Download presentation
Presentation is loading. Please wait.

1
Chapter22 Population Genetics
Individuals can carry only two different alleles of a given gene. A group of individuals can carry a large number of different alleles, give rise to a reservoir of genetic diversity. Diversity contained in the population can be measured by the Hardy-Weinberg law. Mutation is the ultimate source of genetic variation and other factors such as drift, migration, and selection can alter the amount of genetic variation in population

2
22.1 Population and gene pool
22.2 Calculating allele frequency 22.3 The Hardy-Weinberg law 22.4 Extension of the Hardy-Weinberg law 22.5 Using the Hardy-Weinberg law, calculating heterozygote frequency 22.6 Factors that later allele frequency in population 22.7 Natural selection 22.8 Mutation 22.9 Migration 22.10 Genetic drift 22.11 Nonrandom mating

3
Alfred Russell Wallace and Charles Darwin first identified natural selection as the mechanism of adaptive evolution in the mid-nineteenth century, based on a series of observations of populations of organisms: (1) Phenotypic variations exist among individuals within populations; (2) these differences are passed from parents to offspring; (3) more offspring are born than will survive and reproduce; and (4) some variants are more successful at surviving and/or reproducing than others. In populations where all four factors operate, the relative abundance of the population's different phenotypes changes across generations. In other words, the population evolves.
 Phenotypic variations exist among individuals within populations; (2) these differences are passed from parents to offspring; (3) more offspring are born than will survive and reproduce; and. (4) some variants are more successful at surviving and/or reproducing than others. In populations where all four factors operate, the relative abundance of the population s different phenotypes changes across generations. In other words, the population evolves.")
4
Although Wallace and Darwin described how organisms evolve by natural selection, there was no accurate model of the mechanisms responsible for variation and inheritance. Gregor Mendel published his work on the inheritance of traits in 1865, For many years, theorists focused on developing mathematical models that would describe the genetic structure of populations. Prominent among the theoreticians who developed these models were Sewall Wright, Ronald Fisher, and J. B. S. Haldane. Following their work, experimentalists and field workers tested the models using biochemical and molecular techniques that measure variation directly at the protein and DNA levels. These experiments examined allele frequencies and the forces that alter the frequencies, such as selection, mutation, migration, and random genetic drift. In this chapter we consider some general aspects of population genetics and also discuss other areas of genetics that relate to evolution.

5
22.1 Populations and Gene Pools
Members of a species often range over a wide geographic area. A population is a group of individuals from the same species that lives in the same geographic area(孟德尔群体 ), and that actually or potentially interbreeds. If we consider a single genetic locus in this population, we may find that individuals within the population have different genotypes. To study population genetics, we compute frequencies at which various alleles and genotypes occur, and how these frequencies change from one generation to the next.
, and that actually or potentially interbreeds. If we consider a single genetic locus in this population, we may find that individuals within the population have different genotypes. To study population genetics, we compute frequencies at which various alleles and genotypes occur, and how these frequencies change from one generation to the next.")
6
When we consider generational changes in alleles and genotypes, we look at gene pools.
A gene pool consists of all gametes made by all the breeding members of a population in a single generation Gene pool基因库 将群体中所有个体共有的全部基因定义称为一个基因库(gene pool)。 : gametes to zygotes - next generation. alleles proportion of gametes in a gene pool
。 : gametes to zygotes - next generation. alleles. proportion of gametes in a gene pool.")
7
22.2 Calculating Allele Frequencies
Most genetic population researchers first measure the frequencies at which alleles occur at a particular locus. To do this, the genotypes of a large number of individuals in the population must be determined. In some cases, researchers can infer genotypes directly from phenotypes. In other cases, proteins or DNA sequences are analyzed to determine genotypes. To understand how allele frequencies are calculated, we consider an example that involves HIV infection rates.

8
In 1996, Rong Liu and colleagues discovered that two exposed-but-uninfected individuals were homozygous for a mutant allele of a gene called CC-CKR-5. The CC-CKR-5 gene, chromosome 3, a protein called the C-C chemokine(趋化因子) receptor-5(CCR5). Chemokines are cell-surface signaling molecules found on cells in the immune system. When cells with CCR5 receptor proteins bind to chemokine signals, the cells move into inflamed tissues to fight infection. The CCR5 protein is a co-receptor for strains of HIV-1. To gain entry into cells, a protein called Env (short for envelope protein) on the surface of HIV-1 binds to the CD4 protein on the surface of the host cell. Binding to CD4 causes Env to change shape and form a second binding site. This second site binds to CCR5, which in turn initiates the fusion of the viral protein coat with the host cell membrane. The merging of viral envelope with cell membrane transports the HIV viral core into the host cell's cytoplasm.
 receptor-5(CCR5). Chemokines are cell-surface signaling molecules found on cells in the immune system. When cells with CCR5 receptor proteins bind to chemokine signals, the cells move into inflamed tissues to fight infection. The CCR5 protein is a co-receptor for strains of HIV-1. To gain entry into cells, a protein called Env (short for envelope protein) on the surface of HIV-1 binds to the CD4 protein on the surface of the host cell. Binding to CD4 causes Env to change shape and form a second binding site. This second site binds to CCR5, which in turn initiates the fusion of the viral protein coat with the host cell membrane. The merging of viral envelope with cell membrane transports the HIV viral core into the host cell s cytoplasm.")
9
The mutant allele of the CCR5 gene contains a 32-bp deletion in one of its coding regions. As a result, the protein encoded by the mutant allele is shortened and made nonfunctional. The protein never makes it to the cell membrane, so HIV-1 cannot enter these cells. The gene's normal allele is called CCR51 (or 1) and its allele with the 32-bp deletion is called CCK5- 32 (or 32), The two uninfected individuals described by Liu both had the genotype 32/ 32. As a result, they had no CCR5 on the surface of their cells, and were resistant to infection by strains of HIV-1 that require CCR5 as a co-receptor.

10
Table 22.1 CCRS genotype and phenotype
At least three 32/32 individuals who are infected with HIV-1 have been found. Curiously, researchers have not discovered any adverse effects associated with this genotype. Heterozygotes with genotype 1/32 are susceptible to HIV-1 infection, but evidence suggests that they progress more slowly to AIDS. Table 22.1 summarizes the genotypes possible at the CCRS locus, and the phenotypes associated with each. Table 22.1 CCRS genotype and phenotype 1/ susceptible to sexually transmitted HIV-1 strain 1/ susceptible, but may progress to AIDS slowly 32/ 32 resistant to most sexually transmitted HIV-1s

11
The discovery of the CCR5- 32 allele, and the fact that it provides some protection against AIDS, generates two important questions: Which human populations harbor the 32 allele, and how common is it? To address these questions, several teams of researchers surveyed a large number of people from a variety of populations. Genotypes were determined by direct analysis of DNA (Figure 22-1).
..")
13
Figure 22-2 shows the frequency of the CCR5-A32 allele in the 18 European populations surveyed. The studies show that populations in Northern Europe have the highest frequencies of the A32 allele. The frequency of the allele declines to the south and to the west. In populations without European ancestry the A32 allele is essentially absent. The highly patterned global distribution of the A32 allele presents an evolutionary puzzle that we'll return to later in the chapter

14
22.3 The Hardy-Weinberg Law
The large variation in the frequency of the CCR5-A32 allele among populations raises a number of questions. For example, can we expect the allele to increase in populations in which it is currently rare? Population genetics explores such questions using a mathematical model developed independently by the British mathematician Godfrey H. Hardy and the German physician Wilhelm Weinberg. This model, called the Hardy-Weinberg law, shows what happens to allele and genotype in an "ideal" population (free of many of the complications that affect real populations) using a set of simple assumptions.
 using a set of simple assumptions.")
15
1. Individuals of all genotypes have equal rates of survival and equal reproductive success; that is, there is no selection. 2. No new alleles are created or converted from one allele into another by mutation. 3. Individuals do not migrate into or out of the population. 4. The population is infinitely large, which in practical terms means that the population is large enough that mate randomly sampling errors and other random effects are negligible. 5.Individuals in the population mate randomly

16
The Hardy-Weinberg law demonstrates that an "ideal" population has these properties:
1. The frequency of alleles does not change from generation to generation; in other words, the population does not evolve. 2. After one generation of random mating, offspring genotype frequencies can be predicted from the parent allele frequencies. What makes the Hardy-Weinberg law useful, however, is its assumptions. By specifying the assumptions under which the population cannot evolve, the Hardy-Weinberg law identifies the real-world forces that cause allele frequencies to change. In other words, by holding certain conditions constant, the Hardy-Weinberg law isolates the forces of evolution.

17
A Demonstration of the Law
To demonstrate how the Hardy-Weinberg law works, we begin with a specific case, and then consider the general case. In both examples, we focus on a single locus with two alleles, A and a. Imagine a population in which the frequency of allele A, in both eggs and sperm, is 0.7, a is 0.3, Note that = 1, indicating that all the alleles for gene A present in the gene pool are accounted for. We assume, per Hardy-Weinberg requirements, that individuals mate randomly, which we visualize as follows. We place all the gametes in the gene pool, in a barrel and stir. We then randomly draw eggs and sperm from the barrel and pair them to make zygotes. What genotype frequencies does this give us?

18
The probability of genotype AA will occur 49%
Aa is = 0.42 = 42 % aa is 9 %

19
We started with the frequency of a particular allele in a specific gene pool and calculated the probability that certain genotypes would be produced from this pool. When the zygotes develop into adults and reproduce, what will be the frequency of distribution of alleles in the new gene pool? Recall that under the Hardy-Weinberg law, we assume that all genotypes have equal rates of survival and reproduction. This means that in the next generation, all genotypes contribute equally to the new gene pool.

20
The AA individuals 49% carry allele A,
Likewise, Aa individuals 42% Half (0.5) of these gametes carry allele A. Frequency of allele A in the gene pool is (0.5) 0.42 = 0.7. The other half carry allele a. The aa individuals 9 % (0.5) = 0.3. As a check on our calculation, note that = 1.0, accounting for all of the gametes in the gene pool of the new generation.
 of these gametes carry allele A. Frequency of allele A in the gene pool is (0.5) 0.42 = 0.7. The other half carry allele a. The aa individuals 9 % (0.5) = 0.3. As a check on our calculation, note that = 1.0, accounting for all of the gametes in the gene pool of the new generation.")
21
We have arrived back where we began, with a gene pool in which the frequency of allele A is 0.7 and the frequency of allele a is 0.3. These calculations demonstrate the Hardy-Weinberg law: Allele frequencies in our population do not change from one generation to the next, and after just one generation of random mating the genotype frequencies can be predicted from the allele frequencies. In other words, this population does not evolve. use variables instead of, numerical values for the allele frequencies. A - p a - q, p + q = 1. Zygote that carry allele A is p X p. AA = p2. Aa=2pq. aa=q2

22
Figure 22-4 shows these calculations
p2 + 2pq + q2 = 1

23
For our general case we ask what the allele frequencies in the new gene pool will be when these zygotes develop into adults and reproduce. All gametes from AA individuals carry allele A, as do half of the gametes from Aa individuals. Thus we predict that the frequency of allele A in the new gene pool will be p2 + (1/2) 2pq = p2 + pq Or P2 + p(l ~ p) = p2 + P - P2 = P Likewise, the frequency of allele a in the new gene pool will be (1/2)2pq + q2 = pq + q2 Or (1 - q)q + q2 = q - q2 + q2 = q
 2pq = p2 + pq. Or P2 + p(l ~ p) = p2 + P - P2 = P. Likewise, the frequency of allele a in the new gene pool will be. (1/2)2pq + q2 = pq + q2. Or (1 - q)q + q2 = q - q2 + q2 = q.")
24
Consequences of the Law
The Hardy-Weinberg law has several important consequences. First, it shows that dominant traits do not necessarily increase from one generation to the next. Second, it demonstrates that genetic variability can be maintained in a population since, once established in an ideal population, allele frequencies remain unchanged. Third, if we invoke Hardy-Weinberg assumptions, then knowing the frequency of just one genotype enables us to calculate the frequencies of all other genotypes. This relationship is particularly useful in human genetics because we can now calculate the frequency of heterozygous carriers for recessive genetic disorders even when all we know is the frequency of affected individuals

25
We began this discussion by asking whether we can expect the CCR5-32 allele to increase in populations in which it is currently rare. From what we now know about the Hardy-Weinberg law, we can say for the general case that if (l) individuals of all genotypes have equal rates of survival and reproduction, (2) there is no mutation, (3) no one migrates into or out of the population, (4) the population is extremely large, (5) individuals in the population choose their mates randomly, then the frequency of the 32 allele will not change. Of course, for real populations, few if any of these assumptions are likely to hold.
 individuals of all genotypes have equal rates of survival and reproduction, (2) there is no mutation, (3) no one migrates into or out of the population, (4) the population is extremely large, (5) individuals in the population choose their mates randomly, then the frequency of the 32 allele will not change. Of course, for real populations, few if any of these assumptions are likely to hold.")
26
The general case demonstrates the most important role of the Hardy-Weinberg law: It is the foundation upon which population genetics is built. By showing that "ideal" populations do not evolve, we can use the Hardy-Weinberg law to identify forces that do cause populations to evolve.

27
when the assumptions of the Hardy-Weinberg law are broken—-
because of natural selection, mutation, migration, and random sampling errors (also known as genetic drift)—the allele frequencies in a population may change from one generation to the next. Nonrandom mating does not, by itself, alter allele frequencies, but by altering genotype frequencies it indirectly affects the course of evolution. The Hardy-Weinberg law tells geneticists where to look to find the causes of evolution in real populations. We return to the CCR5-A32 allele later in the chapter to see how a population genetics perspective has generated fruitful hypotheses for further research.
—the allele frequencies in a population may change from one generation to the next. Nonrandom mating does not, by itself, alter allele frequencies, but by altering genotype frequencies it indirectly affects the course of evolution. The Hardy-Weinberg law tells geneticists where to look to find the causes of evolution in real populations. We return to the CCR5-A32 allele later in the chapter to see how a population genetics perspective has generated fruitful hypotheses for further research.")
28
Testing for Equilibrium
One way we establish whether one or more of the Hardy-Weinberg assumptions do not hold in a given population is by determining whether the population's genotypes are in equilibrium. To do this, we first determine the frequencies of the genotypes, either directly from the phenotypes (if heterozygotes are recognizable) or by analyzing proteins or DNA sequences. We then calculate the allele frequencies from the genotype frequencies, as demonstrated earlier. Finally, we use the parents' allele frequencies to predict the offspring's genotype frequencies. According to the Hardy-Weinberg law, the genotype frequencies are predicted to fit the p2 + 2pq + q2 = 1 relationship. If they do not, then one or more of the assumptions are invalid for the population in question.
 or by analyzing proteins or DNA sequences. We then calculate the allele frequencies from the genotype frequencies, as demonstrated earlier. Finally, we use the parents allele frequencies to predict the offspring s genotype frequencies. According to the Hardy-Weinberg law, the genotype frequencies are predicted to fit the p2 + 2pq + q2 = 1 relationship. If they do not, then one or more of the assumptions are invalid for the population in question.")
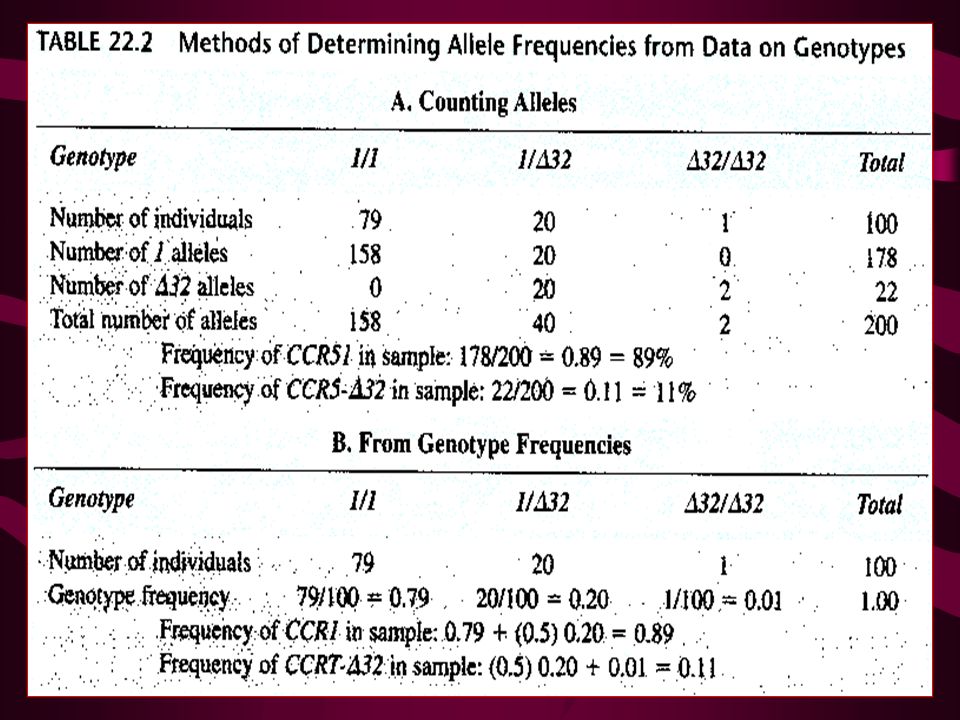
29
We will use the CCR5 genotypes of a population in Britain to demonstrate the Hardy-Weinberg law. The population includes 283 individuals, of which 223 have genotype 1/1, 57 have genotype 1/A32, and 3 have genotype A32/A32. These numbers represent genotype frequencies of 223/283 = 0.788, 57/283 = 0.201, and 3/283 = 0.011, respectively. From the genotype frequencies we compute the CCR5I allele frequency as 0.89 and the frequency of the CCR5-A32 allele as From these allele frequencies, we can use the Hardy-Weinberg law to determine whether this population is in equilibrium. The allele frequencies predict the genotype frequencies as follows: Expected frequency of genotype 1/1 = p2 = (0.89) Exp genotype 1/A32 - 2pq = 2(0.89)(0.11) = 0.196 Expected genotype A32IA32 = q2 = (0.11 )2 = 0.012
 Exp genotype 1/A32 - 2pq = 2(0.89)(0.11) = Expected genotype A32IA32 = q2 = (0.11 )2 =")
30
These expected frequencies are nearly identical to the observed frequencies. Our test of this population has failed to provide evidence that Hardy-Weinberg assumptions are being violated. This conclusion is confirmed by a X2 analysis (see Chapter 3). The X2 value in this case is tiny: To be statistically significant at even the most generous, accepted level, p = 0.05, the X2 value would have to be (In a test for Hardy-Weinberg equilibrium, the degrees of freedom are given by k m, where k is the number of genotypes and m is the number of independent allele frequencies estimated from the data.

31
On the other hand, if the Hardy-Weinberg test had demonstrated that the population is not in equilibrium, it would indicate that one or more assumptions are not being met. To illustrate this, imagine two hypothetical populations, one living on East Island,all 1/1, 100 % other living on West Island, A32/A32, 100 % Now imagine that 500 people from each island move to the previously uninhabited Central Island.

34
However, it would take only one generation of random mating on Central Island to bring the offspring to the expected allele frequencies, as shown in Figure Therese Markow and colleagues documented a real human population that is not in Hardy-Weinberg equilibrium. These researchers studied 122 Havasupai, a population of Native Americans in Arizona. They determined the genotype of each Havasupai individual at two loci in the major histocompatibility complex (MHC). These genes, HLA-A and HLA-8, encode proteins that are involved in the immune system's discrimination between self and nonself. The immune systems of individuals heterozygous at MHC loci appear to recognize a greater diversity of foreign invaders and thus may be better able to fight disease.
. These genes, HLA-A and HLA-8, encode proteins that are involved in the immune system s discrimination between self and nonself. The immune systems of individuals heterozygous at MHC loci appear to recognize a greater diversity of foreign invaders and thus may be better able to fight disease..")
35
Markow and colleagues observed significantly more individuals who are heterozygous at both loci, and significantly fewer homozygous individuals than would be expected under the Hardy-Weinberg law. Violation of either or both of two Hardy-Weinberg assumptions could explain the excess of heterozygotes among the Havasupai. First, Havasupai fetuses, children, and adults who are heterozygous for HLA-A and HLA-B may have higher rates of survival than individuals who are homozygous. Second, rather than choosing their males randomly, Havasupai people may somehow prefer mates whose MHC genotypes differ from their own.

36
22.4 Extensions of the Hardy-Weinberg Law
We commonly find several alleles of a single locus in a population. The ABO blood group in humans (discussed in Chapter 4) is such an example. The locus I (isoagglutinin) has three alleles (IA, IB, and IO), 6 genotypic A and B codominant, both dominant to O. AA , AO phenotypic identical, BB and BO ” only 4 distinguished Let p, q, and r represent A, B, and 0, respectively. p + q + r = 1
 is such an example. The locus I (isoagglutinin) has three alleles (IA, IB, and IO), 6 genotypic. A and B codominant, both dominant to O. AA , AO phenotypic identical, BB and BO only 4 distinguished. Let p, q, and r represent A, B, and 0, respectively. p + q + r = 1.")
37
Under Hardy-Weinberg assumptions, the frequencies of the genotypes are given by
(P + q + r)2 =p2 + q2+r2 + 2pq + 2r + 2qr = 1 If we know the frequencies of blood types for a population, we can then estimate the frequencies for the three alleles of the ABO system. E.g., in one population sampled, A = 0.53, B = 0.13,AB , and O = Because the O allele is recessive, the population's frequency of type O blood equals the proportion of the recessive genotype r2. Thus, r2 = ,r = √ =.51
2 =p2 + q2+r2 + 2pq + 2r + 2qr = 1. If we know the frequencies of blood types for a population, we can then estimate the frequencies for the three alleles of the ABO system. E.g., in one population sampled, A = 0.53, B = 0.13,AB , and O = Because the O allele is recessive, the population s frequency of type O blood equals the proportion of the recessive genotype r2. Thus, r2 = 0.26 ,r = √.26 =.51.")
38
Using r, we can estimate the allele frequencies for the A and B alleles. The A allele is present in two genotypes, AA and AO. The frequency of the AA genotype is represented by p2, and the AO genotype by Ipr, Therefore, the combined frequency of type A blood and type O blood is given by p2 + 2pr + r2 = If we factor the left side of the equation and take the sum of the terms on the right, we get (p + r)2 = 0.79 p + r = √0./79, p = r ,p = = 0.38 Having estimated p and r, the frequencies of allele A and allele 0, we can now estimate the frequency for the B allele: p + q + r = 1 q = p - r = = 0.11
2 = p + r = √0./79, p = r ,p = = Having estimated p and r, the frequencies of allele A and allele 0, we can now estimate the frequency for the B allele: p + q + r = 1 q = 1 - p - r. = =")
40
22.5 Using the Hardy-Weinberg law: Calculating Heterozygote Frequency
In one application, the Hardy-Weinberg law allows us to estimate the frequency of heterozygotes in a population. The frequency of a recessive trait can usually be determined by counting such individuals in a sample of the population. With this information and the Hardy-Weinberg law, we can then calculate the allele and genotype frequencies. Cystic fibrosis, an autosomal recessive trait, has an incidence of about 1/2500 = in people of northern European ancestry. Individuals with cystic fibrosis are easily distinguished from the population at large by such symptoms as salty sweat,excess amounts of thick mucus in the lungs, and susceptibility to bacterial infections. Because this is a recessive trait, individuals with cystic fibrosis must be homozygous.

41
Their frequency in a population is represented by q2, provided that mating has been random in the previous generation. The frequency of the recessive allele therefore is q = √q2 = √ = 0.02

42
Since p + q = 1, then the frequency of p is
In the Hardy-Weinberg equation, the frequency of heterozygotes is 2pq, 2pq = 2(0.98)(0.02) = 0.04, or 4%, or 1/25 Thus, heterozygotes for cystic fibrosis are rather common in the population (4%), even though the incidence of homozygous recessives is only 1/2500, or 0.04 percent.
(0.02) = 0.04, or 4%, or 1/25. Thus, heterozygotes for cystic fibrosis are rather common in the population (4%), even though the incidence of homozygous recessives is only 1/2500, or 0.04 percent.")
43
In general, the frequencies of all three genotypes can be estimated once the frequency of either allele is known and Hardy-Weinberg assumptions are invoked. The relationship between genotype and allele frequency is shown in Figure 22-6. it is important to note that heterozygotes increase rapidly in a population as the values of p and q move from 0 or 1. This observation confirms our conclusion that when a recessive trait such as cystic fibrosis is rare, the majority of those carrying the allele are heterozygotes. In populations in which the frequencies of p and q are between 0.33 and 0.67, heterozygotes occur at higher frequency than either homozygote.

44
22.6 Factors That Alter Allele Frequencies in Populations
We have noted that the Hardy-Weinberg law establishes an ideal population that allows us to estimate allele and genotype frequencies in populations in which the assumptions of random mating, absence of selection and mutation, and equal viability and fertility hold. Obviously, it is difficult to find natural populations in which all these assumptions hold. In nature, populations are dynamic, and changes in size and gene pool are common. The Hardy-Weinberg law allows us to investigate populations that vary from the ideal. In this and following sections, we discuss factors that prevent populations from reaching Hardy-Weinberg equilibrium, or that drive populations toward a different equilibrium, and the relative contribution of these factors to evolutionary change.

45
Natural Selection The first assumption of the Hardy- Weinberg law is that individuals of all genotypes have equal rates of survival and equal reproductive success. If this assumption does not hold, allele frequencies may change from one generation to the next. To see why, let's imagine a population of 100 individuals in which the frequency of allele A , a Assuming the previous generation mated randomly, the genotype frequencies (0.5) for AA, AA individuals 2(0.5X0.5) = 0.5 for Aa, 50 Aa individuals (0.5)2 = 0.25 for aa, 25 aa individuals.
 for AA, 25 AA individuals. 2(0.5X0.5) = 0.5 for Aa, 50 Aa individuals. (0.5)2 = 0.25 for aa, 25 aa individuals.")
46
Now suppose that individuals with different genotypes have different rates of survival:
All 25 AA individuals survive to reproduce, 90 percent or 45 of the Aa individuals 80 percent or 20 of the aa individuals When the survivors reproduce, each contributes two gametes to the new gene pool, giving us 2(25) + 2(45) + 2(20) = 180 gametes. What are the frequencies of the two alleles in the surviving population? 50 A gametes from AA + 45 A gametes from Aa, so the frequency of A is ( )/180 = 0.53. Frequency of allele a is ( )/180 = 0.47. Allele A has increased, while allele a has declined. A difference among individuals in survival and/or reproduction rate is called natural selection. Natural selection is the principal force that shifts allele frequencies within large populations and is one of the most important factors in evolutionary change.
 + 2(45) + 2(20) = 180 gametes. What are the frequencies of the two alleles in the surviving population 50 A gametes from AA + 45 A gametes from Aa, so the frequency of A is ( )/180 = Frequency of allele a is ( )/180 = Allele A has increased, while allele a has declined. A difference among individuals in survival and/or reproduction rate is called natural selection. Natural selection is the principal force that shifts allele frequencies within large populations and is one of the most important factors in evolutionary change.")
47
Fitness and Selection Selection occurs whenever individuals type enjoy an advantage over other genotypes. However, selection may be weak or strong, depending on the magnitude of the advantage. In the example above, selection was strong. Weak selection might involve just a fraction of a percent difference in the survival rates of different genotypes. Advantages in survival and reproduction ultimately translate into increased genetic contribution to future generations. An individual's genetic contribution to future generations is called fitness. Thus, genotypes associated with high rates of survival and/or high reproductive success are said to have high fitness, whereas genotypes associated with low rates of survival and/or low reproductive success are said to have low fitness.

48
wAA for genotype AA, waa = 1
waa for aa. wafl = 0.8 \Let's consider selection against deleterious alleles. Fitness values wAA = 1, wAa = 1, and wbb = 0 describe a situation in which allele a is a lethal recessive. Homozygote recessive individuals die without leaving offspring, the frequency of allele a will decline. The decline in the frequency of allele a is described by the equation Qg= qo / 1-gqo

49
intensity of selection

50
The manner in which selection affects allele frequencies allows us to make some inferences about the CCK5-A32 allele A32/A32, 1/1 and 1/A32 current frequency of the A32 allele is 0.10. the genotype frequencies for 1/1, 0.18 for 1/A32, and 0.01 for A32/A32. Imagine also that 1 percent of the 1/1 and 1/A32 individuals in this population will contract HIV and die of AIDS. fitness levels as follows: w1/1 = 0.99; w1/432 = 0.99; wA32/A32 = 1.0. Given the assigned fitness, we can predict that the frequency of the CCR5-A32 allele in the next generation will be

51
In fact, it will take about 100 generations (about 2000 years) for the frequency of the A32 allele to reach just 0.11 (Figure 22-9). In other words, the frequency of the A32 allele will probably not change much over the next few generations in most populations that currently harbor it.

52
A population genetic perspective sheds light on the CCR5-A32 story in other ways as well. Two research groups have analyzed genetic variation at marker loci closely linked to the CCR5 gene. Both groups concluded that most, if not all present-day copies of the A32 allele are descended from a single ancestral copy that appeared in northeastern Europe at most a few thousand years ago. In fact, one group estimates that the common ancestor of all A32 alleles existed just 700 years ago. How could a new allele rise from a frequency of virtually zero to as high as 20 percent in roughly 30 generations?

53
It seems that there must have been strong selection in favor of the A32 allele. The agent of selection cannot have been HIV-l. because HIV-1 moved from chimpanzees to humans too recently. Because selection occurred about 700 years ago, J. C. Stephens suggests that the agent of selection was bubonic plague. During the Black Death of , between a quarter and a third of all Europeans died from plague. Bubonic plague is caused by the bacterium Yersinia pestis. This bacterium manufactures a protein that kills some kinds of white blood cells. Stephens hypothesizes that the process through which the bacterial protein kills white cells involves the CCR5 gene product. If true, some mechanism makes individuals homozygous for the A32 allele more likely to survive plague epidemics.

54
If past epidemics of bubonic plague are responsible for the high frequency of the CCR5-A32 allele in European populations, men the virtual absence of the allele in non-European populations is at first somewhat puzzling. Perhaps plague has been more common in Europe than elsewhere. Alternatively, perhaps other populations have different alleles of the CCR5 gene that also confer protection against the plague. Teams of researchers looking for other alleles of the CCR5 gene in various populations have found a total of 20 mutant alleles, including A32. Sixteen of these alleles encode proteins different in structure from that encoded by the CCR51 allele. Some, like A32, are loss-of-function alleles. Some of the alleles appear confined to Asian, others to African. as high as 3-4 percent. Together, these discoveries are consistent with the hypothesis that alteration or loss of the CCR5 protein protects against an as-yet-unidentified infectious disease or diseases.

55
Selection in Natural Populations
effect of natural selection, for example, studied the effect of the insecticide chlorpyrifos on allele frequencies in populations of house mosquitoes (Figure 22-10). Chlorpyrifos kills mosquitoes by interfering with the function of the enzyme acetylcholinesterase (ACE), which under normal circumstances breaks down the neurotransmitter acetylcholine. An allele of the gene encoding ACE called Ace" encodes a slightly altered version of ACE that is immune to interference by chlorpyrifos.
. Chlorpyrifos kills mosquitoes by interfering with the function of the enzyme acetylcholinesterase (ACE), which under normal circumstances breaks down the neurotransmitter acetylcholine. An allele of the gene encoding ACE called Ace encodes a slightly altered version of ACE that is immune to interference by chlorpyrifos.")
56
FIGURE Pesticides used to control insects act as selective agents, changing allele frequencies of resistance genes in the populations

57
Chevillon measured the frequency of the AceR allele in nine populations. In the first four locations, chlorpyrifos had been used to control mosquitoes for 22 years; in the last five locations, chlorpyrifos had never been used. Chevillon predicted that the frequency of AceR would be higher in the exposed populations. The researchers also predicted that the frequencies of alleles for enzymes unrelated to the physiological effects of chlorpyrifos would show no such pattern. Among the control enzymes studied was aspartate amino transferase 1.

58
The results appear in Figure 22-11
The results appear in Figure As the researchers predicted, the frequency of the AceR allele was significantly higher in the exposed populations. Also as predicted, the frequencies of the most common alleles of the control enzyme genes showed no such trends. The explanation is that during the 22 years of exposure to the pesticide, mosquitoes had higher rates of survival if they carried the AceR allele. In other words, the AceR allele had been favored by natural selection

59
Natural Selection and Quantitative Traits Selection for such traits can be classified as
(1) directional, (2) stabilizing, or (3) disruptive.
 directional, (2) stabilizing, or (3) disruptive.")
60
(2) stabilizing In contrast, tends to favor intermediate types, with both extreme phenotypes being selected against. One of the clearest demonstrations of stabilizing selection is provided by the data of Mary Karn and Sheldon Penrose on human birth weight and survival for 13,730children born over an 11-year period. Figure shows the distribution of birth weight and the percentage of mortality at 4 weeks of age. Infant mortality increases on either side of the optimal birth weight of 7.5 pounds, and quite dramatically so at the low end. At the genetic level, stabilizing selection acts to keep a population well adapted to its environment. In this situation, individuals closer to the average for a given trait will have higher fitness.

61
(3) disruptive
 disruptive")
62
Disruptive selection is selection against intermediates and for both phenotypic extremes. It can be viewed as the opposite of stabilizing selection because die intermediate types are selected against. In one set of experiments, John Thoday applied disruptive selection to a population of Drosophila based on bristle number. In every generation he allowed only the flies widi high or low bristle numbers to breed. After several generations, most of the flies could be easily placed in a low or high bristle category (Figure 22-14). In natural populations, such a situation might exist for a population in a heterogeneous environment. The types and effects of selection are summarized in Figure
. In natural populations, such a situation might exist for a population in a heterogeneous environment. The types and effects of selection are summarized in Figure")
63
Mutation Reshuffled each generation to produce new genotypes in the offspring. But do not produce new alleles. Mutation alone acts to create new alleles. It is important to keep in mind that mutational events occur at random—that is, without regard for any possible benefit or disadvantage to the organism. In this section we consider whether mutation is, by itself, a significant factor in causing allele frequencies to change.

64
To determine whether mutation is a significant force in changing allele frequencies, we measure the rate at which mutations are produced. As most mutations are recessive, it is difficult to observe mutation rates directly in diploid organisms. Indirect methods using probability and statistics or large-scale screening programs are employed. For certain dominant mutations, however, a direct method of measurement can be used. To ensure accuracy, several conditions must be met: 1. The allele must produce a distinctive phenotype that can be distinguished from similar phenotypes produced by recessive alleles. 2. The trait must be fully expressed or completely penetrant so that mutant individuals can be identified. 3. An identical phenotype must never be produced by non genetic agents such as drugs or chemicals.

65
First.Mutation rates that must be known ,can be stated as the number of new mutant alleles per given number of gametes. 1/100, X In humans, a dominant form of dwarfism known as achondroplasia u = 1.4 X 10-5 ± 0.5 X 10-5 Knowing the rate of mutation, we can estimate the extent to which mutation can cause allele frequencies to change from one generation to the next. We represent the normal allele as d, and the allele for achondroplasia as D. Imagine a population of 500,000 individuals in which everyone has genotype dd. The initial frequency of d 1.0, and the initial frequency of D is 0.

66
Pg+1 =pg-upg, and qg+l = qg + upg.
If each individual contributes 2 gametes to the gene pool, the gene pool will contain 1,000,000 gametes, all carrying allele d. While the gametes are in the gene pool, 1.4 of every 100,000 d alleles mutates into a D allele. The frequency of allele d is now (1,000, )/1,000,000 = , and the frequency of D is 14/1,000,000 = More generally, if we have two alleles, A with frequency p and a with frequency q, and if u represents the rate of mutations converting A into a, then the frequencies of the alleles in the next generation are given by Pg+1 =pg-upg, and qg+l = qg + upg. where pg+1 and qg+l represent the allele frequencies in the next generation, and pg and qg represent the allele frequencies in the present generation.
/1,000,000 = , and the frequency of D is 14/1,000,000 = More generally, if we have two alleles, A with frequency p and a with frequency q, and if u represents the rate of mutations converting A into a, then the frequencies of the alleles in the next generation are given by. Pg+1 =pg-upg, and qg+l = qg + upg. where pg+1 and qg+l represent the allele frequencies in the next generation, and pg and qg represent the allele frequencies in the present generation.")
68
Figure shows the replacement rate (change over time) in allele A for a population in which the initial frequency of A is 1.0, and the rate of mutations (/j.) converting A into a is 1.0 X 10~5. At this mutation rate, it will take about 70,000 generations to reduce the frequency of A to 0.5. Even if the rate of mutation increases through exposure to higher levels of radioactivity or chemical mutagens, the impact of mutation on allele frequencies will be extremely weak. The ultimate source of the genetic variability, mutation provides the raw material for evolution, but by itself p\ay$ a relatively insignificant role in changing allele frequencies. Instead, the fate of alleles created by mutation is more likely to be determined by natural selection (discussed previously) and genetic drift (discussed later).
 and genetic drift (discussed later)..")
69
The frequency for the mutant alleles causing cystic fibrosis an autosomal recessive disorder, which we discussed previously in relation to calculating heterozygote frequencies, is about 2 percent in European populations. Until recently, most individuals with two mutant alleles died before reproducing, meaning that selection against homozygous recessives was rather strong. This creates a puzzle: In the face of selection against them, what has maintained the mutant alleles at an overall frequency of 2 percent? Although several hypotheses have been put forward, many evolutionary geneticists prefer the heterozygote superiority hypothesis. According to the heterozygote superiority hypothesis, selection against homozygous mutant individuals is counter balanced by selection in favor of heterozygotes.

70
Recent work suggests that cystic fibrosis heterozygotes may have enhanced resistance to typhoid fever. Typhoid fever is caused by the bacterium Salmonella typhi, which infiltrates cells of the intestinal lining. In laboratory studies, mouse intestinal cells that were heterozygous for CFTR-AF508, the analog of the most common cystic fibrosis mutation in humans, acquired 86 percent fewer bacteria than did cells homozygous for the wild-type allele. Whether humans heterozygous for CFTR-A508 also enjoy resistance to typhoid fever remains to be established. If they do, then cystic fibrosis will join sickle-cell anemia as an example of heterozygote superiority.

71
Migration Occasionally, a species divides into populations that to some extent are separated geographically. Various evolutionary forces, including selection can establish different allele frequencies in such populations. Migration occurs when individuals move between the populations. Imagine a species in which a single locus has two alleles, A and a. There are two populations of this species, one on a mainland, and one on an island. The frequency of A on the mainland is represented by pm and the frequency of A on the island is/>j. Under the influence of migration from the mainland to the island, the frequency of A in the next generation on the island (/?,.) is given by
 is given by.")
72
where m represents migrants from (he mainland to the island
where m represents migrants from (he mainland to the island. Under these conditions, the frequency of A in the next generation on the island (/>,.) will be affected by migration. For example, assume that pt = 0.4 and pm = 0.6, and that 1 0 percent of the parents of the next generation are migrants from the mainland, so that m = In the next generation, the frequency of allele A on the island will be /),• = [( ) X 0.4] + (0.1 X 0.6) = In this case, migration from the mainland has changed the frequency of A on the island from 0.40 to 0.42 in a single generation.
 will be affected by migration. For example, assume that pt = 0.4 and pm = 0.6, and that 1 0 percent of the parents of the next generation are migrants from the mainland, so that m = In the next generation, the frequency of allele A on the island will be. /),• = [( ) X 0.4] + (0.1 X 0.6) = In this case, migration from the mainland has changed the frequency of A on the island from 0.40 to 0.42 in a single generation.")
73
If either m is large or if pm is very different from p{, then a rather large change in the frequency of A can occur in a single generation. If migration is the only force acting to change the allele frequency on the island, then an equilibrium will be attained only when p, = pm. These calculations reveal that the change in allele frequency attributable to migration is proportional to the differences in allele frequency between the donor and recipient populations and to the rate of migration. As m can have a wide range of values, the effect of

74
i migration can substantially alter allele frequencies in popu-I lations, as shown for the 6 allele of the ABO blood group in Figure Although migration can be difficult to quan-| tify, it can often be estimated. I Migration can also be regarded as the flow of genes be-j tween populations that were once, but are no longer, geo-! graphically isolated. Esteban Parra and colleagues measured j allele frequencies for several different DNA sequence poly-I morphisms in African-American and European-Am eric an ] populations, and in African and European populations rep-I resentative of the ancestral populations from which the two : American populations are descended.

75
One locus they stud-; ied, a restriction site polymorphism called FY-NULL, has two alleles, FY-NULL*! and FY-NULL*2. Figure shows the frequency of FY-NULL*! in each population. The frequency of this allele is 0 in the three African populations, and 1.0 in the three European populations, but lies between these extremes in all African-American and European-American populations. The simplest explanation for these data is that genes have mixed between American populations with predominantly African ancestry and American populations with predominantly European ancestry. Based on FY-NULL and several other loci, researchers estimate that African-American populations derive between 11.6 and 22.5 percent of their ancestry from Europeans, and that European-American populations derive between 0.5 and 1.2 percent of their ancestry from Africans.

77
Genetic Drift In laboratory crosses, one condition essential to realizing theoretical genetic ratios (e.g., 3:1, 1:2:1,9:3:3:1) is a fairly large sample size. A large sample size is also important to the study of population genetics as allele and genotype frequencies are examined or predicted. For example, if a population consists of 1000 randomly mating heterozygotes (Aa), the next generation will consist of approximately 25 percent AA, 50 percent Aa, and 25 percent aa genotypes. Provided that the initial and subsequent populations are large, only minor deviations from this mathematical ratio will occur, and the frequencies of A and a will remain about equal (at 0.50).
 is a fairly large sample size. A large sample size is also important to the study of population genetics as allele and genotype frequencies are examined or predicted. For example, if a population consists of 1000 randomly mating heterozygotes (Aa), the next generation will consist of approximately 25 percent AA, 50 percent Aa, and 25 percent aa genotypes. Provided that the initial and subsequent populations are large, only minor deviations from this mathematical ratio will occur, and the frequencies of A and a will remain about equal (at 0.50).")
78
However, if a population consists of only one set of heterozygous parents and they produce only two offspring, the allele frequency can change drastically. We can also predict genotypes and alleie frequencies in the offspring in this case. As Table 22.5 shows, in 10 of 16 times such a cross is made, the allele frequencies would be altered. In 2 of 16 times, either the A or a allele would be eliminated in a single generation. Founding a population with only one set of heterozygous parents that produce two offspring is an extreme example, but it illustrates the point that large interbreeding populations are essential to maintain Hardy-Weinberg equilibrium. In small populations, significant random fluctuations in allele frequencies are possible by chance deviation. The degree of fluctuation increases as the population size decreases, a situation known as genetic drift. In the extreme case, genetic drift can lead to the chance fixation of one allele to the exclusion of another allele.

79
To study genetic drift in laboratory populations of Drosophila melanogaster, Warwick Kerr and Sewall Wright set up over 100 lines, with four males and four females as the parents for each line. Within each line, the frequency of the sex-linked bristle mutant forked (/) and its wild-type allele (/+) was 0.5. In each generation, four males and four females were chosen at random to parent the next generation. After 16 generations, the complete loss of one allele and the fixation of the other had occurred in 70 lines—29 in which only the forked allele was present and 41 in which the wild-type allele had become fixed. The remaining lines were still segregating the two alleles or had gone extinct. If fixation had occurred randomly, then an equal number of lines should have become fixed for each allele. In fact, the experimental results do not differ statistically from ihe expected ratio of 35:35, demonstrating that alleles can spread through a population and eliminate other alleles by chance alone.

80
How are small populations created in nature
How are small populations created in nature? In one scenario, a large population may be split by some event (like war), creating a small isolated subpopulation. A disaster such as an epidemic might occur, leaving a small number of survivors to breed. Or, a small group might emigrate from the larger population and become founders in a new environment, such as a volcanically created island. Allele frequencies in certain isolated human populations demonstrate the role of drift as an evolutionary force in natural populations. The Pingelap atoll in the western Pacific Ocean (lat. 6° N, long. 160° E) has in the past been devastated by typhoons and famine, and around 1780 there were only about nine surviving males.
, creating a small isolated subpopulation. A disaster such as an epidemic might occur, leaving a small number of survivors to breed. Or, a small group might emigrate from the larger population and become founders in a new environment, such as a volcanically created island. Allele frequencies in certain isolated human populations demonstrate the role of drift as an evolutionary force in natural populations. The Pingelap atoll in the western Pacific Ocean (lat. 6° N, long. 160° E) has in the past been devastated by typhoons and famine, and around 1780 there were only about nine surviving males.")
81
Today there are fewer than 2000 inhabitants, all of whose ancestry can be traced to the typhoon survivors. About 4-10 percent of the current population is blind from infancy. These people are affected by an autosomal recessive disorder, achromatopsia, which causes ocular disturbances, a form of color blindness, and cataract formation. The disorder is extremely rare in the human population as a whole. However, the mutant allele is present at a relatively high frequency in the Pingelap population. Genealogical reconstruction shows that one of the original survivors (about 30 individuals) was heterozygous for the condition- If we assume that he was the only carrier in the founding population, the initial gene frequency was 1/60, or On average, about 7 percent of the current population is affected (a homozygous recessive genotype), so the frequency of the allele in the present population has increased to The story of the inhabitants of Pingelap is told by Oliver Sacks, in his book, The Island of rhe Colorblind,
 was heterozygous for the condition- If we assume that he was the only carrier in the founding population, the initial gene frequency was 1/60, or On average, about 7 percent of the current population is affected (a homozygous recessive genotype), so the frequency of the allele in the present population has increased to The story of the inhabitants of Pingelap is told by Oliver Sacks, in his book, The Island of rhe Colorblind,.")
83
Nonrandom Mating We have explored how violations of the first four assumptions of the Hardy-Weinberg law, in the form of selection, mutation, migration, and genetic drift, can cause allele frequencies to change. The fifth assumption is that the members of a population mate at random. Nonrandom mating itself does not directly alter the frequencies of alleles. It can, however, alter the frequencies of genotypes in a population and thereby indirectly affect the course of The most important form of nonrandom mating, and the form we focus on here, is inbreeding, mating between relatives. For a given allele, inbreeding increases the proportion of homozygotes in the population. Over time, with complete inbreeding, only homozygotes will remain in the population. To demonstrate this concept, we consider the most extreme form of inbreeding, self-fertilization.

84
Inbreeding Figure shows the results of four generations of self-fertilization starting with a single individual heterozygous for one pair of alleles. By the fourth generation, only about 6 percent of the individuals are still heterozygous, and 94 percent of the population is homozygous. Note, however, that alleles A and a still remain at 50 percent. In humans, inbreeding (called consanguineous marriage) is related to population size, mobility, and social customs governing marriages among relatives. To describe the amount of inbreeding in a population, Sewall Wright devised the coefficient of inbreeding. Expressed as F, the coefficient of inbreeding is defined as the probability that the two alleles of a given gene in an individual are identical because they are cestor. If F = 1, all individuals are homozygous, and both alleles in every individual are derived from the same ancestral copy. If F = 0, no individual has two alleles derived from a common ancestral copy.
 is related to population size, mobility, and social customs governing marriages among relatives. To describe the amount of inbreeding in a population, Sewall Wright devised the coefficient of inbreeding. Expressed as F, the coefficient of inbreeding is defined as the probability that the two alleles of a given gene in an individual are identical because they are. cestor. If F = 1, all individuals are homozygous, and both alleles in every individual are derived from the same ancestral copy. If F = 0, no individual has two alleles derived from a common ancestral copy.")
85
Figure 22-20 shows a pedigree of a first-cousin marriage
Figure shows a pedigree of a first-cousin marriage. The fourth-generation female (shaded pink) is the daughter of first cousins (purple). Suppose her great-grandmother (green) was a carrier of a recessive lethal allele, a. What is the probability that the fourth-generation female will inherit two copies of her great-grandmother's lethal allele? For this to happen, (1) the great-grandmother had to pass a copy of the allele to her son, (2) her son had to pass it to his daughter, and (3) his daughter has to pass it to her daughter (the pink female). Also, (4) the great-gran dm other had to pass a copy of the allele to her daughter, (5) her daughter had io pass it to her son, and (6) her son has to pass it to his daughter (the pink female).
 is the daughter of first cousins (purple). Suppose her great-grandmother (green) was a carrier of a recessive lethal allele, a. What is the probability that the fourth-generation female will inherit two copies of her great-grandmother s lethal allele For this to happen, (1) the great-grandmother had to pass a copy of the allele to her son, (2) her son had to pass it to his daughter, and (3) his daughter has to pass it to her daughter (the pink female). Also, (4) the great-gran dm other had to pass a copy of the allele to her daughter, (5) her daughter had io pass it to her son, and (6) her son has to pass it to his daughter (the pink female).")
86
Each of the six necessary events has an individual probability of 1/2, and they all have to happen, so the overall probability that the pink female will inherit two copies of her great-grandmother's lethal allele is (1/2)6 = 1/64. This takes us most of the way toward calculating F for a child of a first-cousin marriage. We need only note that the fourth-generation female could also inherit two copies of any of the other three alleles present in her great-grandparents. Because any of four possibilities would give the pink female two alleles identical by descent from an ancestral copy, F = 4 X (1/64) = 1/16.
 = 1/16.")
89
Genetic Effects of Inbreeding
Inbreeding results in the production of individuals homozy-gous for recessive alleles that were previously concealed in heterozygotes. Because many recessive alleles are deleterious when homozygous, one consequence of inbreeding is an increased chance that an individual will be homozygous for a recessive deleterious allele. Inbred populations often have a lowered mean fitness. Inbreeding depression is a measure of the loss of fitness caused by inbreeding. In domesticated plants and animals, inbreeding and selection have been used for thousands of years, and these organisms already have a high degree of homozygosity at many loci. Further inbreeding will usually produce only a small loss of fitness.

90
However, inbreeding among individuals from large, randomly maiing populations can produce high levels of inbreeding depression. This effect can be seen by examining the mortality rates in offspring of inbred animals in zoo pop- many zoos use DNA fingerprinting to estimate the related-ness of animals used in breeding programs and choose the least related animals as parents. As natural populations of endangered species decrease, the concern about inbreeding is a factor in designing programs to restore these species. One example is a project in Scandanavian zoos to preserve the Fennoscandic wolf. Conservationists established a captive-bred wolf population wilh four founding individuals.

91
Later, two Russian wolves were added to the population to reduce the level of inbreeding, a strategy some conservationists later came to regret because it introduced foreign alleles into the Fennoscandic gene pool. Because the captive wolf line was founded by such a small number of individuals, it is severely inbred, and individuais show inbreeding depression in the form of smaller body weight, reduced reproductive success, and reduced longevity. Furthermore, a number of wolves in the line are blind. Researchers concluded that blindness in the captive wolf locus.

92
The bad news is that all the remaining pure-bred Fennoscandic wolves (i.e., individuals with no genes from the two Russian wolves) have at least a 6 percent chance of being carriers for blindness. Some pure-bred individuals have a 67 dividuals with a greater than 30 percent chance of being carriers can be removed from the breeding population without reducing the remaining genetic variation in the population by more than 10 percent. Removing the likely carriers would reduce the frequency of the blindness allele from 14 percent to 7 percent, and would improve the long-term prospects of maintaining a viable Fennoscandic wolf population. abortions, neonatal deaths, congenital deformities and recessive genetic disorders. Although less common than in the past, inbreeding occurs in many regions of the world where Kittles and James Neel analyzed data from numerous studies on different cultures. They found that the rate of child mortality (i.e., death in the first several years of life) varies dramatically from culture to culture.
 varies dramatically from culture to culture.")
93
No matter what the baseline mortality rate for children of unrelated parents, however, children of first cousins virtually always have a higher death rate—typically by about 4.5 percentage points. Over many generations, inbreeding should eventually reduce the frequency of deleterious recessive alleles (if homozygotes die before reproducing). However, some studies indicate that parents who are first cousins tend to have more children to compensate for those lost to genetic disorders. On average, two-thirds of the surviving offspring are heterozygous carriers of the deleterious allele. It is important to note that inbreeding is not always harmful. Indeed, inbreeding has long been recognized as a useful tool for breeders of domesticated plants and animals. When an inbreeding program is initiated, homozygosity increases, and some breeding stocks become fixed for favorable alleles and others for unfavorable alleles. By selecting the more viable and vigorous plants or animals, the proportion of individuals carrying desirable traits can be increased.

94
If members of two favorable inbred lines are mated, hybrid
ther of the parental lines. This phenol grams established for maize, crop yields increased tremendously. Unfortunately, the hybrid vigor extends only through the first generation. Many hybrid lines are sterile, and those that are fertile show subsequent declines in yield. crossing the original inbred parental tines. Hybrid vigor has been explained in two ways. The first theory, the dominance hypothesis, incorporates the obvious reversal of inbreeding depression, which inevitably must occur ailed hybrid maize with the following genotypes: Strain A Strain B F, aaBBCCddee X AAbbccDDEE » AaBhCcDdEe

95
The F, hybrids are heterozygotes at all loci shown
The F, hybrids are heterozygotes at all loci shown. The deleterious recessive alleles present in the homozygous form in the parents is masked by the more favorable dominant alleles in the hybrids. Such masking is thought to cause hybrid vigor. The second theory, overdominance. holds that in many cases the heterozygote is superior to either homozygote. This may relate to the fact that in the heterozygote two forms of a gene product may be present, providing a form of biochemical diversity. Thus, the cumulative effect of heterozy-gosity at many loci accounts for the hybrid vigor. Most likely,

96
plained by both hypotheses.
We have seen that nonrandom mating can drive the genotype frequencies in a population away from their expected values under the Hardy-We in berg law. This can indirectly affect the course of evolution. As in stocks purposely inbred by animal and plant breeders, inbreeding in a natural population may increase the frequency of homozygotes for a deleterious recessive allele. As in domestic stocks, this increases the efficiency with which selection removes the deleterious allele from the population.

97
That’ s all for this chapter !

Similar presentations











>")















