Download presentation
Presentation is loading. Please wait.

1
Visual Sensation & Perception How do we see?

2
Structure of the eye

3
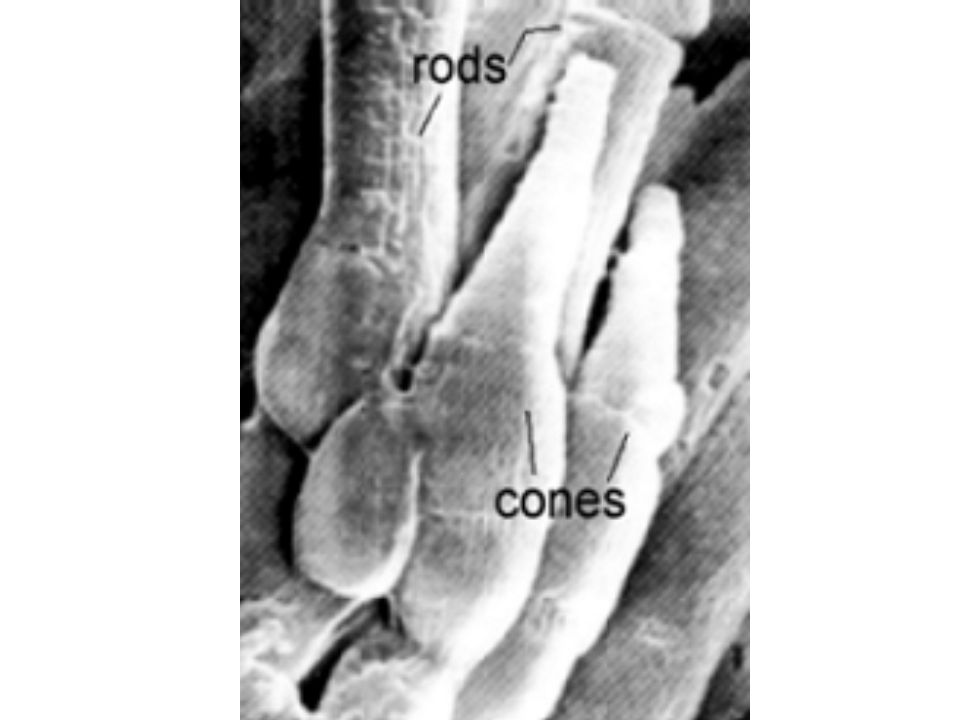
The Retina

5
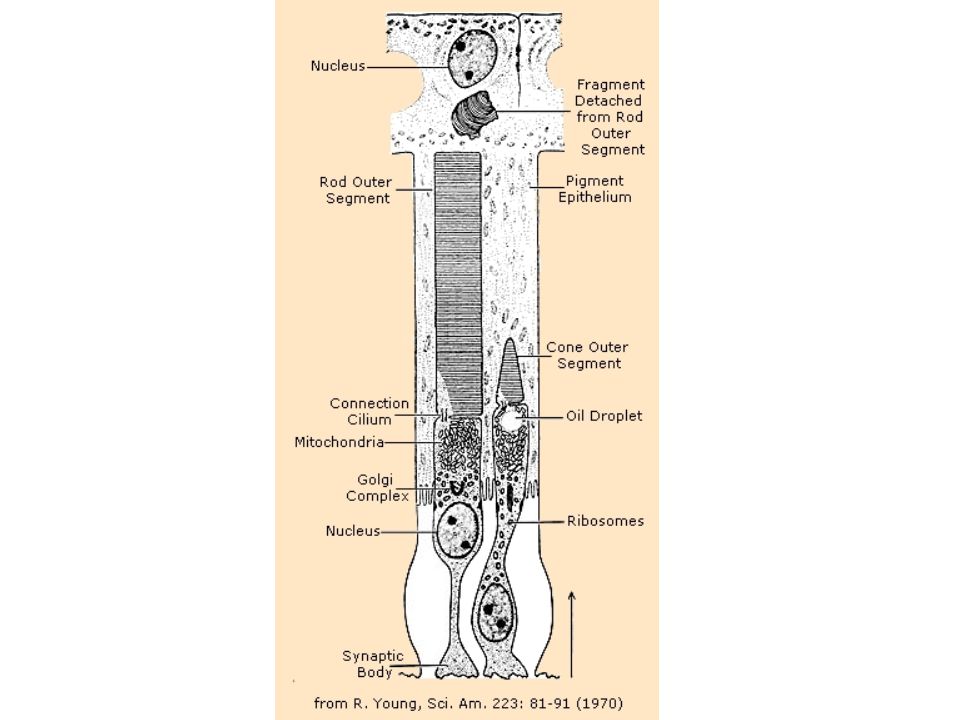
Visual Receptors Rods –Slowly adapting –Black & White vision –120 million; None in fovea Cones –Rapidly adapting –Color vision –6 million; 50,000 in fovea

9
Chemistry of the photoreceptor Light-sensitive protein + retinal (a pigment) absorbs a photon, causing the retinal to change shape. Through a series of enzyme-based chemical reactions, this causes a closing of the Na + channels in the receptor. This has the effect of hyperpolarizing the cell, since sodium can no longer enter. This inhibits the latent activity of the cell, releasing less glutamate into the next layer of the retina - the bipolar cells. This causes one set of bipolar cells to be hyperpolarized (those that had excitatory connections with the receptor) and one set to be depolarized (those that had inhibitory connections).
 and one set to be depolarized (those that had inhibitory connections)..")
10
Retinal Ganglion cells Gather information from many rods and cones across an area of the retina. –How many rods and cones depends on the size of the ganglion cell’s receptive field –The closer to the fovea, the smaller the receptive field. Project out of the eye through the optic nerve, creating a blind spot. 1 million retinal ganglion cells (receiving signals from 125 million receptors).
..")
12
Receptive fields of retinal ganglion cells Center-surround –Most are excitatory center, inhibitory surround. –Some are the opposite

13
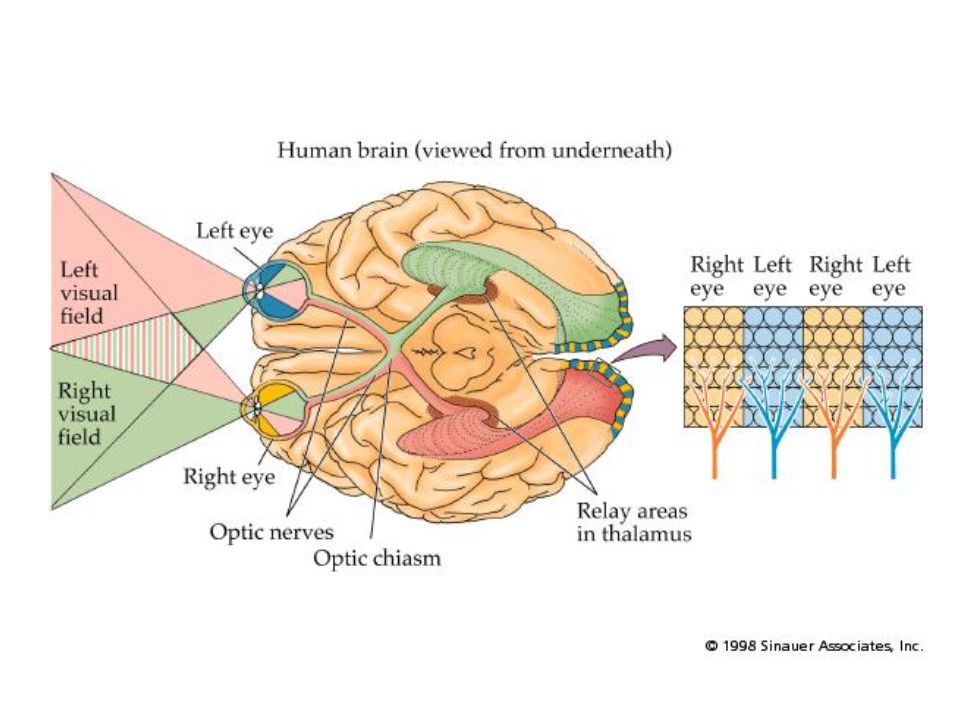
Out of the retina Signals from the two eyes cross over to the opposite brain hemisphere at the optic chiasm. –Not all signals from an eye go to contra-lateral hemisphere. –Which hemisphere the signal goes to is based on which visual hemifield the ganglion cell receives information from.

15
Into the brain Ganglion cells synapse in the lateral geniculate nucleus (LGN) of the thalamus. For comparison, the auditory nerve synapses in the medial geniculate nucleus. The LGN divides the signals into layers depending on which eye they come from, and whether they come from the fovea or not. 1, 4, & 6 from the contralateral eye; 2, 3, and 5 from the ipsilateral eye. 1 and 2 (magnocellular levels) from the fovea. Other layers (parvocellular) from extra-foveal regions. 400,000 cells leave the LGN
 from the fovea. Other layers (parvocellular) from extra-foveal regions. 400,000 cells leave the LGN.")
17
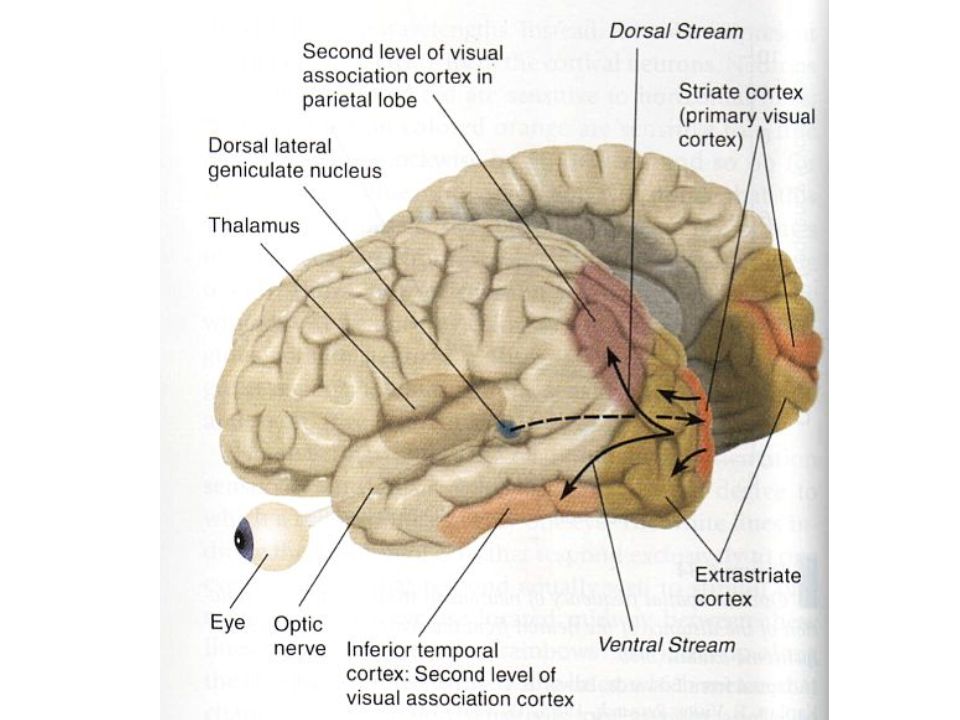
V1 From the LGN, the signals are sent to area V1 in the very back of the occipital lobe. Signals are organized into a retinotopic map based on where on the retina they come from, and which eye they come from. All cells project to layer IV of striate cortex, but M and P cells project to slightly different parts. Furthermore, P cells project from layer IV into layers II and III, creating blob and interblob areas.

19
Feature detectors in V1 The retinotopic map is not simply a light/dark detector. Signals are beginning to be combined into simple feature detectors that can detect lines at various orientations. All of the feature detectors for a particular area of the retina are anatomically organized into a column. A hypercolumn is two columns from corresponding parts of both retinas.

21
Beyond V1 From V1, signals go to area V2 where the combine into more complex features (corners and simple shapes). After V2, the signal splits into two streams of information. –The what stream passes through V3 (which does color detection) into the temporal lobe. –The where stream passes through V4 (which aids with motion detection) into the parietal lobe.
 into the temporal lobe. –The where stream passes through V4 (which aids with motion detection) into the parietal lobe..")
Similar presentations











 2013.6.30.>")


 n 3 types of color receptors l Cones n Differential sensitivity.>")












