Download presentation
Presentation is loading. Please wait.

1
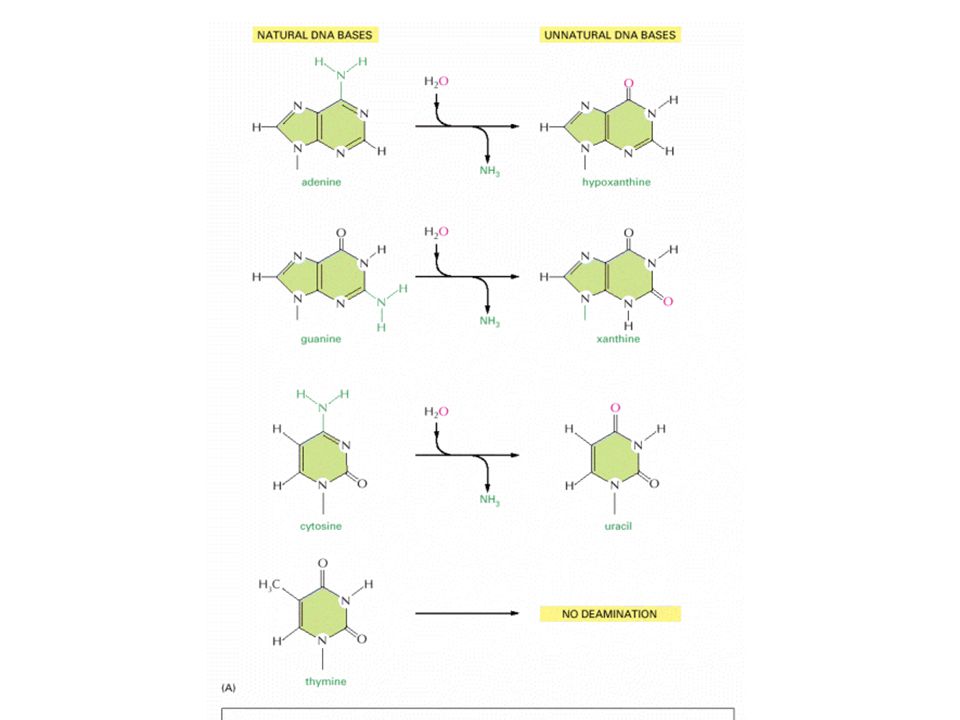
A summary of spontaneous alterations likely to require DNA repair. The sites on each nucleotide that are known to be modified by spontaneous oxidative damage (red arrows), hydrolytic attack (blue arrows), and uncontrolled methylation by the methyl group donor S-adenosylmethionine (green arrows) are shown, with the width of each arrow indicating the relative frequency of each event.
, hydrolytic attack (blue arrows), and uncontrolled methylation by the methyl group donor S-adenosylmethionine (green arrows) are shown, with the width of each arrow indicating the relative frequency of each event..")
3
Depurination and deamination. These two reactions are the most frequent spontaneous chemical reactions known to create serious DNA damage in cells. Depurination can release guanine (shown here), as well as adenine, from DNA. The major type of deamination reaction (shown here) converts cytosine to an altered DNA base, uracil, but deamination occurs on other bases as well. These reactions take place on double-helical DNA; for convenience, only one strand is shown.
, as well as adenine, from DNA. The major type of deamination reaction (shown here) converts cytosine to an altered DNA base, uracil, but deamination occurs on other bases as well. These reactions take place on double-helical DNA; for convenience, only one strand is shown..")
5
Deamination of (a) cytosine and (b) 5-methylcytosine
 cytosine and (b) 5-methylcytosine")
7
The loss of a purine residue (guanine) from a single strand of DNA. The sugar- phosphate backbone is left intact.

9
Alkylation-induced specific mispairing. The alkylation (in this case, EMS-generated ethylation) of the O-6 position of guanine, as well as the O-4 position of thymine, can lead to direct mispairing with thymine and guanine, respectively, as shown here. In bacteria, where mutations have been analyzed in great detail, the principal mutations detected are G·C → A·T transitions, indicating that the O-6 alkylation of guanine is most relevant to mutagenesis
 of the O-6 position of guanine, as well as the O-4 position of thymine, can lead to direct mispairing with thymine and guanine, respectively, as shown here. In bacteria, where mutations have been analyzed in great detail, the principal mutations detected are G·C → A·T transitions, indicating that the O-6 alkylation of guanine is most relevant to mutagenesis.")
10
Repair of a UV-induced pyrimidine photodimer by a photoreactivating enzyme, or photolyase. The enzyme recognizes the photodimer (here, a thymine dimer) and binds to it. When light is present, the photolyase uses its energy to split the dimer into the original monomers
 and binds to it. When light is present, the photolyase uses its energy to split the dimer into the original monomers.")
11
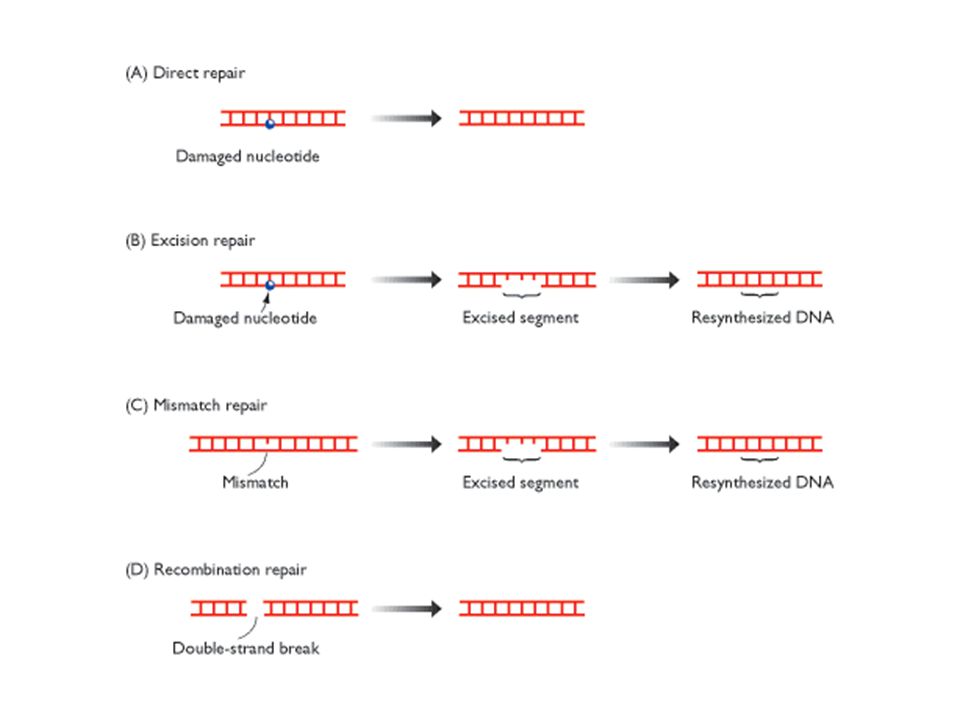
How chemical modifications of nucleotides produce mutations. (A) Deamination of cytosine, if uncorrected, results in the substitution of one base for another when the DNA is replicated. As shown in, deamination of cytosine produces uracil. Uracil differs from cytosine in its base-pairing properties and preferentially base-pairs with adenine. The DNA replication machinery therefore adds an adenine when it encounters a uracil on the template strand. (B) Depurination, if uncorrected, can lead to either the substitution or the loss of a nucleotide pair. When the replication machinery encounters a missing purine on the template strand, it may skip to the next complete nucleotide as illustrated here, thus producing a nucleotide deletion in the newly synthesized strand. Many other types of DNA damage also produce mutations when the DNA is replicated if left uncorrected.
 Deamination of cytosine, if uncorrected, results in the substitution of one base for another when the DNA is replicated. As shown in, deamination of cytosine produces uracil. Uracil differs from cytosine in its base-pairing properties and preferentially base-pairs with adenine. The DNA replication machinery therefore adds an adenine when it encounters a uracil on the template strand. (B) Depurination, if uncorrected, can lead to either the substitution or the loss of a nucleotide pair. When the replication machinery encounters a missing purine on the template strand, it may skip to the next complete nucleotide as illustrated here, thus producing a nucleotide deletion in the newly synthesized strand. Many other types of DNA damage also produce mutations when the DNA is replicated if left uncorrected..")
12
Uracil Repair. Uracil bases in DNA, formed by the deamination of cytosine, are excised and replaced by cytosine.

13
Examples of human DNA glycosylases DNADNA glycosylaseSpecific for MBD4Uracil MPGEthenoadenine, hypoxanthine, 3- methyladenine NTH1Cytosine glycol, dihydrouracil, formamidopyrimidine, thymine glycol OGG1Formamidopyrimidine, 8-oxoguanine SMUG1Uracil TDGEthenocytosine, uracil UNGUracil, 5-hydroxyuracil

14
Long patch mismatch repair in Escherichia coli

15
Methylation of newly synthesized DNA in Escherichia coli does not occur immediately after replication, providing a window of opportunity for the mismatch repair proteins to recognize the daughter strands and correct replication errors.

16
Outline of the events involved during nucleotide excision repair in eukaryotes. The endonucleases that remove the damaged region make cuts specifically at the junction between single-stranded and double-stranded regions of a DNA molecule. The DNA is therefore thought to melt either side of the damaged nucleotide, as shown in the diagram, possibly as a result of the helicase activity of TFIIH.

17
Model of mismatch repair by the E. coli MutHLS system. This repair system operates soon after incorporation of a wrong base, before the newly synthesized daughter strand becomes methylated. MutH binds specifically to a hemimethylated GATC sequence, and MutS binds to the site of a mismatch. Binding of MutL protein simultaneously to MutS and to a nearby MutH activates the endonuclease activity of MutH, which then cuts the unmethylated (daughter) strand in the GATC sequence. A stretch of the daughter strand containing the mispaired base is excised, followed by gap repair and ligation and then methylation of the daughter strand
 strand in the GATC sequence. A stretch of the daughter strand containing the mispaired base is excised, followed by gap repair and ligation and then methylation of the daughter strand.")
18
Structure of DNA-Repair Enzyme. A complex between the DNA-repair enzyme AlkA and an analog of an apurinic site. Note that the damaged base is flipped out of the DNA double helix into the active site of the enzyme for excision

19
Excision Repair. Repair of a region of DNA containing a thymine dimer by the sequential action of a specific excinuclease, a DNA polymerase, and a DNA ligase. The thymine dimer is shown in blue, and the new region of DNA is in red.

20
Mismatch Repair. DNA mismatch repair in E. coli is initiated by the interplay of MutS, MutL, and MutH proteins. A G-T mismatch is recognized by MutS. MutH cleaves the backbone in the vicinity of the mismatch. A segment of the DNA strand containing the erroneous T is removed by exonuclease I and synthesized anew by DNA polymerase III.

21
Short patch nucleotide excision repair in Escherichia coli. The damaged nucleotide is shown distorting the helix because this is thought to be one of the recognition signals for the UvrAB trimer that initiates the short patch process. See the text for details of the events occurring during the repair pathway.

22
A model for strand-directed mismatch repair in eucaryotes. (A) The two proteins shown are present in both bacteria and eucaryotic cells: MutS binds specifically to a mismatched base pair, while MutL scans the nearby DNA for a nick. Once a nick is found, MutL triggers the degradation of the nicked strand all the way back through the mismatch. Because nicks are largely confined to newly replicated strands in eucaryotes, replication errors are selectively removed. In bacteria, the mechanism is the same, except that an additional protein in the complex (MutH) nicks unmethylated (and therefore newly replicated) GATC sequences, thereby beginning the process illustrated here. (B) The structure of the MutS protein bound to a DNA mismatch. This protein is a dimer, which grips the DNA double helix as shown, kinking the DNA at the mismatched base pair. It seems that the MutS protein scans the DNA for mismatches by testing for sites that can be readily kinked, which are those without a normal complementary base pair
 The two proteins shown are present in both bacteria and eucaryotic cells: MutS binds specifically to a mismatched base pair, while MutL scans the nearby DNA for a nick. Once a nick is found, MutL triggers the degradation of the nicked strand all the way back through the mismatch. Because nicks are largely confined to newly replicated strands in eucaryotes, replication errors are selectively removed. In bacteria, the mechanism is the same, except that an additional protein in the complex (MutH) nicks unmethylated (and therefore newly replicated) GATC sequences, thereby beginning the process illustrated here. (B) The structure of the MutS protein bound to a DNA mismatch. This protein is a dimer, which grips the DNA double helix as shown, kinking the DNA at the mismatched base pair. It seems that the MutS protein scans the DNA for mismatches by testing for sites that can be readily kinked, which are those without a normal complementary base pair.")
23
Model for mismatch repair in E. coli. Because DNA is methylated by enzymatic reactions that recognize the A in a GATC sequence, directly after DNA replication the newly synthesized strand will not be methylated. The “hemimethylated” DNA duplex serves as a recognition point for the mismatch-repair system in discerning the old from the new strand. Here a G·T mismatch is shown. The mismatch- repair system can recognize and bind to this mismatch, determine the correct (old) strand because it is the methylated strand of a hemimethylated duplex, and then excise the mismatched base from the new strand. Repair synthesis restores the normal base pair
 strand because it is the methylated strand of a hemimethylated duplex, and then excise the mismatched base from the new strand. Repair synthesis restores the normal base pair.")
24
DNA polymerase III, shown in blue, stops at a non-coding lesion, such as the T·C photodimer shown here, generating single-stranded regions that attract the Ssb protein (dark purple) and RecA (light purple), which forms filaments. The presence of RecA filaments helps to signal the cell to synthesize UmuD (red circles), which is cleaved by RecA to yield UmuD′ (pink circles) and UmuC (yellow ovals). The UmuC is recruited to form a complex with UmuD′ that permits DNA polymerase III to proceed past the blocking lesion.
, which is cleaved by RecA to yield UmuD′ (pink circles) and UmuC (yellow ovals). The UmuC is recruited to form a complex with UmuD′ that permits DNA polymerase III to proceed past the blocking lesion..")
30
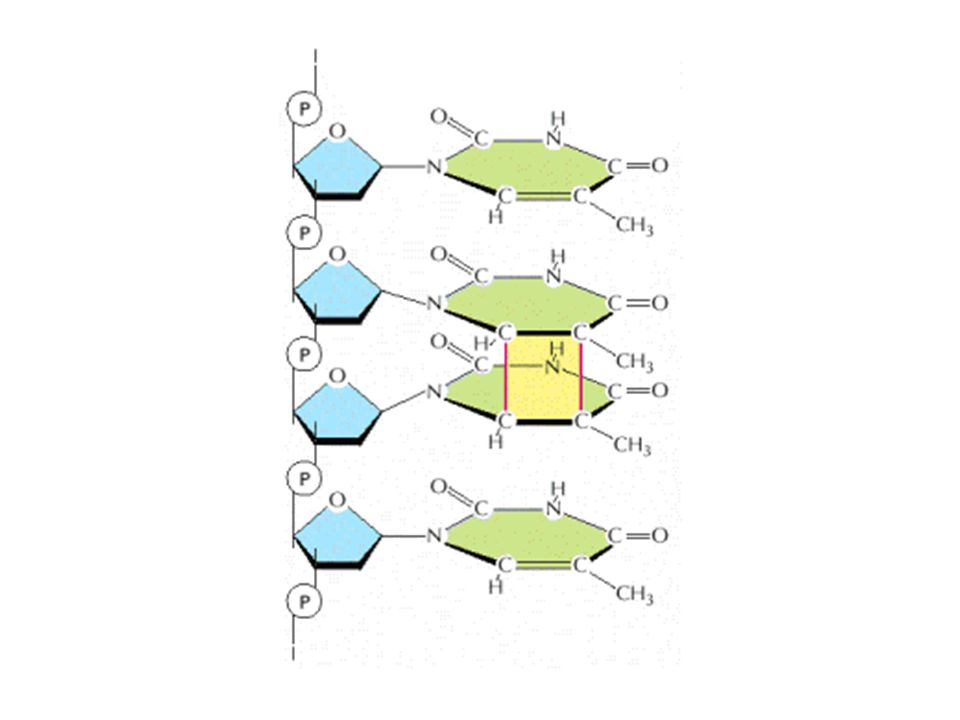
Structure of a cyclobutane pyrimidine dimer. Ultraviolet light stimulates the formation of a four-membered cyclobutyl ring (green) between two adjacent pyrimidines on the same DNA strand by acting on the 5,6 double bonds. (b) Structure of the 6-4 photo-product. The structure forms most prevalently with 5′-C-C-3′ and 5′-T-C-3′, between the C-6 and C-4 positions of two adjacent pyrimidines, causing a significant perturbation in local structure of the double helix.
 between two adjacent pyrimidines on the same DNA strand by acting on the 5,6 double bonds. (b) Structure of the 6-4 photo-product. The structure forms most prevalently with 5′-C-C-3′ and 5′-T-C-3′, between the C-6 and C-4 positions of two adjacent pyrimidines, causing a significant perturbation in local structure of the double helix..")
31
The SOS response of Escherichia coli

32
Two different types of end-joining for repairing double-strand breaks. (A) Nonhomologous end-joining alters the original DNA sequence when repairing broken chromosomes. These alterations can be either deletions (as shown) or short insertions. (B) Homologous end-joining is more difficult to accomplish, but is much more precise.
 Nonhomologous end-joining alters the original DNA sequence when repairing broken chromosomes. These alterations can be either deletions (as shown) or short insertions. (B) Homologous end-joining is more difficult to accomplish, but is much more precise..")
Similar presentations















 Space-filling model C T A A T C G GC A C G A T A T AT T A C T A 0.34 nm 3.4 nm.>")
 If any mismatch escapes the proof reading mechanisms it will cause distortion of the.>")








