Download presentation
Presentation is loading. Please wait.

4
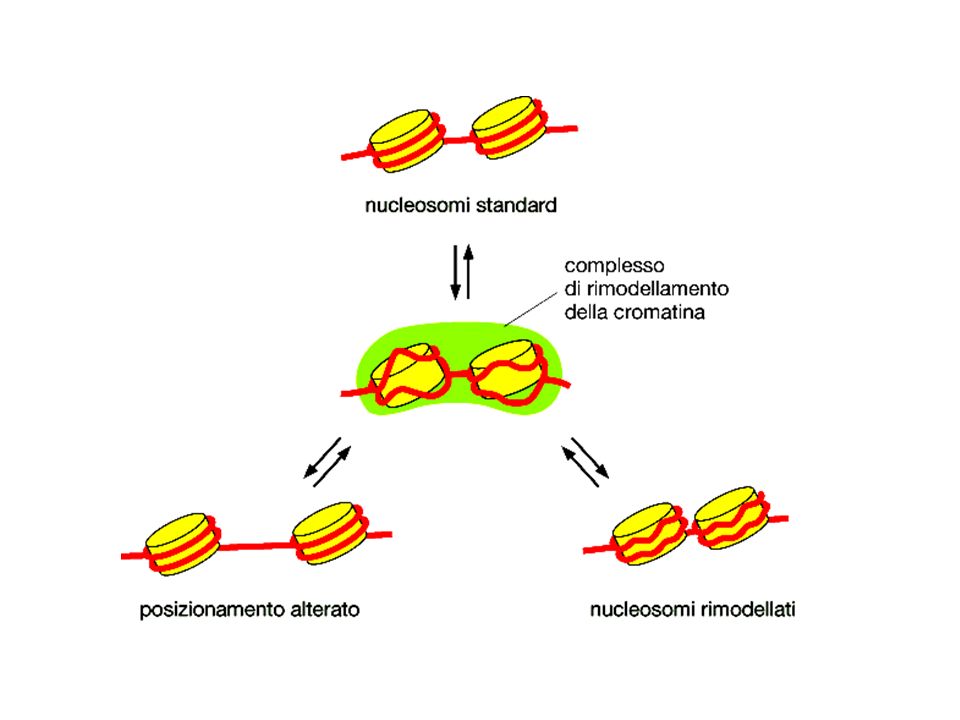
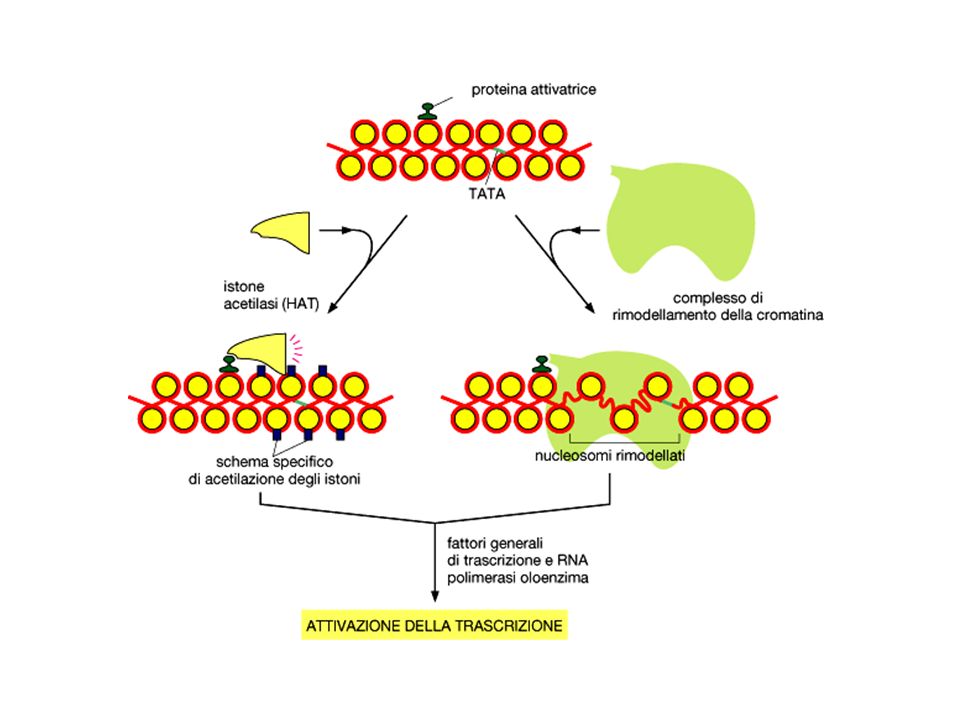
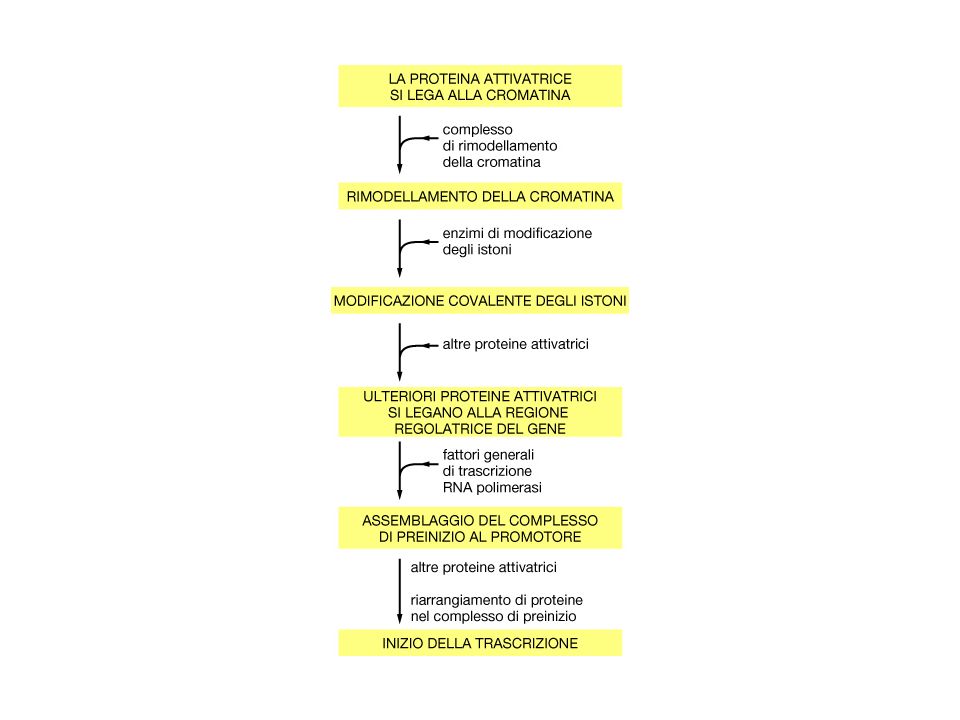
Legame cometitivo al DNA
Mascheramento della superficie di attivazione Interazione con Fattori generali della trascrizione Reclutamento di complessi di rimodellamento della cromatina repressivi Reclutamento di istone deacetilasi

5
HELICASE/ATPase domain
SWI/SNF(yest) ATPase bromodomain (Bromo) histones helicase-SANT-associated (HSA) domain association of the subunit with actin and actin-related proteins slides and ejects nucleosomes at many loci ma non roles in chromatin assembly. ISWI (Droshphila) SANT domain SLIDE domain (SANT-like ISWI), a binds to an unmodified histone tail and DNA optimize nucleosome spacing to promote chromatin assembly and the repression of transcription. certain complexes (NURF) can randomize spacing, which can assist RNAPII activation CHD(xenopus leavis) (chromo-ATPase/helicase-DNA-binding) double chromodomain H3K4 methylation CHD remodelers slide or eject nucleosomes to promote transcription. others have repressive roles, including Mi-2/NuRD complex, which contains histone deacety- lases (HDAC1/2) and methyl CpG-binding do- main (MBD) proteins. INO80(s.Cerevisiae) (inositol requiring 80) family remodelers SWR1-related complexes split” ATPase domain, with a long insertion present in the middle of the ATPase domain, Both yINO80 and ySWR1 complexes also contain actin and Arp4. INO80 has diverse functions, including promoting transcriptional activation and DNA repair. SWR1 is unique in its ability to restructure the nucleosome by removing canonical H2A-H2B dimers and replacing them with H2A.Z-H2B dimers,
 ATPase bromodomain (Bromo) histones. helicase-SANT-associated (HSA) domain association of the subunit with actin and actin-related proteins. slides and ejects nucleosomes at many loci ma non roles in chromatin assembly. ISWI (Droshphila) SANT domain SLIDE domain (SANT-like ISWI), a binds to an unmodified histone tail and DNA. optimize nucleosome spacing to promote chromatin assembly and the repression of transcription. certain complexes (NURF) can randomize spacing, which can assist RNAPII activation. CHD(xenopus leavis) (chromo-ATPase/helicase-DNA-binding) double chromodomain H3K4 methylation. CHD remodelers slide or eject nucleosomes to promote transcription. others have repressive roles, including Mi-2/NuRD complex, which contains histone deacety- lases (HDAC1/2) and methyl CpG-binding do- main (MBD) proteins. INO80(s.Cerevisiae) (inositol requiring 80) family remodelers. SWR1-related complexes. split ATPase domain, with a long insertion present in the middle of the ATPase domain, Both yINO80 and ySWR1 complexes also contain actin and Arp4. INO80 has diverse functions, including promoting transcriptional activation and DNA repair. SWR1 is unique in its ability to restructure the nucleosome by removing canonical H2A-H2B dimers and replacing them with H2A.Z-H2B dimers,")
6
SNF2 ATPase activity ROLE of SNF2/BRM domains Roles of SWI2/SNF2
BROMO domain binds acetylated lysines on histone tails HSA domain protein interactions actin/ARP transcription factors ATPase domain SNF2 ATPase activity change nucleosome position increased regulatory protein access! change nucleosome conformation eject histone octamer displace H2A/H2B dimer Roles of SWI2/SNF2 Role in activation or repression of genes Inducible gene expression: transcription initiation transcription elongation Splicing Repair

7
Roles of ISWI ROLE of ISWI domains nucleosome array formation
chromatin assembly, replication heterochromatin formation transcriptional regulation some PolII, PolI ROLE of ISWI domains SANT/HAND domain contacts histone tails Slide domain linker DNA contact, ’measures’ distance equal spacing of nucleosomes ATPase domain

8
CHD ATPases ROLE of CHD ATPase domains
CHD1: role in chromatin assembly; open chromatin in pluripotent cells NuRD complex also contains Me-DNA binding protein (MBD2) complex connects deacetylation, chromatin remodeling and DNA methylation; repressive function ROLE of CHD ATPase domains CHROMO and PHD domains bind methylated lysines on histone tails modulate activity of remodelers ATPase domain
 complex connects deacetylation, chromatin remodeling. and DNA methylation; repressive function. ROLE of CHD ATPase domains. CHROMO and PHD domains. bind methylated lysines on histone tails. modulate activity of remodelers. ATPase domain.")
9
INO80 SWR1 chromatin assembly DNA repair
interacts with phosphorylated H2A.X (gammaH2A.X) transcription SWR1 H2A exchange with H2A.Z Boundary to heterochromatin spreading transcriptionally poised promoters (together with H3.3) INO80 subfamily complexes have been identified as transcriptional regulators. subfamily are: INO80 and SWR1 in S. cerevisiae; INO80, Snf2-related CBP activator protein (SRCAP) and p400 in mammals; and INO80 and p400 in Drosophila melanogaster. yin yang 1 (YY1; in mam- mals) These trans- cription factors, which are involved in cell proliferation, differentiation and embryonic developmentspecify the genes that are targeted for INO80-mediated chromatin remodelling SWR1- regulated genes in S. cerevisiae are enriched near telomeres, suggesting that the chromatin-remodelling activity of the SWR1 complex might be needed to RuvB-like helicases in the complexesRuvB helicase is a DNA repair factor in bacteria19,20, and thus the presence of RuvB-like subunits in the complexes suggests a role for the INO80 subfamily in DNA damage responses Interestingly, complexes of the INO80 subfamily have a prominent and distinct affinity for the histone variants γ-H2AX and H2AZ. (Phosphorylated H2AX is often referred to as γ-H2AX) γ-H2AX and H2AZ are not found in canonical nucleosomes and function to direct the activities of the INO80 subfamily in specific processes, such as DNA repair and DNA damage checkpoint responses
 transcription. SWR1. H2A exchange with H2A.Z. Boundary to heterochromatin spreading. transcriptionally poised promoters (together with H3.3) INO80 subfamily complexes have been identified as transcriptional regulators. subfamily are: INO80 and SWR1 in S. cerevisiae; INO80, Snf2-related CBP activator protein (SRCAP) and p400 in mammals; and INO80 and. p400 in Drosophila melanogaster. yin yang 1 (YY1; in mam- mals) These trans- cription factors, which are involved in cell proliferation, differentiation and embryonic developmentspecify the genes that are targeted for INO80-mediated chromatin remodelling. SWR1- regulated genes in S. cerevisiae are enriched near telomeres, suggesting that the chromatin-remodelling activity of the SWR1 complex might be needed to. RuvB-like helicases in the complexesRuvB helicase is a DNA repair factor in bacteria19,20, and thus the presence of RuvB-like subunits in the complexes suggests a role for the INO80 subfamily in DNA damage responses. Interestingly, complexes of the INO80 subfamily have a prominent and distinct affinity for the histone variants γ-H2AX and H2AZ. (Phosphorylated H2AX is often referred to as γ-H2AX) γ-H2AX and H2AZ are not found in canonical nucleosomes and function to direct the activities of the INO80 subfamily in specific processes, such as DNA repair and DNA damage checkpoint responses.")
10
the presence of a spacer region that splits the conserved ATPase domain
Deletion of this spacer region in S. cerevisiae Swr1 ATPase results in loss of the association of several subunits from the SWR1 complex, including the RuvB-like Rvb1 and SWI/SNF family have helicase domains in their respec- tive ATPase subunits but do not exhibit helicase activity. However, the S. cerevisiae INO80 complex is unique because it does exhibit in vitro ATP-dependent DNA helicase activity 4. INO80 influences nucleosome eviction a rather dramatic chromatin-remodelling event in which the nucleosomes are completely removed from the DNA, leaving the DNA accessible to participate in nuclear functions without the presence of histones SWR1 complex co-purifies with the histone variant Htz1 (H2AZ in mammals) Histone variants are often incorporated into nucleosomes at site-specific locations to accommodate cell-ular functions, such as transcription and replication, H2AZ has essential functions in a range of cellular processes, such as transcriptional regulation, heterochromatin establishment chromosome segregation, cell cycle progression and DNA damage responses Analyses of chromatin fibres in vitro suggest that H2AZ promotes different conformational states of the nucleosome that might facilitate varied chromatin functions Specifically, the crystal structure of H2AZ-containing nucleosomes suggests that the nucleosome is destabilized by a region that includes the carboxyl-terminal α-helix of H2AZ, which contains a high degree of sequence divergence from H2A45 (BOX 1). Loss of this H2AZ C-terminal region results in poor survival in yeast27 and impaired developmen D.melanogaster 46. this region of H2AZ is also essential for recognition by the SWR1 chromatin-remodelling complex for histone exchange2
 Histone variants are often incorporated into nucleosomes at site-specific locations to accommodate cell-ular functions, such as transcription and replication, H2AZ has essential functions in a range of cellular processes, such as transcriptional regulation, heterochromatin establishment chromosome segregation, cell cycle progression and DNA damage responses. Analyses of chromatin fibres in vitro suggest that H2AZ promotes different conformational states of the nucleosome that might facilitate varied chromatin functions. Specifically, the crystal structure of H2AZ-containing nucleosomes suggests that the nucleosome is destabilized by a region that includes the carboxyl-terminal α-helix of H2AZ, which contains a high degree of sequence divergence from H2A45 (BOX 1). Loss of this H2AZ C-terminal region results in poor survival in yeast27 and impaired developmen. D.melanogaster 46. this region of H2AZ is also essential for recognition by the SWR1 chromatin-remodelling complex for histone exchange2.")
11
regulation of aTP-dependent chromatin remodelling.
mechanisms in combination can generate numerous structural and functional diversities for a single chromatin-remodelling complex. Transcription factors can target the complex for recruitment to specific target genes. The binding of structured DNA, such as three-way and four-way junctions that represent DNA replication and repair intermediates, can also recruit and potentially alter the activity of the complex. Small molecules, can modulate the activity of the complex. subunits in the complex can be post-translationally modified or exchanged to alter the function of the complex. The activity of specific subunits can also be regulated by transient association of the complex with activating factors, Finally, the association of unique chromatin substrates, such as histone variants and post-translationally modified histones, can further specify the chromatin-remodelling mechanisms of the complex.

12
The helicase-like region (H) is highly conserved between the two types of ATPases whereas the flanking sequences are not conserved. (b) Model for monomeric chromatin remodeling ATPases: (i) ATP binding by the helicase domain (H) causes DNA distortion at SHL−2 and stabilization of unpeeled DNA ‘slack’ by the DNA-binding domain (DBD). The H domain binds tightly in this state. (ii) ATP hydrolysis causes translocation of the H domain on the disrupted DNA at SHL−2, generating a DNA loop. (iii) Phosphate release weakens affinity of H domain increasing the probability of falling off. (iv) Dissociation of enzyme allows stochastic collapse of disrupted nucleosome into multiple distinct products. C Model for dimeric chromatin remodeling ATPases: (i) Only one monomer can bind ATP via its H domain at a given time. ATP can rapidly switch between the two monomers. The monomer that contacts the longer flanking DNA hydrolyzes ATP faster. Here this is the black monomer. (ii) ATP hydrolysis causes translocation of the H domain on the DNA at SHL−2, generating a DNA loop. (iii) Phosphate release from the black H domain weakens the affinity of the black H domain but at the same time allows ATP binding to the white H domain switching it to a tight binding state at SHL+2. This allows propagation of the DNA loop without enzyme dissociation. In this state the white DBD does not properly engage the flanking DNA preventing ATP hydrolysis by the white H domain. (iv) Loss of ADP from the black H domain allows the white DBD to engage the flanking DNA. At this stage ATP can rapidly switch between the two monomers and further DNA movement can occur or the enzyme can dissociate resulting in a repositioned nucleosome.
 Model for monomeric chromatin remodeling ATPases: (i) ATP binding by the helicase domain (H) causes DNA distortion at SHL−2 and stabilization of unpeeled DNA ‘slack’ by the DNA-binding domain (DBD). The H domain binds tightly in this state. (ii) ATP hydrolysis causes translocation of the H domain on the disrupted DNA at SHL−2, generating a DNA loop. (iii) Phosphate release weakens affinity of H domain increasing the probability of falling off. (iv) Dissociation of enzyme allows stochastic collapse of disrupted nucleosome into multiple distinct products. C Model for dimeric chromatin remodeling ATPases: (i) Only one monomer can bind ATP via its H domain at a given time. ATP can rapidly switch between the two monomers. The monomer that contacts the longer flanking DNA hydrolyzes ATP faster. Here this is the black monomer. (ii) ATP hydrolysis causes translocation of the H domain on the DNA at SHL−2, generating a DNA loop. (iii) Phosphate release from the black H domain weakens the affinity of the black H domain but at the same time allows ATP binding to the white H domain switching it to a tight binding state at SHL+2. This allows propagation of the DNA loop without enzyme dissociation. In this state the white DBD does not properly engage the flanking DNA preventing ATP hydrolysis by the white H domain. (iv) Loss of ADP from the black H domain allows the white DBD to engage the flanking DNA. At this stage ATP can rapidly switch between the two monomers and further DNA movement can occur or the enzyme can dissociate resulting in a repositioned nucleosome.")
14
WSTF Williams syndrome transcription factor
WINAC-WICH WSTF Williams syndrome transcription factor he known ATP-dependent chromatin remodeling complexes the identified WINAC components. WSTF is a component of SWI/SNF-type WINAC as well as ISWI-type WICH, which is composed of WSTF and hSNF2h (mammalian ISWI ). WSTF is involved in BAZ family family is involved in distinct chromatin remodeling complexes and has distinct roles. WSTF contains a bromodomain and a PHD finger, which have been identified as a chromatin-recognition domain, but the function of this protein had been unknown. Besides, there is another WSTF-containing compelx designated as WICH, which is known to work as an ISWI-type chromatin remodeling complex especially works at DNA replication
. WSTF is involved in BAZ family. family is involved in distinct chromatin remodeling complexes and has distinct roles. WSTF contains a bromodomain and a PHD finger, which have been. identified as a chromatin-recognition domain, but the. function of this protein had been unknown. Besides, there is another WSTF-containing compelx designated. as WICH, which is known to work as an ISWI-type. chromatin remodeling complex especially works at. DNA replication.")
15
Cyclic recruitment of the co-regulator complexes
Recently several types of chromatin remodeling complexes are known to be recruited in cyclically manner to the promoter regions in the process of ligand-dependent transcription mediated by nuclear receptors. WINAC takes part in this cyclic recruitment of co-regulator complexes in the VDR-mediated transcriptional process. Lipophilic ligands, vitamins A and D/steroid hormones, are thought to exert their physiological effects through transcriptional control of target genes via cognate nuclear receptors (NRs) [24].
 [24].")
16
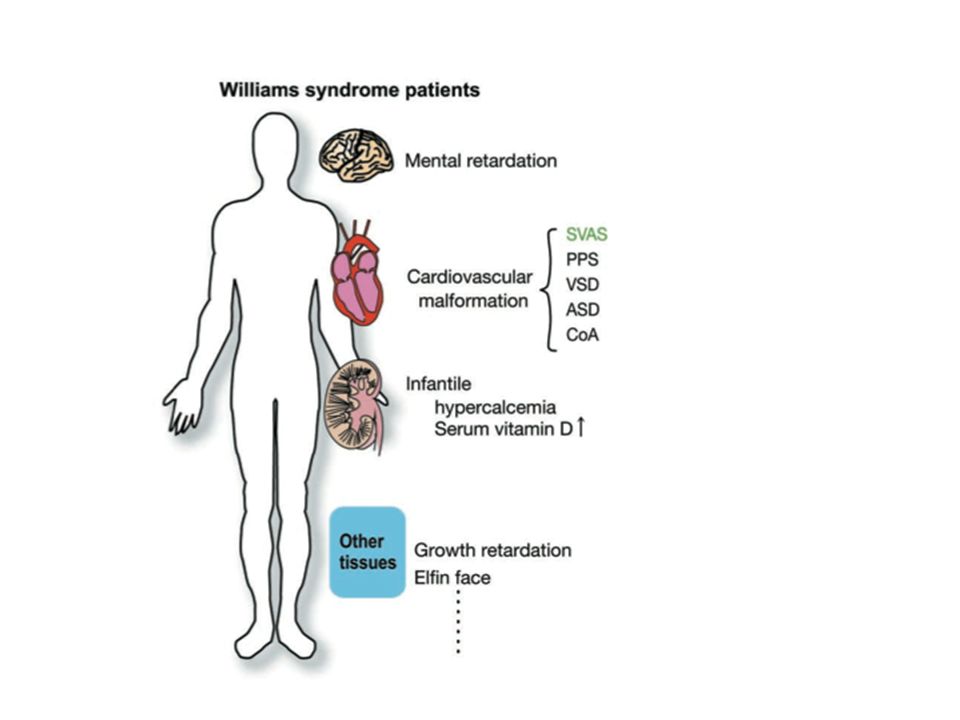
Williams syndrome is a ‘Chromatin remodeling disease’
Some symptoms observed in Williams syndrome patients are attributed to the dysfunction of WINAC or WICH Williams syndrome is caused by the aberrant chromatin remodeling activit WSTF is a chromatin remodeler that is essential for physiological functions of certain sequence-specific transcriptional regulators which are essential for cardiac development. As we found that WSTF works as a component of WINAC in the transcription process, WICH is indispensable for repair of DNA damage, since restored expression of WSTF, together with the WICH component Snf2h, amelio- rated impaired survival after DNA damage in WSTF -/- MEF cells. In conclusion, the WSTF subunit appears to serve as a chromatin remodeler and is a component of two func- tionally-distinct complexes

17
Chromatin remodeling complexes work at various situations to facilitate access of the biological effectors to the target regions of the genome through altering the adjacent chromatin status Considering the roles of the components specific for each chromatin remodeling complex, we believe that WSTF, as a specific component of WINAC, works as a sensor of the various intracellular signalings for turning on its chromatin remodeling activity when required. WSTF was phosphorylated by the stimulation of MAPK cascades in vitro and in vivo . S158 residue in the WAC domain, located close to the N-terminus of WSTF, was iden- tified as a major phosphorylation target. WINAC mediates a pre - viously unknown MAPK-dependent step in epigenetic regulation this MAPK-dependent switching mech anism between the two functionally distinct WSTF- containing complexes might underlie the diverse func tions of WSTF in various nuclear events

21
Properties that distinguish pioneer factors from other transcription factors. Most transcription factors cannot access their target sequences on nucleosomes or in compacted chromatin, yet can do so when binding in a highly cooperative fashion with other transcription factors. Pioneer factors can access their target sequences on nucleosomes and certain forms of compacted chromatin, although not all forms. Pioneer factor binding occurs for a stable period and precedes the binding of other transcription factors.

22
Passive and active roles for pioneer factors in endowing transcriptional competence. In the passive role, stable, prior binding of pioneer factors to a complex regulatory sequence, such as an enhancer element, reduces the number of additional factors that are needed to bind at a later time in order to create an active enhancer. Such “priming” can increase the rapidity of a transcriptional response and is seen during development and in hormonal regulation (Gualdi et al. 1996; Carroll et al. 2005). In the active role, pioneer factors can directly facilitate other factors binding to nucleosomal DNA (Cirillo and Zaret 1999) or open up the local chromatin and thereby indirectly allow other factors to bind (Cirillo et al. 2002).
. In the active role, pioneer factors can directly facilitate other factors binding to nucleosomal DNA (Cirillo and Zaret 1999) or open up the local chromatin and thereby indirectly allow other factors to bind (Cirillo et al. 2002)..")
23
tight chromatin binding
FoxA factors possess features of linker histones and conventional transcription factors. The crystal structures of the DNA-binding domains of linker histone (Ramakrishnan et al. 1993) and FoxA3 (Clark et al. 1993) are shown side by side. The “winged helix” motif is evident in each, consisting of a helix–turn–helix motif (HTH) flanked by wings of polypeptide chain (wing 1 and wing 2) that make minor groove contacts along the long axis of the DNA. Such binding allows other proteins (e.g., core histones) to reside on the other side of the DNA. Like linker histone, the FoxA proteins move much more slowly in chromatin than most other transcription factors, as assessed by fluorescence recovery after photobleaching (FRAP) (Sekiya et al. 2009). Unlike linker histone, FoxA proteins make specific base contacts that direct target site binding and possess an N-terminal trans-activation domain (TAD) (Pani et al. 1992). Also, FoxA proteins possess a C-terminal domain that binds directly to core histone proteins and is necessary for the factor to open chromatin (Cirillo et al. 2002). tight chromatin binding helix–turn–helix motif (HTH)
 and FoxA3 (Clark et al. 1993) are shown side by side. The winged helix motif is evident in each, consisting of a helix–turn–helix motif (HTH) flanked by wings of polypeptide chain (wing 1 and wing 2) that make minor groove contacts along the long axis of the DNA. Such binding allows other proteins (e.g., core histones) to reside on the other side of the DNA. Like linker histone, the FoxA proteins move much more slowly in chromatin than most other transcription factors, as assessed by fluorescence recovery after photobleaching (FRAP) (Sekiya et al. 2009). Unlike linker histone, FoxA proteins make specific base contacts that direct target site binding and possess an N-terminal trans-activation domain (TAD) (Pani et al. 1992). Also, FoxA proteins possess a C-terminal domain that binds directly to core histone proteins and is necessary for the factor to open chromatin (Cirillo et al. 2002). tight chromatin binding. helix–turn–helix motif (HTH)")
24
CANCRO al SENO FoxA1 binding in turn enables binding by Estrogen Receptor and its cofactors, leading to the stimulation of transcription of target genes under estrogen hormonal control.

25
stepwise assembly model for transcription initiation.
Depicted are cooperative chromatin remodeling events (left) induced by ATP-dependent factors (purple) and histone-modifying acetyltransferase activities (red and orange) through their interactions (broken arrows) with each other and some transcriptional regulators (blue circle) that have access (thick arrow) to specific DNA sequences through gene-specific nucleosome positioning effects. The core machinery and some co-factors that act at subsequent steps are depicted as a pool to the right. (B) Such chromatin remodeling events could lead to nucleosome shifting relative to specific target sequences as well as histone acetylation (grey lines), which together likely allow full template accessibility to other transcription factors (blue diamond and Sp1, green hexagon) and the core machinery (right). A large body of biochemical studies suggests that activators and their co-regulators can interact (thick black arrow) with multiple components of the core initiation machinery; however, the in vitro reconstitution of promoter binding and transcription initiation requires a specific ordered assembly (small black arrows) with defined biochemical steps (see text). (C) Activated transcription (black arrow) requires the assembly of a large oligomeric initiation complex and, likely, multiple concerted signals (red arrows) from several gene regulators. Association of RNA processing factors [i.e. capping enzyme (CE, red), elongation factors (EFs, cyan), and splicing factors (SFs, orange)] with the initiation complex requires additional signals including, but not limited to, hyperphosphorylation of RNA Pol II (yellow, purple shading)
 induced by ATP-dependent factors (purple) and histone-modifying acetyltransferase activities (red and orange) through their interactions (broken arrows) with each other and some transcriptional regulators (blue circle) that have access (thick arrow) to specific DNA sequences through gene-specific nucleosome positioning effects. The core machinery and some co-factors that act at subsequent steps are depicted as a pool to the right. (B) Such chromatin remodeling events could lead to nucleosome shifting relative to specific target sequences as well as histone acetylation (grey lines), which together likely allow full template accessibility to other transcription factors (blue diamond and Sp1, green hexagon) and the core machinery (right). A large body of biochemical studies suggests that activators and their co-regulators can interact (thick black arrow) with multiple components of the core initiation machinery; however, the in vitro reconstitution of promoter binding and transcription initiation requires a specific ordered assembly (small black arrows) with defined biochemical steps (see text). (C) Activated transcription (black arrow) requires the assembly of a large oligomeric initiation complex and, likely, multiple concerted signals (red arrows) from several gene regulators. Association of RNA processing factors [i.e. capping enzyme (CE, red), elongation factors (EFs, cyan), and splicing factors (SFs, orange)] with the initiation complex requires additional signals including, but not limited to, hyperphosphorylation of RNA Pol II (yellow, purple shading)")
Similar presentations





 3.Histone code marks active and.>")


 = 6 x 10 9 bp Human genome (in diploid cells) = 6 x 10 9 bp 6 x 10.>")




 - Institute of Genetics and Cytology, School of Life Sciences, School of Life Sciences, ->")








