Download presentation
Presentation is loading. Please wait.

1
Adventures in Computational Enzymology John Mitchell

2
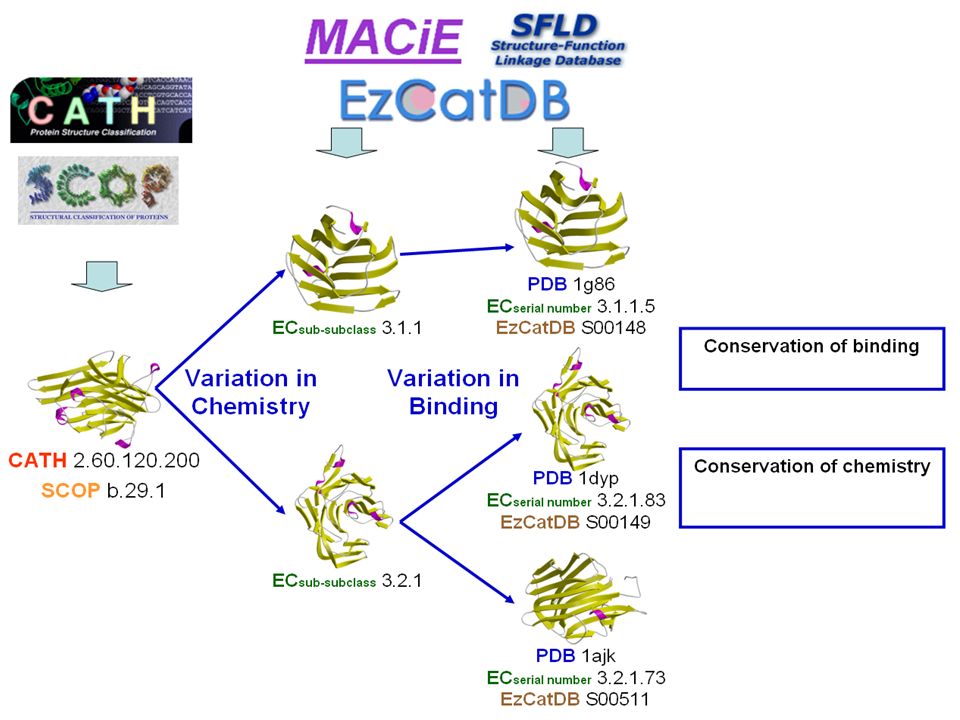
MACiE Mechanism, Annotation and Classification in Enzymes. http://www.ebi.ac.uk/thornton-srv/databases/MACiE/ MACiE Database G.L. Holliday et al., Nucl. Acids Res., 35, D515-D520 (2007)
.")
3
Enzyme Nomenclature and Classification EC Classification Class Subclass Sub-subclass Serial number

4
EC Classification Enzyme Commission (EC) Nomenclature, 1992, Academic Press, San Diego, 6 th Edition Chemical reaction
 Nomenclature, 1992, Academic Press, San Diego, 6 th Edition Chemical reaction")
5
The EC Classification Reaction direction arbitrary. Doesn’t deal with structural and sequence information. Thus, cofactors and active site residues ignored. However, it was never intended to describe mechanism. Only deals with overall reaction.

6
A New Representation of Enzyme Reactions? Should be complementary to, but distinct from, the EC system. Should take into account: Reaction Mechanism; Structure; Sequence. Need a database of enzyme mechanisms.

7
MACiE Mechanism, Annotation and Classification in Enzymes. http://www.ebi.ac.uk/thornton-srv/databases/MACiE/ MACiE Database

8
Coverage of MACiE Representative – based on a non-homologous dataset, and chosen to represent each available EC sub-subclass.

9
Coverage of MACiE Representative – based on a non-homologous dataset, and chosen to represent each available EC sub-subclass. Structures exist for: 6 EC 1.-.-.- 56 EC 1.2.-.- 184 EC 1.2.3.- 1312 EC 1.2.3.4 MACiE covers: 6 EC 1.-.-.- 53 EC 1.2.-.- 156 EC 1.2.3.- 199 EC 1.2.3.4

10
Repertoire of Enzyme Catalysis G.L. Holliday et al., J. Molec. Biol., 372, 1261-1277 (2007)
")
11
Repertoire of Enzyme Catalysis Enzyme chemistry is largely nucleophilic

12
Protontransfer Ad N 2 E1 SN2SN2SN2SN2E2 RadicalreactionTautom.Others Repertoire of Enzyme Catalysis

13
Residue Catalytic Propensities

14
Evolution of Enzyme Function D.E. Almonacid et al., to be published

15
Work with domains - evolutionary & structural units of proteins. Map enzyme catalytic mechanisms to domains to quantify convergent and divergent functional evolution of enzymes. Domains

16
Functional Classification: EC Enzyme Commission (EC) Nomenclature, 1992, Academic Press, San Diego, 6 th Edition Chemical reaction
 Nomenclature, 1992, Academic Press, San Diego, 6 th Edition Chemical reaction")
17
Enzyme Catalysis Databases G.L. Holliday et al., Nucleic Acids Res., 35, D515 (2007) S.C. Pegg et al., Biochemistry, 45, 2545 (2006) N. Nagano, Nucleic Acids Res., 33, D407 (2005)
 N. Nagano, Nucleic Acids Res., 33, D407 (2005).")
18
Coverage of MACiE Representative – based on a non-homologous dataset, and chosen to represent each available EC sub-subclass.

19
Coverage of SFLD Based on a few evolutionarily related families

20
Coverage of EzCatDB But without mechanisms.

21
Structural Classification: CATH Orengo, C. A., et al. Structure, 1997, 5, 1093

22
Dataset CATH Enzymes in (single-domain) PDB Database entries395 >>799 EC sub-subclasses114 184 EC serial numbers326 1312 To avoid the ambiguity of multi-domain structures we use only single-domain proteins.
 PDB Database entries395 >>799 EC sub-subclasses EC serial numbers To avoid the ambiguity of multi-domain structures we use only single-domain proteins.")
23
Numbers of CATH code occurrences per EC number C A T H c.-.-.- c.s.-.- c.s.ss.- c.s.ss.sn 3.17 11.00 28.00 38.33 1.73 3.27 4.89 5.80 1.38 1.93 2.24 2.46 1.11 1.60 1.19 1.22 Results: Convergent Evolution 2.46 CATH/EC reaction Convergent Evolution

24
Numbers of CATH code occurrences per EC number C A T H c.-.-.- c.s.-.- c.s.ss.- c.s.ss.sn 3.17 11.00 28.00 38.33 1.73 3.27 4.89 5.80 1.38 1.93 2.24 2.46 1.11 1.60 1.19 1.22 Results: Convergent Evolution 2.46 CATH/EC reaction: Convergent Evolution An average reaction has evolved independently in 2.46 superfamilies

25
EC reactions/CATH C 4.75 19.50 39.25 90.00 c.-.-.-c.s.-.- c.s.ss.- c.s.ss.sn A 3.14 7.00 10.48 17.90 T 1.36 1.79 2.08 3.05 H 1.20 1.36 1.46 2.05 database entries/CATH 2.18 Results: Divergent Evolution 1.46 EC reactions/CATH Divergent Evolution

26
EC reactions/CATH C 4.75 19.50 39.25 90.00 c.-.-.-c.s.-.- c.s.ss.- c.s.ss.sn A 3.14 7.00 10.48 17.90 T 1.36 1.79 2.08 3.05 H 1.20 1.36 1.46 2.05 database entries/CATH 2.18 Results: Divergent Evolution 1.46 EC reactions/CATH: Divergent Evolution An average superfamily has evolved 1.46 different reactions

27
Density Functional Theory Calculations on Dehydroquinase Mattias Blomberg et al., to be published

28
DFT – System Size System sizes of ~100- 150 atoms can be treated using DFT That raises the question of how to treat the rest of the protein.

29
Dielectric Continuum or QM/MM? One approach is to cut out the active site residues and treat the rest of the protein as a dielectric continuum. Another approach is to treat the active site as QM and the rest of the protein using MM. QMQM ε=4 QMQM MM

30
Dielectric Continuum or QM/MM? One approach is to cut out the active site residues and treat the rest of the protein as a dielectric continuum. Another approach is to treat the active site as QM and the rest of the protein using MM. QMQM ε=4 QMQM MM

31
Dehydroquinase - Part of the Shikimate Pathway

32
Shikimate & Chorismate Pathways

33
Dehydroquinase (Shikimate Pathway)
")
34
Shikimate & Chorismate Pathways Biosynthetic pathway for phenylalanine, tyrosine and tryptophan. Present in plants, microorganisms and fungi but not in mammals. The target for Glyphosate, an important herbicide. Understanding the mechanisms and developing inhibitors is of great importance for the development of new herbicides, fungicides and antibiotics.

35
Two Types of Dehydroquinases Type I: E. coli and S. typhi, (EC 4.2.1.10) MACiE M0054 Mechanism: cis-dehydration, imine intermediate. Type II: S. coelicor, M. tuberculosis and H. pylori (EC 4.2.1.10). MACiE M0055 Mechanism: trans-dehydration, enol(ate) intermediate.
 MACiE M0054 Mechanism: cis-dehydration, imine intermediate. Type II: S. coelicor, M. tuberculosis and H. pylori (EC ). MACiE M0055 Mechanism: trans-dehydration, enol(ate) intermediate..")
36
Proposed Mechanism of DHQase

37
Models of DHQase Active Site

38
Energetics of DHQase Model A

39
Does Asn16 Protonate the DHQ Enolate?

40
Other Things we do Chemoinformatics for pharmaceutical design … …using Machine Learning for prediction of solubility, bioavailability and bioactivity.

41
Machine Learning Methods Recognise patterns in data Similar inputs Similar outputs Make full use of all available information One application is solubility

42
Machine Learning Methods Can be used for Classification or for Regression Can be used with chemoinformatics, physicochemical or experimental (e.g., assay) data as descriptors
 data as descriptors")
43
Solubility is an important issue in drug discovery and a major source of attrition This is expensive for the industry A good model for predicting the solubility of druglike molecules would be very valuable.

44
Drug Disc.Today, 10 (4), 289 (2005)
, 289 (2005)")
45
Machine Learning Method Random Forest

46
Machine Learning Method k-Nearest Neighbours

47
Machine Learning Method Winnow (“Molecular Spam Filter”)
")
48
Future Directions

49
Current coverage of MACiE Representative – based on a non-homologous dataset

50
Future coverage of MACiE Adding homologues – to facilitate study of divergent evolution

51
Divergent Evolution using MACiE This will use our reaction similarity work to measure changes in chemistry

53
Using Machine Learning Methods to calculate and predict protein-ligand binding energies Building on our previous work … P.M. Marsden et al., Org. Biomol. Chem., 2, 3267 (2004)
.")
54
Computational Toxicology Predicting bioavailability problems, off-target activities and side effects of drug candidates

55
QM, QM/MM and MD Simulation Work Using computational chemistry to study enzyme mechanisms Fosfomycin Resistance Protein A

56
ACKNOWLEDGEMENTS Dr Gemma Holliday Dr Daniel Almonacid Dr Noel O’Boyle Dr Mattias Blomberg Prof. Janet Thornton (EBI) Dr Peter Murray-Rust Dr Jochen Blumberger
 Dr Peter Murray-Rust Dr Jochen Blumberger.")
57
ACKNOWLEDGEMENTS Cambridge Overseas Trust

58
All slides after here are for information only

59
Similarity of Enzyme Mechanisms N.M. O'Boyle, et al., J. Molec. Biol., 368, 1484-1499 (2007)
")
60
Measuring Similarity of Enzyme Mechanisms

61
Coverage of MACiE Representative – based on a non-homologous dataset, and chosen to represent each available EC sub-subclass.

62
Unimolecular Heterolytic Bimolecular Intramolecular Elimination Unimolecular HomolyticBimolecular Intramolecular ElectrophilicBimolecular Intramolecular Addition NucleophilicBimolecular Intramolecular HomolyticBimolecular Intramolecular Unimolecular ElectrophilicBimolecular Intramolecular Unimolecular SubstitutionNucleophilicBimolecular Intramolecular Unimolecular HomolyticBimolecular Intramolecular Ingold, C. K. Cornell University Press, 1969. Repertoire of enzyme catalysis

63
“Other reactions” and Named organic reactions currently supported in MACiE ______________________________________________ Aldol CondensationHydride Transfer Amadori RearrangementIsomerisation A-S N 1Michael Addition A-S N 2Nucleophilic Attack A-S N iPericyclic Reaction Claisen RearrangementProton Transfer CondensationRadical Formation E1cbRadical Propagation Group TransferRadical Termination HeterolysisRedox HomolysisTautomerisation ______________________________________________ Repertoire of enzyme catalysis

64
Functionality for amino acids currently supported in the MACiE ________________________________________________ Activating residueProton acceptor Charge destabiliserProton donor Charge stabiliserProton relay Covalently attachedRadical acceptor ElectrophileRadical donor Hydride relayRadical relay Hydrogen bond acceptorRadical stabiliser Hydrogen bond donorSpectator Leaving groupSteric hindrance Metal ligandUnknown function NucleophileUnspecified steric role ________________________________________________ Function of catalytic residues

65
CMLReact Customisable mark-up language Allows validation Uses dictionary technology Separates content from presentation Open Source BUT still under development

66
An Overview of MACiE and CMLReact

67
Energetics of DHQase Model A

68
TS1 - Proton Transfer

69
TS2 - Dehydration Mattias Blomberg 69/41

70
Model C

71
Model A

72
Model B

73
Model C

74
Models A, B & C

75
MD and QM/MM Calculations on Fosfomycin Resistance Protein A

76
Fosfomycin Resistance Protein A

77
Fosfomycin Resistance Proteins Fosfomycin inhibits the first step in the bacterial cell-wall synthesis (MurA). Mn(II)-dependent soluble glutathione (GSH) transferase. FosA homologues in pathogenic bacteria: FosB and FosX.
-dependent soluble glutathione (GSH) transferase. FosA homologues in pathogenic bacteria: FosB and FosX..")
78
Impact on Pathogens Low toxicity and broad-spectrum activity have resulted in an increased clinical use of fosfomycin Fosfomycin is most commonly used in treatments of lower urinary tract infections Fosfomycin alone or in combination with other drugs could also be useful against resistant Staphylococci and E. Coli, which can give serious infections for hospitalized patients (pneumonia, urinary tract infections, skin infections and bacteraemia).
..")
79
Proposed Mechanism Lys90, Tyr100 and Arg119 mutants have a large effect on the turnover of the enzyme. They are all involved in the stabilization of the phosphonate group (Beharry et al, J Biol Chem, 2005, 17786.) Recent docking and mutation studies indicate that Trp34, Gln36, Tyr39, Ser50, Lys90 and Arg93 are involved in the binding of GSH (Rigsby et al, Arch. Biochem. Biophys, 2007, 277.) Tyr39 has been proposed to participate in the ionization of GSH (Rigsby et al, Arch. Biochem. Biophys, 2007, 277.)
 Recent docking and mutation studies indicate that Trp34, Gln36, Tyr39, Ser50, Lys90 and Arg93 are involved in the binding of GSH (Rigsby et al, Arch. Biochem. Biophys, 2007, 277.) Tyr39 has been proposed to participate in the ionization of GSH (Rigsby et al, Arch. Biochem. Biophys, 2007, 277.).")
80
Docking of GSH in FosA 10 structures from the lowest energy conformations. The GSH thiol is placed in the vicinity of FCN. 30 LGA Dockings using AutoDock 4, 1.5 Å clustering.

81
MD simulations Amber 9. FF03 force field, TIP3P water model. Truncated octahedron > 10 Å of water around the solute. 10 Å cutoff on non-bonding interactions Charges and Force constants for the Mn-centre (His, Glu, Mn, FCN) calculated using Gaussian 03.
 calculated using Gaussian 03..")
82
Backbone RMSD residue 1-268 GS - GSH t (ps)
")
83
Distance GSH(S) – FCN (C) of the different Protonation States of GSH GS - GSH t (ps) GS - Leaves the Binding Pocket
 – FCN (C) of the different Protonation States of GSH GS - GSH t (ps) GS - Leaves the Binding Pocket")
84
MD snapshot of FosA active site

85
Residues Shown to Affect FosA Actvity and Interactions with the Modelled GSH ResidueInteracting with GSHComments Arg93Yes Lys90Yes Ser50NoFCN Tyr39Yes Gln36Yes Trp34Yes Gln91No His64NoMn-ligand Tyr62Yes Cys48NoFCN Tyr128Yes Arg119Yes Trp46NoFCN Tyr65No Ser94NoFCN Glu95Yes Ser98NoFCN Tyr100NoFCN Asp103No His107No Glu110NoMn-ligand Thr9NoFCN Most of the observed changes in FosA activity can be identified with the interactions with FCN or the modelled binding of GSH

86
QM/MM-model of FosA Unrestricted Restricted

87
Preliminary Energetics for FosA

Similar presentations






![Review: Amino Acid Side Chains Aliphatic- Ala, Val, Leu, Ile, Gly Polar- Ser, Thr, Cys, Met, [Tyr, Trp] Acidic (and conjugate amide)- Asp, Asn, Glu, Gln.](/15/4531817/big_thumb.jpg "Review: Amino Acid Side Chains Aliphatic- Ala, Val, Leu, Ile, Gly Polar- Ser, Thr, Cys, Met, [Tyr, Trp] Acidic (and conjugate amide)- Asp, Asn, Glu, Gln.>")












