Download presentation
Presentation is loading. Please wait.

1
The Biochemistry of LTP Induction From Mechanisms of Memory by J. David Sweatt, Ph.D.

3
From Sheng and Kim

4
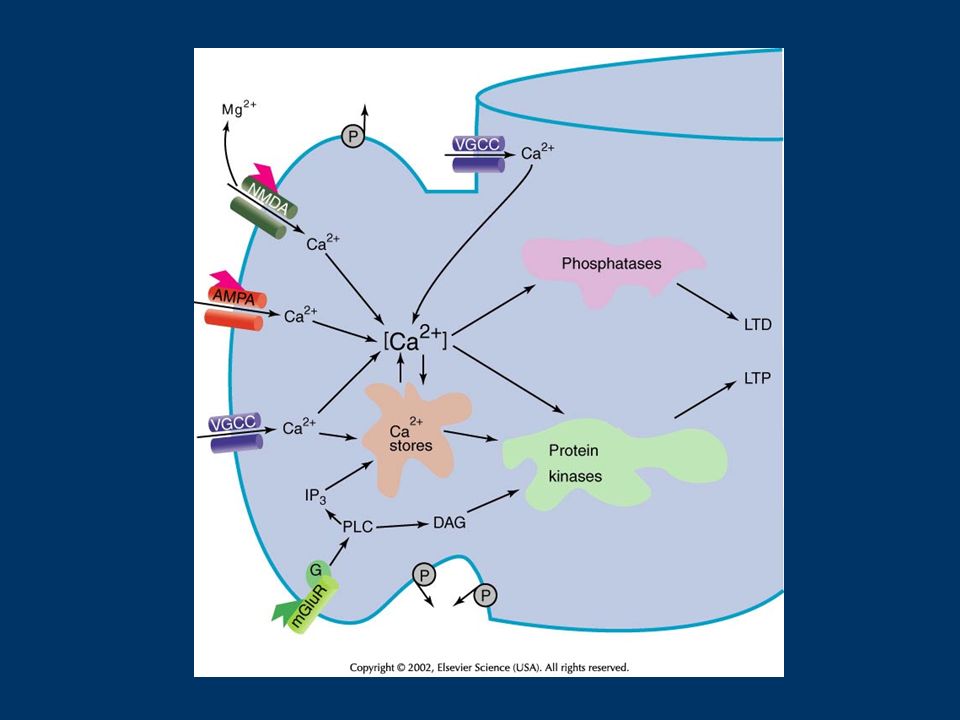
Neurotransmitter Receptor NMDA Receptor AMPA Receptor K Channels Ca ++ Channels IP 3 Receptor 1 2 23 3 4 4 5 LTP induction machinery Synaptic Infrastructure Ca ++ 6 Persisting Signal

5
The Biochemistry of LTP Induction 1.Mechanisms upstream of the NMDA receptor that directly regulate NMDA receptor function. 2. Mechanisms upstream of the NMDA receptor that control membrane depolarization. 3. The components of the synaptic infrastructure that are necessary for the NMDA receptor and the synaptic signal transduction machinery to function normally. 4. Feed-forward and feedback mechanisms that regulate the level of calcium attained. 5. Extrinsic signals that regulate the response to the calcium influx. 6. The mechanisms for the generation of the actual persisting biochemical signals.

7
Glutamate Receptors NR1120 NR2A180 NR2B180 GluR6 + 7117 mGluR1a200 Scaffolding and adaptors PSD-9595 ChapSyn110/PSD-93110 Sap102115 GKAP/SAPAP95-140 Shank200 Homer28/45 Yotiao200 AKAP150150 NSF83 PKA PKA catalytic subunit40 PKA-R2β53 PKC PKCβ80 PKCγ80 PKCε90 CaM Kinase CaM Kinase II β60 phosph-CaM Kinase60 Phosphatases PP136 PP2A36 PP2B(calcineurin)61 PPs50 PTPID/SHP272 Tyrosine Kinases Src60 PYK2116 MAP Kinase pathway ERK (pan ERK)42/44 ERK142/44 ERK242 MEK145 MEK246 MKP243 Rsk90 Rsk-290 c-Raf174 Small G-proteins and modulators Rac121 Rap221 SynGAP10,12,35,60 NF160,101 Other signaling molecules Calmodulin15 nNOS155 PI3 Kinase85 PLCγ130 cPLA2110 Citron183 Arg3.155 Cell adhesion and cytoskeletal proteins N-Cadherin150 Desmoglein165 β-Caternin92 LI200 pp120cas120 MAP2B280 Actin45 α-actinin 2110 Spectrin240/280 Myosin (brain)205 Tubulin50 Coractin80/85 CortBP-1180/200 Clathryn heavy chain180 Dynamin100 Hsp-7070 MoleculeMr (kD)MoleculeMr (kD)MoleculeMr (kD) Husi et al. (2001) Nature Neuroscience 3: 661-669.
 Nature Neuroscience 3:")
8
The Biochemistry of LTP Induction 1.Mechanisms upstream of the NMDA receptor that directly regulate NMDA receptor function. 2. Mechanisms upstream of the NMDA receptor that control membrane depolarization. 3. The components of the synaptic infrastructure that are necessary for the NMDA receptor and the synaptic signal transduction machinery to function normally. 4. Feed-forward and feedback mechanisms that regulate the level of calcium attained. 5. Extrinsic signals that regulate the response to the calcium influx. 6. The mechanisms for the generation of the actual persisting biochemical signals.

9
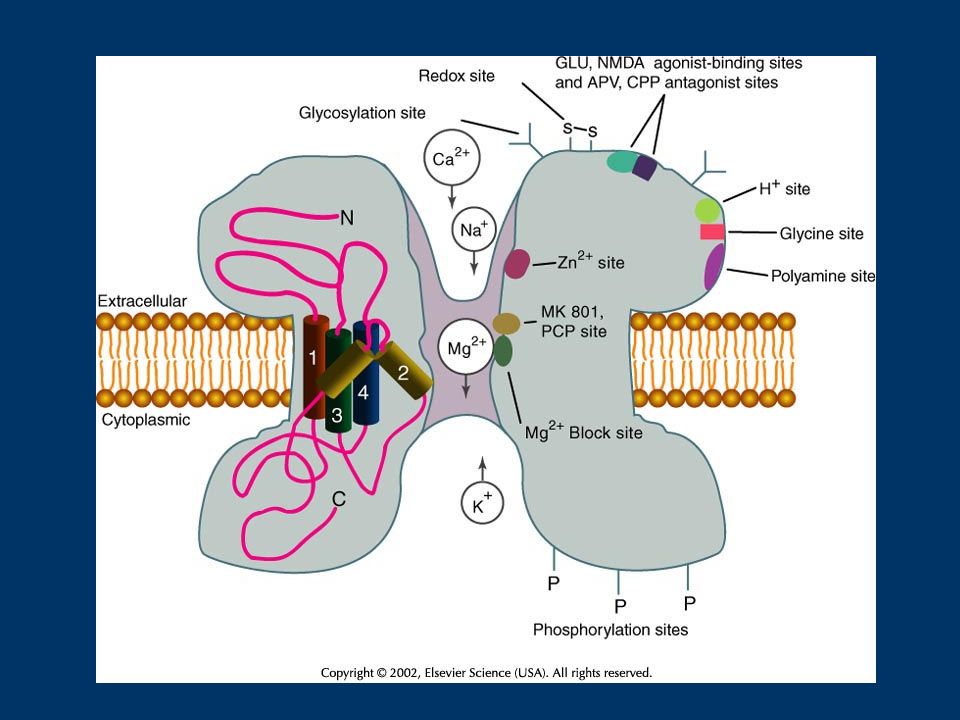
ModulatorMechanismEffect Src family tyrosine kinases (src, fyn)tyrosine phosphorylationenhancement loss of Zn inhibition Scaffolding proteins RACK1bindinginhibitory PSD-95scaffoldingmodulatory PKC ser/thr phosphoryation (direct)enhancement src activation (indirect) PKA/PP1/Yotiaophosphorylationenhancement dephosphorylationinhibition Cyclin dependent kinase 5ser/thr phosphorylationenhancement Nitric Oxide/redoxsulfhydryl nitrosylationinhibition or oxidation Polyamines (e.g. spermine, spermidine)direct binding to a modulatoryaugmentation site Caseine kinase IIser/thr phosphorylationenhancement modulation of polyamine effects TABLE I – DIRECT MODULATORS OF THE NMDA RECEPTOR
direct binding to a modulatoryaugmentation site Caseine kinase IIser/thr phosphorylationenhancement modulation of polyamine effects TABLE I – DIRECT MODULATORS OF THE NMDA RECEPTOR.")
10
Yotiao Leptin Receptor ApoE Receptor EphB Receptor NMDA Receptor Neurotransmitter Receptor Coupled To PLC Neurotransmitter Receptor Coupled To Acetyl Choline LeptinApoEEphrin B pyk2 ERK RACK Src/Fyn PSD95 Tyr PO 4 PI3K/MAPK ? ? ? Complex formation STEP PKC PL C PIP X Ser/Thr PO 4 PKA PP1 CDK5 CK II ATPcAMP ? DAG Receptor Modulation of the NMDA receptor

11
The Biochemistry of LTP Induction 1.Mechanisms upstream of the NMDA receptor that directly regulate NMDA receptor function. 2. Mechanisms upstream of the NMDA receptor that control membrane depolarization. 3. The components of the synaptic infrastructure that are necessary for the NMDA receptor and the synaptic signal transduction machinery to function normally. 4. Feed-forward and feedback mechanisms that regulate the level of calcium attained. 5. Extrinsic signals that regulate the response to the calcium influx. 6. The mechanisms for the generation of the actual persisting biochemical signals.

12
Ionic CurrentMolecules InvolvedRole Mechanisms of Modulation K Currents Voltage-dependentKv4.2 (and Kv4.3)limit bpAPsERK, PKA, CaMKII “A” currents limit EPSP magnitude “H” CurrentsNCN channelsregulate excitability cyclic nucleotides (direct) (HCN) Na Currents AMPA ReceptorsGluR1, GluR2depolarize membranePKA, CaMKII, PKC Aka GluR-A,B Voltage-dependentNa(v)1.6, 1.1,1.2AP propagation PKC (decreased inactivation) Na+ currents Ca Currents? – likely manyAP propagationPKA (hypothetical) Cl Currents GABA Receptorsall GABA-AAP firingnumerous receptor subunitsexcitability TABLE II – MECHANISMS UPSTREAM OF THE NMDA RECEPTOR INVOLVED IN MEMBRANE DEPOLARIZATION
 Cl Currents GABA Receptorsall GABA-AAP firingnumerous receptor subunitsexcitability TABLE II – MECHANISMS UPSTREAM OF THE NMDA RECEPTOR INVOLVED IN MEMBRANE DEPOLARIZATION.")
13
Three-way Coincidence Detection ↓ Kv4.2 Strong Input Back propagating Action Potential ACh CA1 Pyramidal Neuron NMDAR Glu 1 1 2 2 3

14
The Biochemistry of LTP Induction 1.Mechanisms upstream of the NMDA receptor that directly regulate NMDA receptor function. 2. Mechanisms upstream of the NMDA receptor that control membrane depolarization. 3. The components of the synaptic infrastructure that are necessary for the NMDA receptor and the synaptic signal transduction machinery to function normally. 4. Feed-forward and feedback mechanisms that regulate the level of calcium attained. 5. Extrinsic signals that regulate the response to the calcium influx. 6. The mechanisms for the generation of the actual persisting biochemical signals.

15
ComponentTargetsRole Cell Adhesion Molecules Integrinssrc, rho, rac, ras/MAPKsTransmembrane signaling, Interactions with extracellular matrix, NMDAR regulation MLCK, FAK?spine morphology? Syndecan-3fyn, NMDARsignaling from matrix heparan sulfates to the NMDA receptor N-Cadherinother Cadherins,spine morphology? cytoskeletonPre-post adhesion? Actin Cytoskeleton/Associated Proteins Rhomembrane/cytoskeletonregulate synaptic structure interactions Cdk5NMDA receptorincrease NMDA receptor function FilaminK channelsK channel localization Presynaptic Processes Glutamate releasesynaptic glutamateNMDA receptor activation Glutamate re-uptakesynaptic glutamatelimiting NMDA receptor desensitization TABLE III – COMPONENTS OF THE SYNAPTIC INFRASTRUCTURE NECESSARY FOR NMDA RECEPTOR FUNCTION

16
ComponentTargetsRole Anchoring/Interacting proteins PSD-95receptors,postsynaptic organization signal transduction mechs nNOS, SynGAP, GKAP NMDA receptormultiple proteinseffector localization, structural organization Rack1/fynNMDA receptor direct regulation of NMDA receptor Shank/HOMERmetabotropic receptorseffector localization, cytoskeleton GRIPAMPA receptors,postsynaptic organization PICK-1/PKC AKAPPKA, PP2B kinase and phosphatase localization CaMKIIsignal transduction regulate likelihood of LTP induction TABLE III – COMPONENTS OF THE SYNAPTIC INFRASTRUCTURE NECESSARY FOR NMDA RECEPTOR FUNCTION ( Continued)
")
17
NMDAR NR2 NMDAR NR2 AMPAR GluR2,GluR3 AMPAR GAP PSD-95 rap actin n-NOS SynGAP GKAP PSD95 GKAP Shank Homer IP 3 R PLC actin ras IP 3 + DAG SPAR cortactin - Spectrin PICK-1 PKC GRIP NSF GRASP1 (GEF for ras) ras PKAPKC AKAP79PP2B SAP97 CamKII β-AR Receptor Trafficking liprin Group I mGluR PSD-95 as an Anchoring Protein for NMDA Receptors
 ras PKAPKC AKAP79PP2B SAP97 CamKII β-AR Receptor Trafficking liprin Group I mGluR PSD-95 as an Anchoring Protein for NMDA Receptors")
18
From Sheng and Kim

23
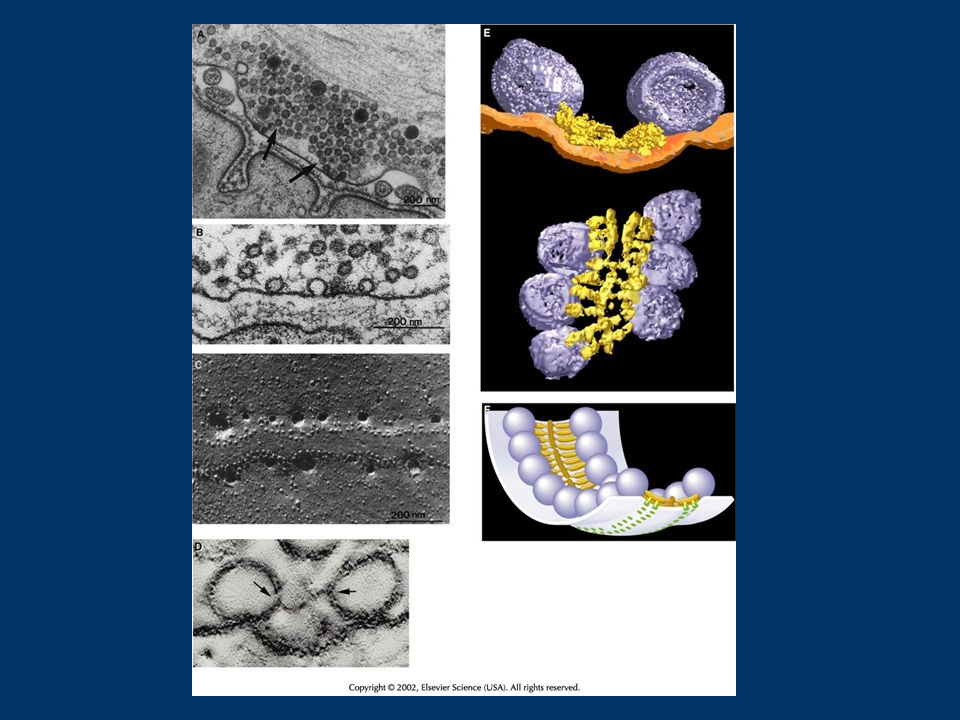
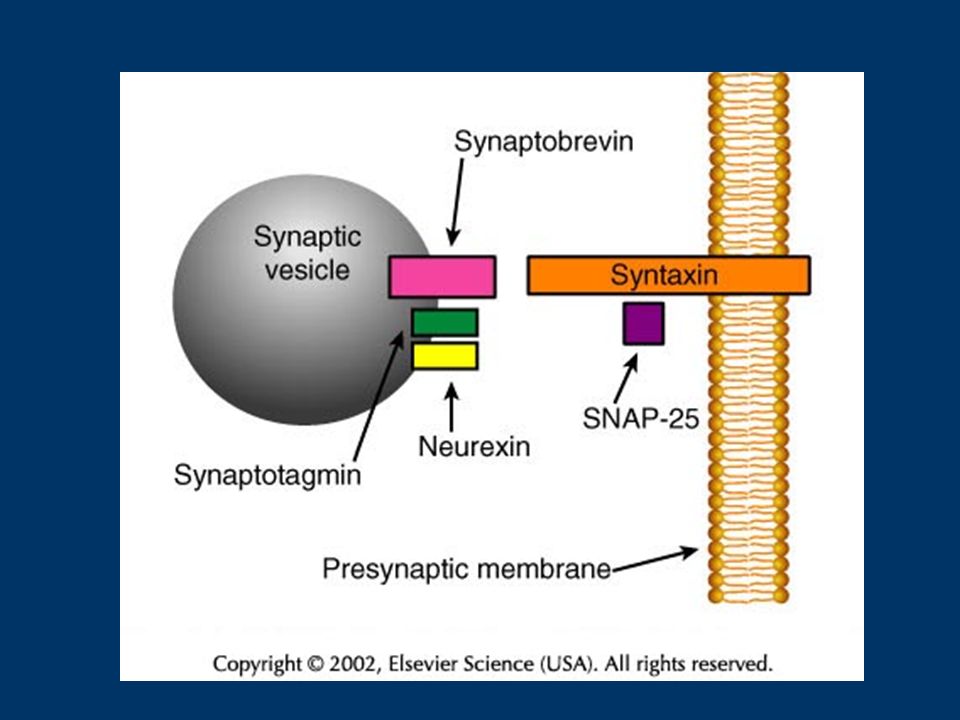
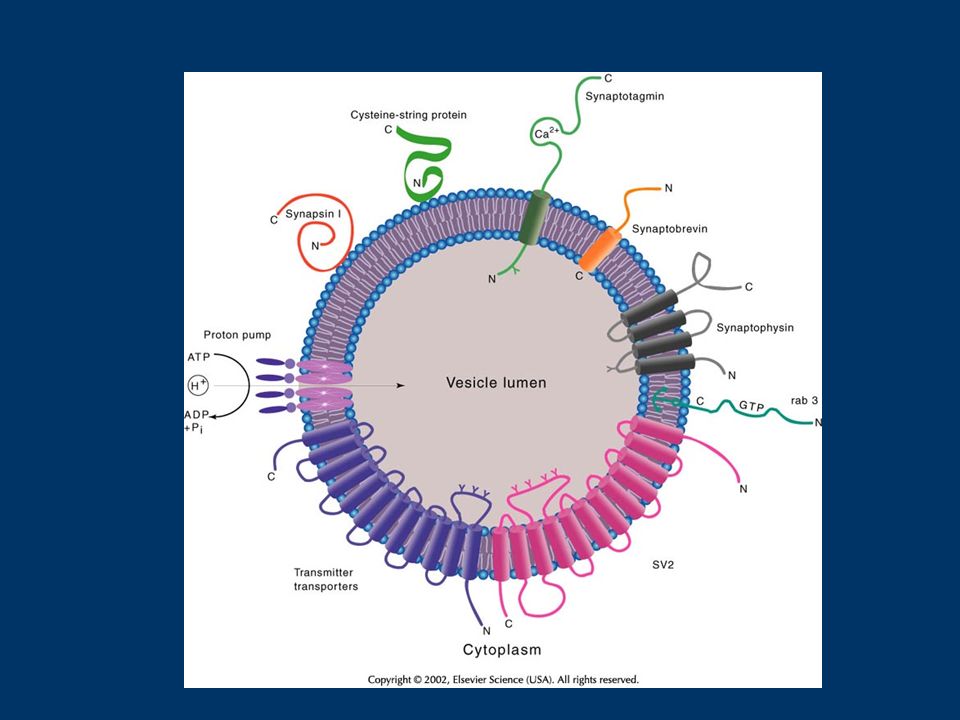
Fig. 1. RIM1 and the priming of synaptic vesicle fusion. (a) After docking, synaptic vesicles (SV) are tethered at the active zone by binding of Rab3 to the N-terminal (N) of RIM1 (Rab3-interactive molecule-1). Munc-13 is recruited to the active zone by activity of phospholipase C (PLC) and the second messenger diacylglycerol (DAG). Munc-18 binding to syntaxin (Syntx) keeps syntaxin in a `closed' conformation that cannot bind SNAP-25 (synapstosome-associated protein-25). (b) Activation of second- messenger pathways – such as those involving Ca2+, adenylate cyclase (AC), cAMP and protein kinase A (PKA) – during induction of short-term plasticity leads to a switch in the binding partners of RIM1. Munc-13-1 binds to N-terminal RIM1, competitively inhibiting the binding of Rab3 to RIM1. Thus, a new tethering mechanism holds the SVs at the active zone, as synaptotagmin1/2 (Synat) binds to the C-terminal RIM domains in a Ca2+-dependent manner. Binding of munc-13 to syntaxin removes munc-18 and converts syntaxin's structure to an open conformation. (c) Proximity of synaptotagmin to the plasma membrane, conversion of syntaxin by Munc-13-1 to an open conformation that can interact with SNAP-25, and further increase in cytoplasmic free Ca2+ levels, promote the formation of the synaptobrevin (Syb)–syntaxin–SNAP-25 complex that is required for fusion.
 After docking, synaptic vesicles (SV) are tethered at the active zone by binding of Rab3 to the N-terminal (N) of RIM1 (Rab3-interactive molecule-1). Munc-13 is recruited to the active zone by activity of phospholipase C (PLC) and the second messenger diacylglycerol (DAG). Munc-18 binding to syntaxin (Syntx) keeps syntaxin in a `closed conformation that cannot bind SNAP-25 (synapstosome-associated protein-25). (b) Activation of second- messenger pathways – such as those involving Ca2+, adenylate cyclase (AC), cAMP and protein kinase A (PKA) – during induction of short-term plasticity leads to a switch in the binding partners of RIM1. Munc-13-1 binds to N-terminal RIM1, competitively inhibiting the binding of Rab3 to RIM1. Thus, a new tethering mechanism holds the SVs at the active zone, as synaptotagmin1/2 (Synat) binds to the C-terminal RIM domains in a Ca2+-dependent manner. Binding of munc-13 to syntaxin removes munc-18 and converts syntaxin s structure to an open conformation. (c) Proximity of synaptotagmin to the plasma membrane, conversion of syntaxin by Munc-13-1 to an open conformation that can interact with SNAP-25, and further increase in cytoplasmic free Ca2+ levels, promote the formation of the synaptobrevin (Syb)–syntaxin–SNAP-25 complex that is required for fusion..")
25
Three Pools of F-Actin in Synaptic Spines The upper panels are single computed slices through electron tomographic volumes of spines labeled for F- actin using phaloidin-eosin photo conversion, from hippocampus CA1 (A) and cerebellar cortex molecular layer (B) (see Capani et al., 2001 ). Labeling is concentrated between the lamellae of the spine apparatus (SA) and the postsynaptic density (arrowheads). Bundles of actin are seen traversing between these entities (large arrow). In Purkinje cells, which have no spine apparatus, actin filaments fill the head and also can be followed between the smooth ER and the postsynaptic membrane (large arrow). Diffuse staining for actin is also seen (asterisks). The stereo computer graphic reconstruction in the bottom panel is of the CA1 synapse and shows actin bundles (blue) as well as the spine apparatus (yellow) and the postsynaptic density (purple). These figures were kindly provided by Dr. Mark Ellisman.
 and the postsynaptic density (arrowheads). Bundles of actin are seen traversing between these entities (large arrow). In Purkinje cells, which have no spine apparatus, actin filaments fill the head and also can be followed between the smooth ER and the postsynaptic membrane (large arrow). Diffuse staining for actin is also seen (asterisks). The stereo computer graphic reconstruction in the bottom panel is of the CA1 synapse and shows actin bundles (blue) as well as the spine apparatus (yellow) and the postsynaptic density (purple). These figures were kindly provided by Dr. Mark Ellisman..")
26
Figure 1. LIMK Influences Postsynaptic and Presynaptic Function through Modulation of Actin Filaments Dendritic spines are made up of a head, neck, and postsynaptic density (PSD). Within the PSD, scaffold proteins such as Homer, PSD-95, and Shank, as well as others not described here, link the actin cytoskeleton to postsynaptic receptors including AMPA and NMDA glutamate receptors. Results in this issue of Neuron by Meng et al. (2002 ) demonstrate that LIMK-1 is partially responsible for proper dendritic morphology and long-term potentiation (LTP), presumably via its effect on actin filament dynamics, through phosphorylation and inactivation of ADF/cofilin (AC). In LIMK-1−/− mice, the morphology of dendritic spines is altered. The spines have a thicker neck and smaller postsynaptic density length and smaller spine area. Results presented by Meng et al. (2002 ) also reveal that the LIMK-1−/− mice have enhanced basal release of presynaptic vesicles and an enhanced synaptic depression, suggesting a role for LIMK-1 (and most likely actin dynamics) in neurotransmitter release. Figure by Patrick D. Sarmiere and James R. Bamburg
. Within the PSD, scaffold proteins such as Homer, PSD-95, and Shank, as well as others not described here, link the actin cytoskeleton to postsynaptic receptors including AMPA and NMDA glutamate receptors. Results in this issue of Neuron by Meng et al. (2002 ) demonstrate that LIMK-1 is partially responsible for proper dendritic morphology and long-term potentiation (LTP), presumably via its effect on actin filament dynamics, through phosphorylation and inactivation of ADF/cofilin (AC). In LIMK-1−/− mice, the morphology of dendritic spines is altered. The spines have a thicker neck and smaller postsynaptic density length and smaller spine area. Results presented by Meng et al. (2002 ) also reveal that the LIMK-1−/− mice have enhanced basal release of presynaptic vesicles and an enhanced synaptic depression, suggesting a role for LIMK-1 (and most likely actin dynamics) in neurotransmitter release. Figure by Patrick D. Sarmiere and James R. Bamburg.")
27
Presynaptic Postsynaptic NMDA Receptor Retrograde Signaling Integrins rho rac FAK MLCK ras α-actinin Src/fyn ERK β subunit filamin ? ? cdk5 talin vinculin ? Kv4.2 Channel actin Dynamic Regulation Integrins Extracellular Matrix Interactions among Integrins and Intracellular Effectors

28
The Biochemistry of LTP Induction 1.Mechanisms upstream of the NMDA receptor that directly regulate NMDA receptor function. 2. Mechanisms upstream of the NMDA receptor that control membrane depolarization. 3. The components of the synaptic infrastructure that are necessary for the NMDA receptor and the synaptic signal transduction machinery to function normally. 4. Feed-forward and feedback mechanisms that regulate the level of calcium attained. 5. Extrinsic signals that regulate the response to the calcium influx. 6. The mechanisms for the generation of the actual persisting biochemical signals.

29
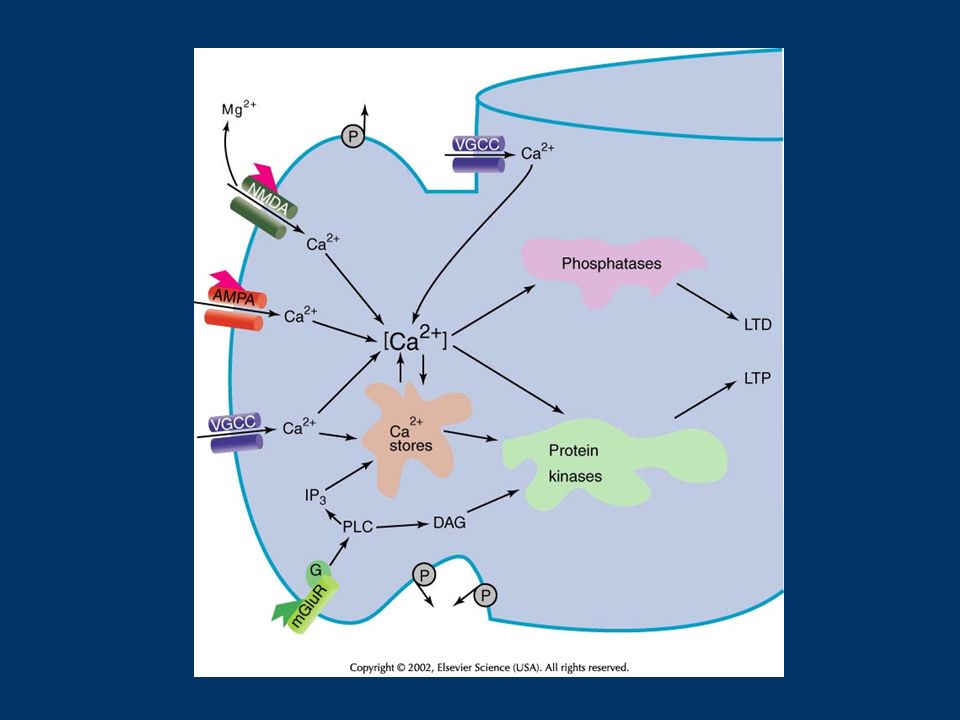
Molecule/OrganelleRoleModulator/Regulator VDCCsaugment NMDAR-dependentPKA Ca influx Ca influx due to bpAPs regulate ERK activation Endoplasmic ReticulumCa efflux from ER, limit LTP?PLC-coupled receptors (Ca ATPase/IP3R/RyR) Presynaptic Mitochondriaregulate presynaptic Ca levelsunknown TABLE IV – CALCIUM FEEDBACK AND FEED-FORWARD MECHANISMS
 Presynaptic Mitochondriaregulate presynaptic Ca levelsunknown TABLE IV – CALCIUM FEEDBACK AND FEED-FORWARD MECHANISMS")
31
The Biochemistry of LTP Induction 1.Mechanisms upstream of the NMDA receptor that directly regulate NMDA receptor function. 2. Mechanisms upstream of the NMDA receptor that control membrane depolarization. 3. The components of the synaptic infrastructure that are necessary for the NMDA receptor and the synaptic signal transduction machinery to function normally. 4. Feed-forward and feedback mechanisms that regulate the level of calcium attained. 5. Extrinsic signals that regulate the response to the calcium influx. 6. The mechanisms for the generation of the actual persisting biochemical signals.

32
Regulatory SystemMolecules InvolvedRole The cAMP GatePKA/PP1/I1/PP2BPhosphatase Inhibition Augmented Kinase Signaling The PKC/NeurograninPLC/PKC/Neurogranin/CaMAugmenting CaMKII Activation System Augmenting Ca-sensitive Cyclase TABLE V – EXTRINSIC SIGNALS MODULATING THE CALCIUM RESPONSE

33
Model for the cAMP Gate Sweatt (2001) Curr. Biol. 11:R391-394.
 Curr. Biol. 11:R")
34
Phospholipase C Neurogranin PO 4 + DAG PKC Calmodulin Metabotropic Receptor PKC Phosphorylation of Neurogranin

35
Augmented PKC cAMP GATE Initial Ca++ Signal Increased Ca++/CaM Augmented CaMKII Activity Adenylyl Cyclase NMDAR Neurogranin DAG Cyclase Coupled Receptors Metabotropic Receptors The PKC/Neurogranin system and the cAMP Gate

36
The Biochemistry of LTP Induction 1.Mechanisms upstream of the NMDA receptor that directly regulate NMDA receptor function. 2. Mechanisms upstream of the NMDA receptor that control membrane depolarization. 3. The components of the synaptic infrastructure that are necessary for the NMDA receptor and the synaptic signal transduction machinery to function normally. 4. Feed-forward and feedback mechanisms that regulate the level of calcium attained. 5. Extrinsic signals that regulate the response to the calcium influx. 6. The mechanisms for the generation of the actual persisting biochemical signals.

37
Four-way Coincidence Detection ↓ Kv4.2 Strong Input Back propagating Action Potential ACh CA1 Pyramidal Neuron NMDAR Glu 1 1 2 2 3 cAMP GATE Norepinephrine 4 4

38
The Biochemistry of LTP Induction From Mechanisms of Memory by J. David Sweatt, Ph.D.

Similar presentations









 Ian Parker Lecture # 6 - Second messenger and Ca 2+ signaling.>")
 limited response –depolarization.>")









