Download presentation
Presentation is loading. Please wait.

1
The Genetic Code and Transcription
PowerPoint® Lecture Presentation for Concepts of Genetics Ninth Edition Klug, Cummings, Spencer, Palladino Chapter 14 The Genetic Code and Transcription Lectures by David Kass with contributions from John C. Osterman. Copyright © 2009 Pearson Education, Inc.

2
Section 14.1 The genetic code is:
written in linear form – composed of mRNA RNA derived from complementary bases in DNA In mRNA, triplet codons specify 1 amino acid code contains “start” and “stop” signals unambiguous degenerate commaless nonoverlapping nearly universal

3
Section 14.4 Genetic code is degenerate, w/many amino acids specified by more than one codon. Only tryptophan and methionine are encoded by a single codon.

4
Section 14.4 Wobble hypothesis predicts that hydrogen bonding between the codon and anticodon at the third position is subject to modified base-pairing rules.

5
Section 14.4 The genetic code shows order in that chemically similar amino acids often share one or two middle bases in the triplets encoding them.

6
Section 14.4 The initial amino acid incorporated into all proteins is a modified form of methionine—N-formylmethionine (fmet). (in bacteria) AUG is the only codon to encode for methionine. Initiator codon When AUG appears internally in mRNA, an unformylated methionine is inserted into the protein.

7
Section 14.4 Three codons (UAG, UAA, and UGA) serve as termination codons and do not code for any amino acid.
 serve as termination codons and do not code for any amino acid.")
8
Section 14.6 The Genetic Code Is Nearly Universal
Mitochondrial DNA revealed some exceptions to the universal genetic code.

9
Section 14.7 In some viruses, overlapping genes have been identified in which initiation at different AUG positions out of frame with one another leads to distinct polypeptides.

10
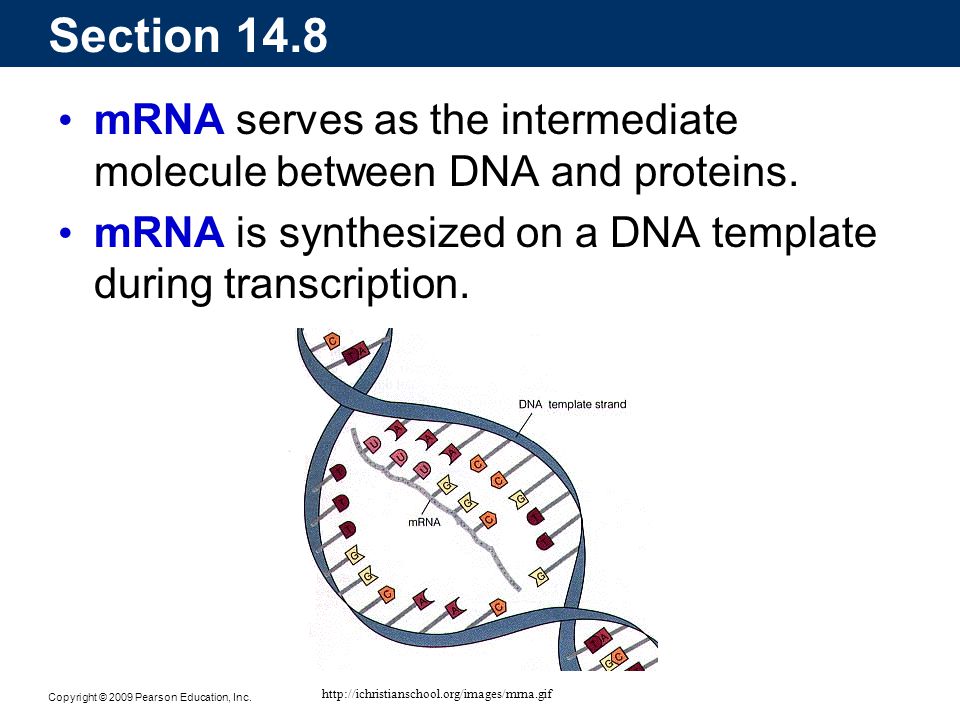
Section 14.8 mRNA serves as the intermediate molecule between DNA and proteins. mRNA is synthesized on a DNA template during transcription.
11
Section 14.10 RNA polymerase directs the synthesis of RNA using a DNA template. No primer is required for initiation The enzyme uses ribonucleotides instead of deoxyribonucleotides.

12
Section 14.10 Transcription begins with template binding by RNA polymerase at a promoter. The s subunit is responsible for promoter recognition (in bacteria).
.")
13
Section 14.10 Transcription begins at the transcription start site, where the DNA double helix is unwound to make the template strand accessible.

14
Section 14.10 E. coli promoters have two consensus sequences, TTGACA and TATAAT, positioned at –35 and –10 with respect to the transcription initiation site.

15
Section 14.10 Once initiation has been completed with the synthesis of the first 8–9 nucleotides, sigma (s) dissociates and elongation proceeds with the core enzyme.
 dissociates and elongation proceeds with the core enzyme.")
16
Section 14.10 At the end of the gene, transcription terminates due to hairpin formation in the RNA. In some cases, termination depends on the rho () termination factor.
 termination factor.")
17
Section 14.11 Transcription in Eukaryotes Differs from Prokaryotic Transcription in Several Ways Occurs in nucleus and is not coupled to translation. Requires chromatin remodeling. In addition to promoters, enhancers also influence transcription regulation. Eukaryotic mRNAs require processing to produce mature mRNAs.

18
Eukaryotes possess three forms of RNA polymerase, each of which transcribes different types of genes. Table RNA Polymerases in Eukaryotes Table 14.7

19
Section 14.11 RNA polymerase II (RNP II) promoters have a core promoter element and promoter and enhancer elements. The TATA box is a core promoter element that binds the TATA-binding protein (TBP) of transcription factor TFIID and determines the start site of transcription. CAAT box
 of transcription factor TFIID and determines the start site of transcription. CAAT box.")
20
Section 14.11 General transcription factors are required for all RNP II mediated transcription and help RNA polymerase II bind to the promoter and initiate transcription.

21
Section 14.11 Heterogeneous nuclear RNA (hnRNA) is posttranscriptionally processed by the addition of a 5' cap and a poly-A tail. Introns are removed by splicing. Exons spliced together.

22
Section 14.12 Introns (intervening sequences) are regions of the initial RNA transcript that are not expressed in the amino acid sequence of the protein. Introns are removed by splicing and the exons (expressed) are joined together in the mature mRNA. The size of the mature mRNA is usually much smaller than that of the initial RNA.
 are regions of the initial RNA transcript that are not expressed in the amino acid sequence of the protein. Introns are removed by splicing and the exons (expressed) are joined together in the mature mRNA. The size of the mature mRNA is usually much smaller than that of the initial RNA.")
23
Figure 14.12

24
Table 13-8 Contrasting Human Gene Size, mRNA Size, and the Number of Introns

25
Group 1 Introns Figure Splicing mechanism involved with group I introns removed from the primary transcript leading to rRNA. The process is one of self-excision involving two transesterification reactions. Figure 14.13

26
Section 14.12 Pre-mRNA introns are spliced out by the spliceosome in a reaction involving the formation of a lariat structure.

27
14.13 Transcription Has Been Visualized by Electron Microscopy
Figure 14.15

Similar presentations



















