals now are characterized by individual growth rates that can change in response to effects of neighbors. Examples: Forest gap phase replacement models that allow competing tree life cycles in a gap (not necessarily spatially explicit). The models are not spatially explicit, but simulate small enough areas (0.1 ha treefall gaps) that all trees can be assumed to interact, - Botkin et al. 1972, Shugart 1984
. The models are not spatially explicit, but simulate small enough areas (0.1 ha treefall gaps) that all trees can be assumed to interact, - Botkin et al. 1972, Shugart")
24
Level 4 of IBM continuum: Addition of growth and and size dependence to life histories, and interactions among individuals What insights have been gained from these model types? The feedback effects between individual organisms (trees) and the entire community are able to generate community succession within a gap. This modeling approach carries the ‘relay floristics’ theoretical concept of secondary succession. The models helped increase appreciation of the non-equilibrium and stochastic nature of competition within gaps - though leading to a relatively stable community structure on a large forest landscape. (These models approach Uchmanski and Grimm’s definition of ‘narrow’ IBM.)
 and the entire community are able to generate community succession within a gap. This modeling approach carries the ‘relay floristics’ theoretical concept of secondary succession. The models helped increase appreciation of the non-equilibrium and stochastic nature of competition within gaps - though leading to a relatively stable community structure on a large forest landscape. (These models approach Uchmanski and Grimm’s definition of ‘narrow’ IBM.).")
25
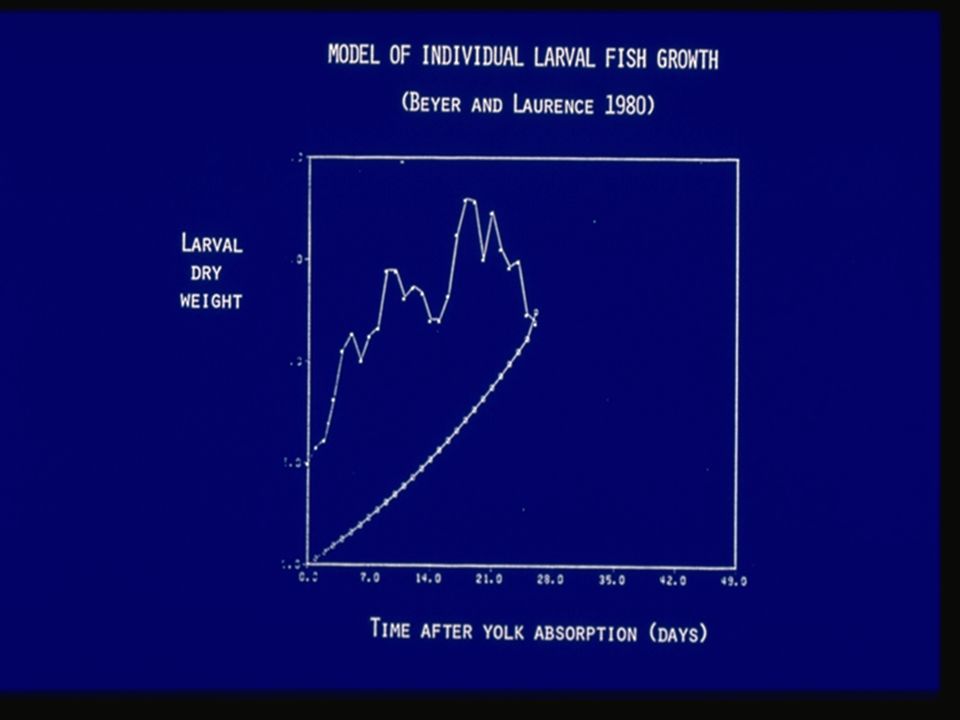
Level 4 of IBM continuum: Life histories where growth rates of individuals tend to be temporally autocorrelated When daily foraging has both stochastic and size-dependent components, then the internal state of an organism (e.g., size) may tend to feed back in a self-amplifying way. Examples; Models of a number of organisms in which there are positive correlations between growth on one time interval and that on the succeeding time interval; e.g., -Beyer and Laurence 1980, Crowder et al. 1993, Pfister and Stevens 2002. The next figure shows growth of larval fish in which there is an autocorrelation in growth from one day to the next.

28
Level 4 of IBM continuum: Life histories where growth rates of individuals tend to be temporally autocorrelated What insights have been gained from these models? These models are able to produce population-level and size structure phenomena. Stochasticity gives rise to minor inequalities in foraging success, which can lead to differences in growth (physiological ‘memory’), causing further divergence in size. Such self-amplifying effects cannot easily be shown in McKendrick- von Foerster or similar classical models.
, causing further divergence in size. Such self-amplifying effects cannot easily be shown in McKendrick- von Foerster or similar classical models..")
29
Level 4 of IBM continuum: Life histories where growth leads to strong size-dependent effects on interactions between individuals When daily foraging has both stochastic and size-dependent components, then the internal state of an organism (e.g., size) may tend to feed back in a self-amplifying way. Examples; Models of growth of plants and some animals (fish, marine invertebrates) with strong size-dependence on foraging and survival; -Ford and Diggle 1981, Scheffer et al. 1995, Lamberson, Railsback, Harvey (2002?) This can be especially pronounced when there is cannibalism within a cohort. The next figures show two cohorts started off with slightly different initial size distributions.
 with strong size-dependence on foraging and survival; -Ford and Diggle 1981, Scheffer et al. 1995, Lamberson, Railsback, Harvey (2002 ) This can be especially pronounced when there is cannibalism within a cohort. The next figures show two cohorts started off with slightly different initial size distributions..")
32
Level 4 of IBM continuum: Life histories where growth leads to strong size-dependent effects on interactions between individuals What insights have been gained from these models? A great variety of (often counterintuitive) results are produced. Simulation outcomes may be extremely sensitive to initial size distributions, sometimes leading to large populations of stunted individuals and sometimes to a few large individuals. Rapid shifts from starvation-controlled to predation controlled mortality rates can occur. Bimodal size distributions, as well as Ricker-type (unimodal) stock- recruitment relationships can emerge fairly easily.
 results are produced. Simulation outcomes may be extremely sensitive to initial size distributions, sometimes leading to large populations of stunted individuals and sometimes to a few large individuals. Rapid shifts from starvation-controlled to predation controlled mortality rates can occur. Bimodal size distributions, as well as Ricker-type (unimodal) stock- recruitment relationships can emerge fairly easily..")
33
Level 4 of IBM continuum: Interaction of life histories, growth, and spatial gradients in systems Example: (Marine intertidal model, Robles and Descharnais 2002) A model of predator-prey interactions has been developed for the intertidal to attempt to explain the main features of spatial patterns. Main Variables: Spatial cells that can be empty or occupied, size and positions of individual mussels Main Mechanisms: Duration of inundation of cells, and exposure to waves of various intensities, recruitment of larval mussels, effects of elevation, wave intensity, and local mussel density on recruitment to various locations; mussel growth, effects of elevation and wave action on growth; predation by seastars on mussels, effects of elevation, wave action, mussel size, and mussel aggregation on predation

34
Level 4 of IBM continuum: Interaction of life histories, growth, and spatial gradients in systems What insights have been gained from this model type? The model allowed many intertidal factors (competition for space, predation, wave energy) to be included at once and provided the basis for testing a number of hypotheses, particularly in the way that gradients of elevation and intensity of wave action would affect the patterns (size-structure, biomass) of mussels. Also, the model was able to demonstrate that there was no need to include and additional specific mechanism of spatial refugia from predation for a certain fraction of mussels. The combined effects of all the other mechanisms created an “effective” refugium, an emergent phenomenon. (This elimination of the need for ad hoc assumptions is characteristic of IBM at this level.)
 to be included at once and provided the basis for testing a number of hypotheses, particularly in the way that gradients of elevation and intensity of wave action would affect the patterns (size-structure, biomass) of mussels. Also, the model was able to demonstrate that there was no need to include and additional specific mechanism of spatial refugia from predation for a certain fraction of mussels. The combined effects of all the other mechanisms created an effective refugium, an emergent phenomenon. (This elimination of the need for ad hoc assumptions is characteristic of IBM at this level.).")
35
Level 4 of IBM continuum: Addition of detailed behavioral aspects These models included behavioral information at the level of individuals that lead to population and community level effects. Examples: Tritrophic level model (Schmitz 2000) Main Variables: 2-dimensional grid landscape, nutrient resources, individual grass and herb plants, grasshoppers and spiders. Main Mechanisms: Depletion of resources, seasonality, foraging, consumption of resources, growth, metabolism, nutrient uptake and competition of plants, movement (random and directed) of animals, prey detection radiuses of individuals, reproduction, withering of plants, predator avoidance behavior by grasshoppers.
 Main Variables: 2-dimensional grid landscape, nutrient resources, individual grass and herb plants, grasshoppers and spiders. Main Mechanisms: Depletion of resources, seasonality, foraging, consumption of resources, growth, metabolism, nutrient uptake and competition of plants, movement (random and directed) of animals, prey detection radiuses of individuals, reproduction, withering of plants, predator avoidance behavior by grasshoppers..")
36
Level 4 of IBM continuum: Addition of detailed behavioral aspects What generalities have been gained from this model type? Schmitz’s (2000) model showed that behavioral details, especially risk avoidance by the grasshopper, did much to structure the trophic web. The presence of spiders, even when they did not prey on the grasshoppers, but merely changed their behaviors, led to an increase in grass biomass compared with simulations where the spiders were not present. What the model also showed was that for this system short term dynamics within a growing season determined the long-term structure of the community. Other examples: Models such as those of Kaiser (1979) and Pettifor et al. (2000) show that ‘carrying capacity’ emerges from behavioral mechanisms in the model. Basset et al. (2002) show that when individual foragers optimized the ‘leaving time’ from a patch, this decreased predator-prey stability.
 model showed that behavioral details, especially risk avoidance by the grasshopper, did much to structure the trophic web. The presence of spiders, even when they did not prey on the grasshoppers, but merely changed their behaviors, led to an increase in grass biomass compared with simulations where the spiders were not present. What the model also showed was that for this system short term dynamics within a growing season determined the long-term structure of the community. Other examples: Models such as those of Kaiser (1979) and Pettifor et al. (2000) show that ‘carrying capacity’ emerges from behavioral mechanisms in the model. Basset et al. (2002) show that when individual foragers optimized the ‘leaving time’ from a patch, this decreased predator-prey stability..")
37
Level 4 of IBM continuum: More complex behaviors These are models in which individuals respond to their environment and each other through complex behavioral rules, based on internal states (e.g. bioenergetics) and other factors. Example: Model of a wood stork breeding colony (Wolff 1994) Main variables: Location, energy balance, amount of captured prey of each adult wood stork; energy balance of offspring Main mechanisms: Foraging decisions; when to leave nest to start foraging, where to forage, when to move, captures of prey, return to nest, feeding of offspring., This and similar models contain bioenergetics submodels, and the animals make decisions based on their internal states as well as varying environmental conditions.
 and other factors. Example: Model of a wood stork breeding colony (Wolff 1994) Main variables: Location, energy balance, amount of captured prey of each adult wood stork; energy balance of offspring Main mechanisms: Foraging decisions; when to leave nest to start foraging, where to forage, when to move, captures of prey, return to nest, feeding of offspring., This and similar models contain bioenergetics submodels, and the animals make decisions based on their internal states as well as varying environmental conditions..")
40
Level 4 of IBM continuum: More complex behaviors What insights have been gained from these models? Individual organisms ‘solve’ the problem of a changing environment through appropriate rule-based actions to deal with their own situation in time and space. Complex sequences of behavior occur, with animals making decisions. Collectively, this adds up to a whole population interacting with its environment. (Classical models, such as the Ideal Free Distribution for foraging, would not work. Decisions are highly contingent on circumstances, and on less than global knowledge.) Such models are a step in the direction of describing how real populations deal with a complex environment, through individual behaviors. These have brought environment back in a basic way in population models.
 Such models are a step in the direction of describing how real populations deal with a complex environment, through individual behaviors. These have brought environment back in a basic way in population models..")
41
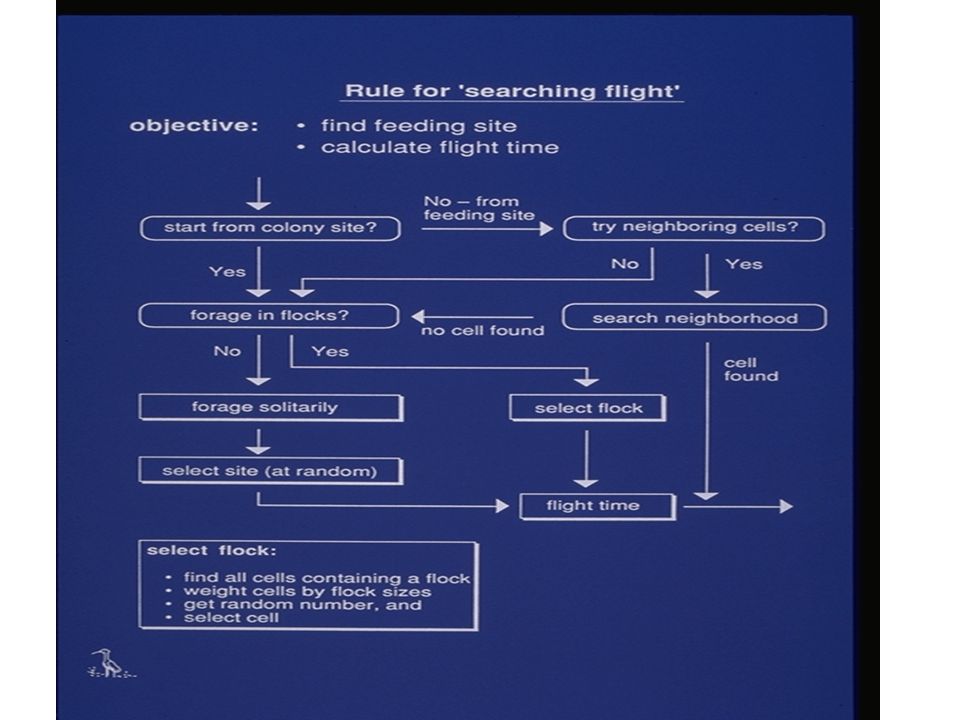
Level 5 of IBM continuum: Individuals as adaptive systems In these models, individuals not only have a repertoire of behavioral rules, but the rules can change as animals encounter different situations and learn. Examples: Using a ‘memory net’, model deer are able to accumulate knowledge about its patchy environment (Folse et al. 1989) Neural net approach was used to allow egrets to learn about their patchy environment. Each egret had at any time, only a limited view of the environment. The knowledge individuals gained eventually determined whether the egrets foraged in flocks or solitarily and whether they nested in colonies or alone (Toquenaga et al. 1995). (Also Strand et al. 2002, for fish life history).
 Neural net approach was used to allow egrets to learn about their patchy environment. Each egret had at any time, only a limited view of the environment. The knowledge individuals gained eventually determined whether the egrets foraged in flocks or solitarily and whether they nested in colonies or alone (Toquenaga et al. 1995). (Also Strand et al. 2002, for fish life history)..")
42
Level 5 of IBM continuum: Individuals as adaptive systems What insights have been gained from these models? A population’s exploitation of resources may be modeled to allow the individual organisms, within their bioenergetic, time, and knowledge constraints, to improve efficiency through learning. This may come close to representing how animals actually behave. This has relevance to theory. The main point made by Railsback was that most published IBMs are based on empirical relationships for specific cases, not mechanistic models based on general ideas of fitness maximization in response to an environment. Railsback recommended integrating complex adaptive system (CAS) theory with IBM. As far as the ‘simple/complex’ dichotomy is concerned, models of such type could be called simple, since the organisms are ‘doing the work’, based on general rules of learning.
 theory with IBM. As far as the ‘simple/complex’ dichotomy is concerned, models of such type could be called simple, since the organisms are ‘doing the work’, based on general rules of learning..")
43
Level 5+ of IBM continuum: Evolving adaptive populations In these models, genetic variation occurs among the life cycle and/or behavioral strategies of individuals. Natural selection on average eliminates some, so that evolution may occur. Examples: Rice (1987) developed a simple model for hypothetical sympatric speciation. The techniques for doing this are developing rapidly, such that more realistic cases can be considered (Huse and colleagues).
 developed a simple model for hypothetical sympatric speciation. The techniques for doing this are developing rapidly, such that more realistic cases can be considered (Huse and colleagues)..")
44
Level 5+ of IBM continuum: Evolving adaptive populations What generalities have been gained from these models? Sympatric speciation seems quite feasible under certain circumstances.

45
Is a paradigmatic use of IBM emerging? The integration of CAS theory with IBM could presage a major advance in theory. It requires taking into account the important aspects of individuals as complex adaptive systems. The models of Folse et al. (1989), Toqeunaga (1995), Strand et al. (2002) and others, show that this can be done. The important steps of discreteness, locality, small number, space, complex life cycle and decisions have been combined. But these have generally been limited to ‘tactical’ models with modest objectives. Huse’s appears to be one of the first that really addresses the major problems (e.g., speciation and biodiversity) of classical ecology and evolutionary theory.
, Toqeunaga (1995), Strand et al. (2002) and others, show that this can be done. The important steps of discreteness, locality, small number, space, complex life cycle and decisions have been combined. But these have generally been limited to ‘tactical’ models with modest objectives. Huse’s appears to be one of the first that really addresses the major problems (e.g., speciation and biodiversity) of classical ecology and evolutionary theory..")
46
Is a paradigmatic use of IBM emerging? Perhaps it is almost feasible to attempt to understand, or even predict the diversity within a community or guild, through CAS/IBM. Consider the wading bird community of the Everglades, for example, which supports about 16 different wading birds. A model of the complex environment and resources, combined with the repertoire of possible wading bird foraging strategies and bioenergetics, may be able to do this. Is there enough information to parameterize such a model? Probably only partly, but theoretical models can rely on reasonable assumptions for some parameters.

47
Example: Wading Bird Community Based on:Kushlan, 1978 The wading bird community faces a complex environment, which is a heterogeneous mixture of: -Marsh -Wet prairie -Slough -Mud bank -Top of reef -Woody edge -etc.

48
Wading Bird Community Vegetation types - Sawgrass -Spike rush -Cattail Water relations - Depth -Still or flowing -etc. Prey - Size of prey -Prey shape -Prey density -Prey behaviors -etc.

49
Is a paradigmatic use of IBM emerging? Within the community are a large repertoire of anatomical, physiological, and behavioral characteristics for extracting energy and nutrients from this environment. Observational data is accumulating on the effectiveness of various wading birds to extract energy from conditions of water depth, fish density, and size distributions (Gawlik and colleagues).
..")
50
. Repertoire of Feeding Behaviors: -Standing-Bill vibrating -Baiting-Standing flycatching -Gleaning-Head swaying -Neck swaying-Head swinging -Probing-Pecking -Walking slowly-Walking quickly -Running-Hopping -Leapfrog feeding-Wing flicking -Openwing feeding -Underwing feeding -Double-wing feeding-Canopy feeding -Foot stirring-Foot raking -Foot probing-Hovering -Hovering stirring-Dipping -Foot dragging-Aerial catching -Plunging-Diving -Feetfirst diving-Jumping -Swimming feeding

51
Other behaviors: Foraging schedules: -Crepuscular -Diurnal -Nocturnal -Keyed to tides -Keyed to thermals Territorial defense -Territorial size -Where defended -When defended Social behaviors -Aggregations (feeding in flocks) -Colonial nesting -Prey robbing -Mating patterns -Help to young
 -Colonial nesting -Prey robbing -Mating patterns -Help to young")
52
Feeding structures : - Bill thickness -Bill length -Bill curvature -Gape -Leg length -Trunk shape -Size -Toe size -Neck length -Eye positions

53
Some features of such a model Energy is the main currency. Complex, dynamic landscape is modeled. Model allows individuals to use a repertoire of behavioral strategies and tactics to exploit prey on this landscape. Constraints on speed of movement, prey extraction rates, weight of prey carried to nest, are imposed. Low condition factors, caused by energy deficiencies, cause nest abandonment or death.

54
Is a paradigmatic use of IBM emerging? Such applications of IBM to major issues of theoretical ecology may not occur soon, but they may be needed, as it is questionable whether classical models of ecology can actually solve these problems. As Schrader-Frechette and McCoy (1993) point out, "... although ecologists' mathematical models may have substantial heuristic power, it may be unrealistic to think that they will ever develop into general laws that are universally applicable and able to provide precise predictions for environmental applications." Generalizations stemming from simple, abstract models are vague, often contradictory, and hotly debated by ecologists (Schrader-Frechette and McCoy). Development of CAS/IBM models of semi-realistic communities, with which simulation ‘experiments’ can be performed, may be a promising theoretical approach.
 point out, ... although ecologists mathematical models may have substantial heuristic power, it may be unrealistic to think that they will ever develop into general laws that are universally applicable and able to provide precise predictions for environmental applications. Generalizations stemming from simple, abstract models are vague, often contradictory, and hotly debated by ecologists (Schrader-Frechette and McCoy). Development of CAS/IBM models of semi-realistic communities, with which simulation ‘experiments’ can be performed, may be a promising theoretical approach..")
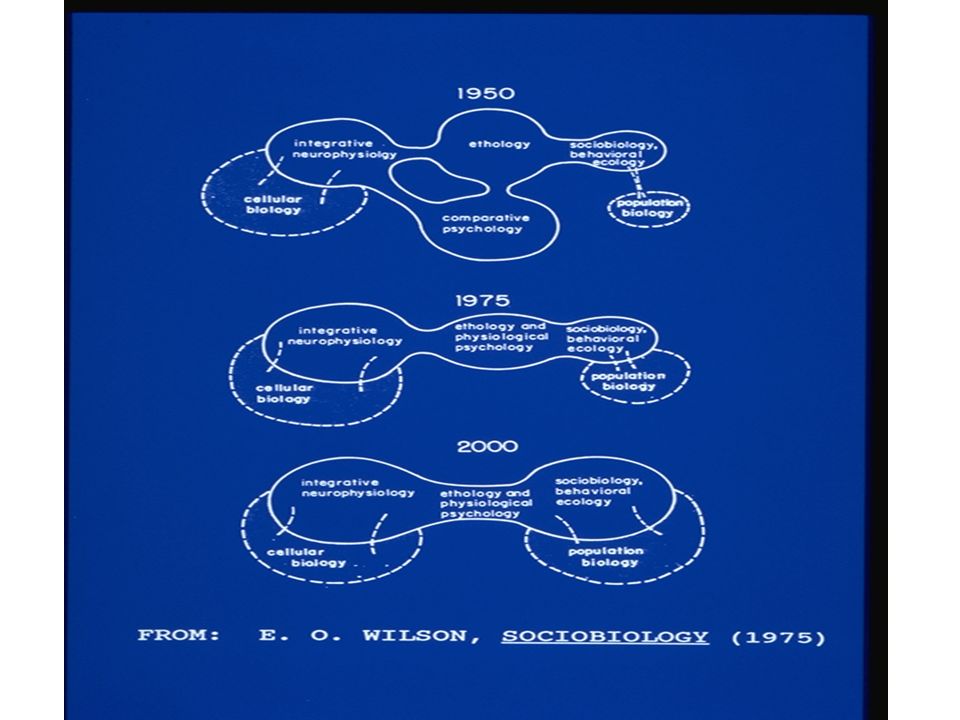
55
Is a paradigmatic use of IBM emerging? In the realm of theory, the individual-based approach provides an avenue for important theoretical progress in ecology. E. O. Wilson (1975) forecast that behavioral ecology and population ecology would be tightly interfaced by the end of the 20th century. Much of this interfacing, if it is to occur, will be accomplished through the extension of population models to incorporate the behavior and energetics of individual organisms in a realistic way. This will pave the way towards theory reduction, or interpreting the "higher level phenomena" of population dynamics in terms of "lower level processes" or mechanisms at the individual level (Schrader-Frechette and McCoy 1993). Because theory reduction is one of the ultimate goals of science, and because theory reduction is a form of simplification in science, the basing of population modeling on individual behavior is a step towards the consolidation and simplification of ecological theory.
 forecast that behavioral ecology and population ecology would be tightly interfaced by the end of the 20th century. Much of this interfacing, if it is to occur, will be accomplished through the extension of population models to incorporate the behavior and energetics of individual organisms in a realistic way. This will pave the way towards theory reduction, or interpreting the higher level phenomena of population dynamics in terms of lower level processes or mechanisms at the individual level (Schrader-Frechette and McCoy 1993). Because theory reduction is one of the ultimate goals of science, and because theory reduction is a form of simplification in science, the basing of population modeling on individual behavior is a step towards the consolidation and simplification of ecological theory..")
57
Ignore following slides

58
Level 1of IBM continuum: Discreteness and local interactions Examples: Smith (1991) Question: How do non-random strategies of predators and prey affect the stability of the predator prey interactions? If natural selection can occur, which searching rules will succeed? Model structure and variables: Landscape of L x L lattice cells, prey individuals, predator individuals Mechanisms : Prey individuals have probability of dividing that is inversely density dependent, probability of mortality on each time step. Predator individuals probability of capturing prey in their vicinity and are temporarily "satisfied" after capturing a prey item, reproduce with a probability dependent on recent prey capture, have a probability of dying that depends on recent prey capture. Movement: the majority of predators and prey initially moved randomly, but an initial 2-3% of predators followed one of six different movement strategies, such as movement of predators towards prey concentrations. Mutations in predator migration rates could occur, but more active migration diverted energy away from reproduction Results show movement rules have different success, and affect dynamics differently. Successful rules can eventually dominate.

59
IBM as a method of inquiry The situation to IBM in ecology is similar: The environment is complex and individual physiological and behavioral adaptations are also complex. Accurate modeling of populations and communities must take judicious account of this complexity. If general principles are to be found for ecological systems they must be founded on the adaptations of organisms to a complex and changing environment

60
Is a paradigmatic use of IBM emerging? To address still larger questions of ecology (e.g., biodiversity), CAS/IBM could be extended to larger systems of species. Simpler ( Level 1) IBM approaches (e.g., Pachepsksy et al. 2001) have been aimed at such questions. But it seems that the IBM approach becomes most interesting when individuals have the capability of altering behavior in response to complex environment. Is this something that can be done? Huse’s work suggests that it can be.
, CAS/IBM could be extended to larger systems of species. Simpler ( Level 1) IBM approaches (e.g., Pachepsksy et al. 2001) have been aimed at such questions. But it seems that the IBM approach becomes most interesting when individuals have the capability of altering behavior in response to complex environment. Is this something that can be done. Huse’s work suggests that it can be..")
Similar presentations

















 Sandi Merchant D-dudes meeting November 21, 2005.>")






