Download presentation
Presentation is loading. Please wait.

1
Extensions of the Breeder’s Equation: Permanent Versus Transient Response Response to selection on the variance

2
Permanent Versus Transient Response Considering epistasis and shared environmental values, the single-generation response follows from the midparent-offspring regression R=h 2 S+ S æ 2 z µ æ 2 AA 2 + æ 2 AAA 4 +¢¢¢+æ(E sire ;E o )+æ(E dam ;E o ) ∂ Breeder’s Equation Response from epistasis Response from shared environmental effects Permanent component of response Transient component of response --- contributes to short-term response. Decays away to zero over the long-term

3
Response with Epistasis R=S µ h 2 + æ 2 AA 2æ 2 z ∂ R(1+ø)=S µ h 2 +(1°c) ø æ 2 AA 2æ 2 z ∂ The response after one generation of selection from an unselected base population with A x A epistasis is The contribution to response from this single generation after generations of no selection is c is the average (pairwise) recombination between loci involved in A x A Contribution to response from epistasis decays to zero as linkage disequilibrium decays to zero Response from additive effects (h 2 S) is due to changes in allele frequencies and hence is permanent. Contribution from A x A due to linkage disequilibrium

4
R(t+ø)=th 2 S+(1°c) ø R AA (t) Why unselected base population? If history of previous selection, linkage disequilibrium may be present and the mean can change as the disequilibrium decays More generally, for t generation of selection followed by generations of no selection (but recombination) R AA has a limiting value given by Time to equilibrium a function of c
 R AA has a limiting value given by Time to equilibrium a function of c.")
5
What about response with higher-order epistasis? Fixed incremental difference that decays when selection stops

6
Maternal Effects: Falconer’s dilution model z = G + m z dam + e Direct genetic effect on character G = A + D + I. E[A] = (A sire + A dam )/2 Maternal effect passed from dam to offspring is just A fraction m of the dam’s phenotypic value m can be negative --- results in the potential for a reversed response The presence of the maternal effects means that response is not necessarily linear and time lags can occur in response
/2 Maternal effect passed from dam to offspring is just A fraction m of the dam’s phenotypic value m can be negative --- results in the potential for a reversed response The presence of the maternal effects means that response is not necessarily linear and time lags can occur in response.")
7
Parent-offspring regression under the dilution model In terms of parental breeding values, E(z o jA dam ;A sire ;z dam )= A dam 2 + A sire 2 +mz dam Regression of BV on phenotype A=π A +b Az (z°π z )+e With no maternal effects, b az = h 2 æ A;M =mæ 2 A =(2°m) With maternal effects, a covariance between BV and maternal effect arises, with The resulting slope becomes b Az = h 2 2/(2-m) ¢π z =S dam µ h 2 2°m +m ∂ +S sire h 2 2°m The response thus becomes
= A dam 2 + A sire 2 +mz dam Regression of BV on phenotype A=π A +b Az (z°π z )+e With no maternal effects, b az = h 2 æ A;M =mæ 2 A =(2°m) With maternal effects, a covariance between BV and maternal effect arises, with The resulting slope becomes b Az = h 2 2/(2-m) ¢π z =S dam µ h 2 2°m +m ∂ +S sire h 2 2°m The response thus becomes")
8
109876543210 -0.15 -0.10 -0.05 0.00 0.05 0.10 Generation Cumulative Response to Selection (in terms of S) Response to a single generation of selection Reversed response in 1st generation largely due to negative maternal correlation masking genetic gain Recovery of genetic response after initial maternal correlation decays h 2 = 0.11, m = -0.13 (litter size in mice)
 Response to a single generation of selection Reversed response in 1st generation largely due to negative maternal correlation masking genetic gain Recovery of genetic response after initial maternal correlation decays h 2 = 0.11, m = (litter size in mice)")
9
20151050 -0.5 0.0 0.5 1.0 1.5 Generation Cumulative Response (in units of S) m = -0.25 m = -0.5 m = -0.75 h 2 = 0.35 Selection occurs for 10 generations and then stops
 m = m = -0.5 m = h 2 = 0.35 Selection occurs for 10 generations and then stops")
10
Ancestral Regressions When regressions on relatives are linear, we can think of the response as the sum over all previous contributions For example, consider the response after 3 gens: 8 great-grand parents from generation zero Cov(offspring gen 3 on great-grandparents in gen 0) Selection differential on great-grandparents 2 T-t = number of relatives from gen. t for offspring from gen T T,t = cov(z T,z t )
.")
11
Changes in the Variance under Selection The infinitesimal model --- each locus has a very small effect on the trait. Under the infinitesimal, require many generations for significant change in allele frequencies However, can have significant change in genetic variances due to selection creating linkage disequilibrium Under linkage equilibrium, freq(AB gamete) = freq(A)freq(B) With positive linkage disequilibrium, f(AB) > f(A)f(B), so that AB gametes are more frequent With negative linkage disequilibrium, f(AB) < f(A)f(B), so that AB gametes are less frequent
 = freq(A)freq(B) With positive linkage disequilibrium, f(AB) > f(A)f(B), so that AB gametes are more frequent With negative linkage disequilibrium, f(AB) < f(A)f(B), so that AB gametes are less frequent.")
12
Additive variance with LD: Additive variance is the variance of the sum of allelic effects, Additive variance Genic variance: value of Var(A) in the absence of disequilibrium function of allele frequencies Disequilibrium contribution. Requires covariances between allelic effects at different loci

13
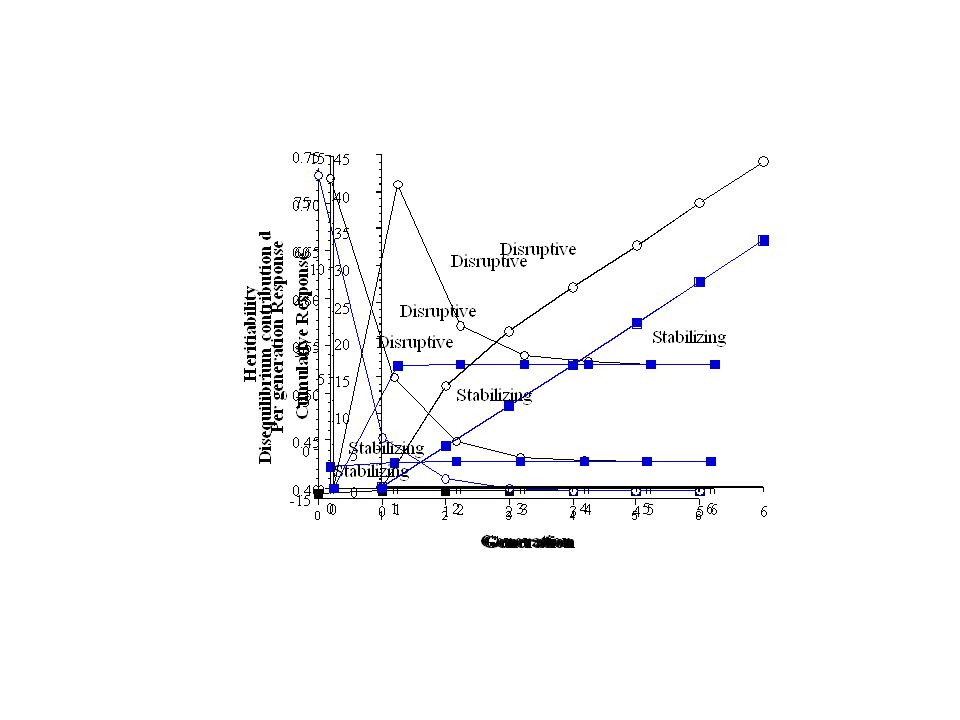
Key: Under the infinitesimal model, no (selection-induced) changes in genic variance 2 a Selection-induced changes in d change 2 A, 2 z, h 2 Dynamics of d: With unlinked loci, d loses half its value each generation (i.e, d in offspring is 1/2 d of their parents,
 changes in genic variance 2 a Selection-induced changes in d change 2 A, 2 z, h 2 Dynamics of d: With unlinked loci, d loses half its value each generation (i.e, d in offspring is 1/2 d of their parents,")
14
Dynamics of d: Computing the effect of selection in generating d Consider the parent-offspring regression Taking the variance of the offspring given the selected parents gives Change in variance from selection

15
Change in d = change from recombination plus change from selection Recombination Selection + = In terms of change in d, This is the Bulmer Equation (Michael Bulmer), and it is akin to a breeder’s equation for the change in variance At the selection-recombination equilibrium,
, and it is akin to a breeder’s equation for the change in variance At the selection-recombination equilibrium,")
16
Application: Egg Weight in Ducks Rendel (1943) observed that while the change mean weight weight (in all vs. hatched) as negligible, but their was a significance decrease in the variance, suggesting stabilizing selection Before selection, variance = 52.7, reducing to 43.9 after selection. Heritability was h 2 = 0.6 e d= e h 4 e ±(æ 2 z ) =0.6 4 (43.9 - 52.7) = -3.2 Var(A) = 0.6*52.7= 31.6. If selection stops, Var(A) is expected to increase to 31.6+3.2= 34.8 Var(z) should increase to 55.9, giving h 2 = 0.62
 as negligible, but their was a significance decrease in the variance, suggesting stabilizing selection Before selection, variance = 52.7, reducing to 43.9 after selection. Heritability was h 2 = 0.6 e d= e h 4 e ±(æ 2 z ) =0.6 4 ( ) = -3.2 Var(A) = 0.6*52.7= If selection stops, Var(A) is expected to increase to = 34.8 Var(z) should increase to 55.9, giving h 2 =")
17
Contribution of within- vs. between-family effects to Var(A) The total additive variance arises from two sources: differences between the mean BVs of families and variation of BVs within families When no LD is present, both these sources contribute equally, Var(A)/2. What happens when LD present? Consider parent-offspring regression in BA The within-family (or mendelian segregation variance) is simply the genic variance and is a constant (if allele frequencies not changing). LD is a function the between-family variance in BV When LD < 0, families are more similar than expected, When LD > 0, families are more dissimilar
 The total additive variance arises from two sources: differences between the mean BVs of families and variation of BVs within families When no LD is present, both these sources contribute equally, Var(A)/2. What happens when LD present. Consider parent-offspring regression in BA The within-family (or mendelian segregation variance) is simply the genic variance and is a constant (if allele frequencies not changing). LD is a function the between-family variance in BV When LD < 0, families are more similar than expected, When LD > 0, families are more dissimilar.")
18
Specific models of selection-induced changes in variances Proportional reduction model: constant fraction k of variance removed Bulmer equation simplifies to Closed-form solution to equilibrium h 2

20
Equilibrium h 2 under direction truncation selection

21
Directional truncation selection

22
Changes in the variance = changes in h 2 and even S (under truncation selection) R(t) = h 2 (t) S(t)
 R(t) = h 2 (t) S(t)")
23
In Class Problem You are selecting the upper 5% of a trait with h 2 = 0.75 and z 2 = 100 initially in linkage equilibrium Compute the response over 3 populations also compute d(t), h 2 (t), z 2 (t), and S(t) Compare the total 3 generations of response with the result from the standard breeder’s equation
, h 2 (t), z 2 (t), and S(t) Compare the total 3 generations of response with the result from the standard breeder’s equation")
24
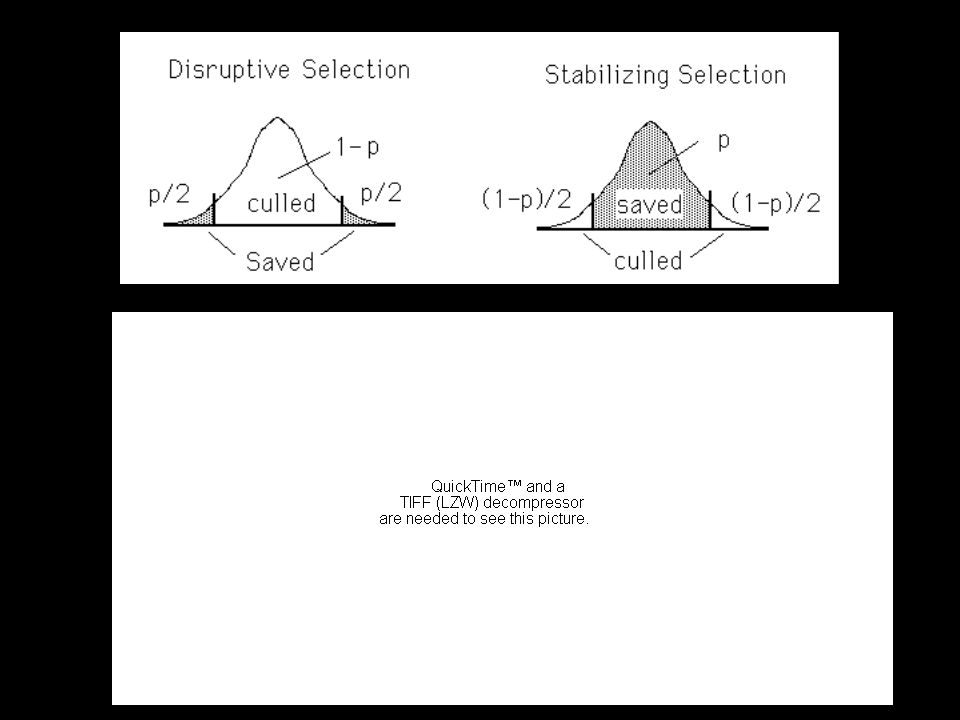
Selection can also focus entirely on the variance (stabilizing & disruptive selection)
")
25
Disruptive selection inflates the variance after selection, generating positive d Stabilizing selection deflates the variance after selection, generating negative d

27
In class problem #2 You have a trait with phenotypic variance 100, heritability 0.4, d(0) = 0 Compare d, h 2 and Var(A) after four generations of truncation selection (p=0.1) vs. double truncation selection (p=0.05 on both tails)
.")
Similar presentations


p 1 p 4 +rp 2 p 3 p 2 ’ =p 2 2 +p 1 p 2 +p 2 p 4 +rp 1 p 4 +(1-r)p 2 p 3 p 3 ’ =p 3 2 +p 3 p.>")


 V p – the total variance.>")


=b T æ(z;z)b=b T Pbæ 2 A I =æ A (b T z;b T z)=b T æ A (z;z)b=b T Gb h 2 I = æ.>")
 Linkage means A) Alleles at different loci are independent B) Alleles at different loci are physically close to each other and on the same chromosome.>")
 controlled by a single locus, with only two alleles: Discrete Variation.>")






 compromise predictions of selection response by changing allele frequencies and generating.>")

 Fragility of covariances to allele frequency.>")