Download presentation
Presentation is loading. Please wait.

1
Fundamentals of protein and nucleic acid structure Lecture 2 Structural Bioinformatics Dr. Avraham Samson 81-871

2
Tree of Life All known life forms use the same building blocks suggesting there was a common ancestor

3
The tree of life is not that simple! Gene transfer across kingdoms continually occurs (cyanobacteria became chlorophylls, proteobacteria mitochondria, viruses)
.")
4
Inside living cells

5
29 atoms of life H, O, C, N, P, S Most common elements: H, O, C, N, P, S (97% of organism weight) Most common ions: Ca 2+, K +, Na +, Mg 2+, Cl -
 Most common ions: Ca 2+, K +, Na +, Mg 2+, Cl -")
6
Forces affecting structure: H-bonding Van der Waals Electrostatics Hydrophobicity Disulfide Bridges 150° < θ < 180° d 2.6 Å < d < 3.1Å

7
Forces affecting structure: H-bonding Van der Waals Electrostatics Hydrophobicity Disulfide Bridges Repulsion דחייה Attraction משיכה d 3 Å < d < 4Å

8
Forces affecting structure: H-bonding Van der Waals Electrostatics Hydrophobicity Disulfide Bridges “IONIC BOND” יוני קשר “SALT BRIDGE” מלח גשר E = Energy k = constant D = Dielectric constant (vacuum = 1; H 2 O = 80) q 1 & q 2 = electronic charges (Coulombs) r = distance (Å) Coulomb’s law d d = 2.8 Å
 q 1 & q 2 = electronic charges (Coulombs) r = distance (Å) Coulomb’s law d d = 2.8 Å")
9
Forces affecting structure: H-bonding Van der Waals Electrostatics Hydrophobicity Disulfide Bridges

10
Forces affecting structure: H-bonding Van der Waals Electrostatics Hydrophobicity Disulfide Bridges 10 Other names: cystine bridge disulfide bridge Hair contains lots of disulfide bonds which are broken and reformed by heat

11
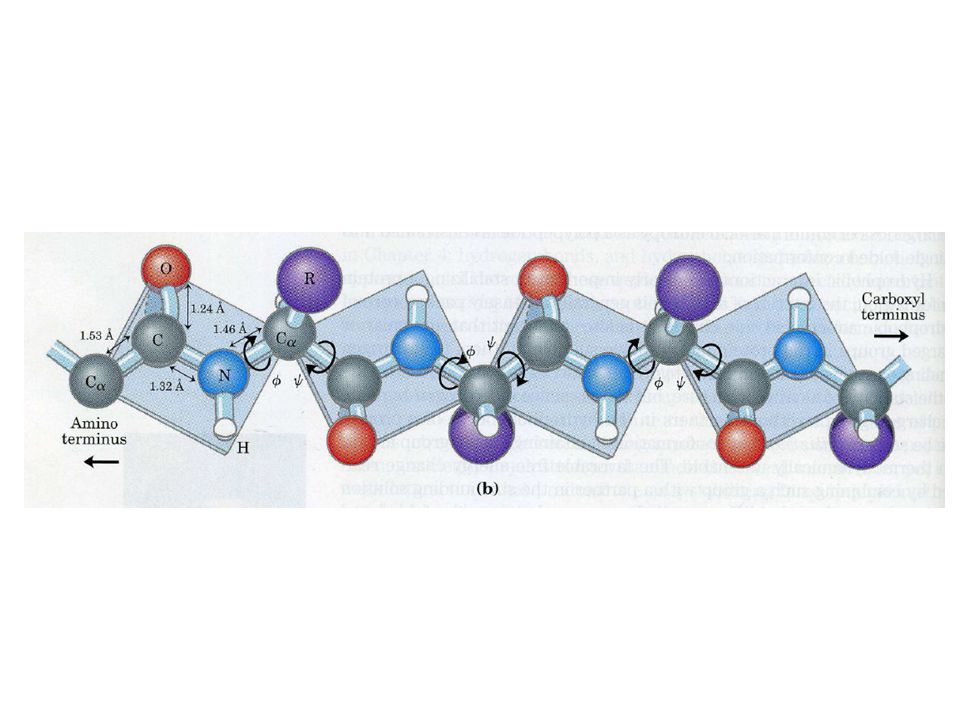
Levels of protein structure Primary: Amino acid sequence Secondary: Local fold pattern of small subsequence Tertiary: Fold of entire protein chain Quaternary: Complex of multiple chains

12
Primary (1 o ) structure The amino acid sequence is the primary structure
 structure The amino acid sequence is the primary structure")
13
13 Primary (1 o ) structure 20 amino acids
 structure 20 amino acids")
14
14 Primary (1 o ) structure
 structure")
15
15 Primary (1 o ) structure
 structure")
16
16 pK a = pH of 50% dissociation

17
17 Amino acid nomenclature Primary (1 o ) structure
 structure")
18
18 Amino acids polymerize through peptide bonds to form polypeptides Primary (1 o ) structure
 structure")
19
19 N-terminal is the start of a polypeptide chain Amino acids are also called residues Primary (1 o ) structure
 structure")
20
20 שלד backbone side chains צדדי שייר Primary (1 o ) structure
 structure")
21
21 Post translational modification are important because they can change the function of proteins (i.e. phosphorylation, acetylation, hydroxylation, carbohydrate and lipid modifications) N-terminal acetylation hydroxyproline O-phosphotyrosine γ-carboxyglutamate Primary (1 o ) structure
 N-terminal acetylation hydroxyproline O-phosphotyrosine γ-carboxyglutamate Primary (1 o ) structure.")
22
22 Amino acids chirality Enantiomers –mirror images - תמונת ראי - אננטיומרים chiral center C כיראלי מרכז Dextro-Laevus in Latin

23
23 Most proteins: only L amino acids איך לקבוע קונפיגורציה L ו D- סדר את האטומים לפי עדיפות על פי ה כללים הבאים כלל.1 אטום עם מספר אטומי יותר גבוה בעל סדר עדיפות יותר גבוה. (I > Cl > O > N> C > H) כלל.2 אם האטומים זהים, העדיפות על פי האטומים המתמירים (C(CH 3 ) 3 > CH(CH 3 ) 2 > CH 2 CH 3 > CH 3 ) שים את האטום עם סדר העדיפות הכי נמוך מאחור. קבע את כיוון החץ, מסדר עדיפות הכי גבוה לככוון הכי נמוך. אם החץ : עם כיוון השעון קונפיגורציה D נגד כיוון השעון קונפיגורציה L רמז יותר קל בשביל לזכור :) CORN תירס ( C chiral center is L configuration
 כלל.2 אם האטומים זהים, העדיפות על פי האטומים המתמירים (C(CH 3 ) 3 > CH(CH 3 ) 2 > CH 2 CH 3 > CH 3 ) שים את האטום עם סדר העדיפות הכי נמוך מאחור. קבע את כיוון החץ, מסדר עדיפות הכי גבוה לככוון הכי נמוך. אם החץ : עם כיוון השעון קונפיגורציה D נגד כיוון השעון קונפיגורציה L רמז יותר קל בשביל לזכור :) CORN תירס ( C chiral center is L configuration.")
24
Bond angles

25
25 Torsion angles , φ, and ψ Unlike , the two backbone dihedral angles φ and ψ are free to rotate This rotation freedom allows protein folding Dihedral angles: -180 o < φ < +180 o -180 o < ψ < +180 o is 0 o or 180 o

26
26 Peptide bond is planar Cα, C, O, N, H, Cα all lie in the same plane

27
27 Torsion angle ( ) is usually trans Steric hindrance Question: What other residues can be cis?
 is usually trans Steric hindrance Question: What other residues can be cis")
28
28 Except for X-Pro bond in which cis is preferred Steric hindrance allows both cis and trans (4:1 ratio) Steric hindrance
 Steric hindrance")
30
30 Ramachandran Diagrams Steric hindrance dictates torsion angle preference Ramachandran plot show preferred regions of φ and ψ dihedral angles Preferred regions

31
Secondary (2 o ) structure Helices -helix 3 10 -helix -helix Sheets Antiparallel Parallel Turns and Unstructured Hydrogen bond: i→i+4 α-helix is the most common 3.6 residues per turn (number of residues in one full rotation of 360°) 5.4 Å pitch (translation along axis for one full rotation of 360°)
 structure Helices -helix helix -helix Sheets Antiparallel Parallel Turns and Unstructured Hydrogen bond: i→i+4 α-helix is the most common 3.6 residues per turn (number of residues in one full rotation of 360°) 5.4 Å pitch (translation along axis for one full rotation of 360°)")
32
Secondary (2 o ) structure Helices -helix 3 10 -helix -helix Sheets Antiparallel Parallel Turns and Unstructured Hydrogen bond: i→i+3 3 10 -helices are rare in proteins: 3.1 residues per turn (number of residues in one full rotation of 360°) 6 Å pitch (translation along axis for one full rotation of 360°)
 structure Helices -helix helix -helix Sheets Antiparallel Parallel Turns and Unstructured Hydrogen bond: i→i helices are rare in proteins: 3.1 residues per turn (number of residues in one full rotation of 360°) 6 Å pitch (translation along axis for one full rotation of 360°)")
33
Secondary (2 o ) structure Helices -helix 3 10 -helix -helix Sheets Antiparallel Parallel Turns and Unstructured Hydrogen bond: i→i+5 π -helices are rare in proteins 4.3 residues per turn (number of residues in one full rotation of 360°) 6 Å pitch (translation along axis for one full rotation of 360°)
 structure Helices -helix helix -helix Sheets Antiparallel Parallel Turns and Unstructured Hydrogen bond: i→i+5 π -helices are rare in proteins 4.3 residues per turn (number of residues in one full rotation of 360°) 6 Å pitch (translation along axis for one full rotation of 360°)")
34
34 Hydrogen bonding in helices CO of residue (n) forms an h-bond with NH of residue: – (n+4) → α-helix – (n+3) → 3 10 -helix – (n+5) → π-helix
 forms an h-bond with NH of residue: – (n+4) → α-helix – (n+3) → helix – (n+5) → π-helix")
35
35 α-helix Ribbon diagram of α-helix

36
36 α-helix Ramachandran of α-helix

37
Secondary (2 o ) structure Helices -helix 3 10 -helix -helix Sheets Antiparallel Parallel Turns and Unstructured In antiparallel -sheets Adjacent β-strands run in opposite directions Hydrogen bonds (dashed lines) between NH and CO stabilize the structure The side chains (in green) are above and below the sheet
 structure Helices -helix helix -helix Sheets Antiparallel Parallel Turns and Unstructured In antiparallel -sheets Adjacent β-strands run in opposite directions Hydrogen bonds (dashed lines) between NH and CO stabilize the structure The side chains (in green) are above and below the sheet")
38
Secondary (2 o ) structure Helices -helix 3 10 -helix -helix Sheets Antiparallel Parallel Turns and Unstructured In parallel -sheets Adjacent β-strands run in same direction Hydrogen bonds (dashed lines) between NH and CO stabilize the structure The side chains (in green) are above and below the sheet
 structure Helices -helix helix -helix Sheets Antiparallel Parallel Turns and Unstructured In parallel -sheets Adjacent β-strands run in same direction Hydrogen bonds (dashed lines) between NH and CO stabilize the structure The side chains (in green) are above and below the sheet")
39
39 Ribbon diagram of β sheet In addition to being purely parallel or antiparallel, β sheets can be mixed, with strands running in both parallel and antiparallel directions Arrow pointing to C-terminal end

40
40 Ramachandran of β-sheet

41
Secondary (2 o ) structure Helices -helix 3 10 -helix -helix Sheets Antiparallel Parallel Turns and Unstructured CO of residue i forms h-bonds with NH of either residue i+2, i+3, i+5, or i+4 β-turn i→i+3 H-bond (most common) γ-turn: i→i+2 H-bond α-turn i→i+4 H-bond π-turn i→i+5 H-bond
 structure Helices -helix helix -helix Sheets Antiparallel Parallel Turns and Unstructured CO of residue i forms h-bonds with NH of either residue i+2, i+3, i+5, or i+4 β-turn i→i+3 H-bond (most common) γ-turn: i→i+2 H-bond α-turn i→i+4 H-bond π-turn i→i+5 H-bond")
42
Secondary (2 o ) structure Helices -helix 3 10 -helix -helix Sheets Antiparallel Parallel Turns and Unstructured
 structure Helices -helix helix -helix Sheets Antiparallel Parallel Turns and Unstructured")
43
43 Proteins fold into compact tertiary structures with hydrophobic cores. (spacefill representation) Structural classification: http://www.cathdb.info/cgi-bin/cath/GotoCath.pl?link=class.html Tertiary (3 o ) structure
 Structural classification: link=class.html Tertiary (3 o ) structure.")
44
44 It is a bit difficult to see and understand anything here (sticks representation) Tertiary (3 o ) structure
 Tertiary (3 o ) structure")
45
45 To simplify representation, secondary structure diagrams are used Tertiary (3 o ) structure
 structure")
46
Quaternary (4 o ) structure Dimer of identical subunits ( Cro protein of bacteriophage lambda)
 structure Dimer of identical subunits ( Cro protein of bacteriophage lambda)")
47
47 Coat of rhinovirus Coat of rhinovirus (common cold) (common cold) 60 copies of 4 60 copies of 4 different subunits, different subunits, 3 outside, 3 outside, red, blue, green red, blue, green Quaternary structure
 (common cold) 60 copies of 4 60 copies of 4 different subunits, different subunits, 3 outside, 3 outside, red, blue, green red, blue, green Quaternary structure")
48
Protein structure databases Primary: UniProt http://www.uniprot.org/http://www.uniprot.org/ Secondary: DSSP http://rcsb.orghttp://rcsb.org Tertiary: PDB http://rcsb.orghttp://rcsb.org Quaternary: PQS http://www.ebi.ac.uk/pdbe/pqs/http://www.ebi.ac.uk/pdbe/pqs/

49
49 NOTE: Components Sugar Base Phosphate 5’ to 3’ direction T->U in RNA RNA - extra –OH at 2’ of pentose sugar DNA - deoxyribose Numbering Single vs double strands DNA more stable Voet, Donald and Judith G. Biochemistry. John Wiley & Sons, 1990, p. 792. DNA and RNA Structure

50
50 NOTE: Pyrimadines and Purines T->U in RNA Names Numbering Bonding character Position of hydrogen Tautomers The 5 Bases of DNA and RNA Purines Pyrimadines

51
51 Keto vs enol (OH) Different hydrogen bonding patterns Tautomeric Structures
 Different hydrogen bonding patterns Tautomeric Structures")
52
A:T and G:C pairs are spatially similar 3 H-bonds vs 2 (GC rich?) Sugar groups are attached asymmetrically on the same side of the pair Leads to a major and minor grove Bases are flat but the hydrogen bonding leads to considerable flexibility Base stacking is flexible Geometry of Watson Crick Base Pairs Voet, Donald and Judith G. Biochemistry. John Wiley & Sons, 1990, p. 797. 52Pharm201 Lecture 2 2010

53
53 Hydrogen bonding of WC base pair Mechanisms of recognition The canonical Watson-Crick base pair, shown as the G-C pair. Positions of the minor and major grooves are indicated. The glycosidic sugar-base bond is shown by the bold line; hydrogen bonding between the two bases is shown in dashed lines. Definition of Major and Minor Groove

54
Pharm201 Lecture 2 201054 Base Morphology The base-pair reference frame is constructed such that the x-axis points away from the (shaded) minor groove edge. Images illustrate positive values of the designated parameters. Reprinted with permission from Adenine Press from (Lu, et al., 1999).
..")
55
Pharm201 Lecture 2 201055 Backbone Conformation Voet, Donald and Judith G. Biochemistry. John Wiley & Sons, 1990, p. 807.

56
Pharm201 Lecture 2 201056 A Beta-nucleoside Ring is never flat – has 5 internal torsional angles The pucker is determined by what is bound A variety of puckers have been observed Pucker has a strong influence on the overall conformation

57
57 The Glycosidic Bond Connects ribose sugar to the base Anti Syn

58
58 Change in sugar conformation affects the backbone C2’-Endo C3’-Endo C3’ C2’

59
Pharm201 Lecture 2 201059 A DNA B DNA..and the position of the bases relative to the helix axis

60
Pharm201 Lecture 2 201060 Neidle, Stephen. Nucleic Acid Structure and Recognition. Oxford University Press, 2002, p. 34. Canonical B DNA

61
Pharm201 Lecture 2 201061 Canonical B DNA First determined experimentally by fiber diffraction (Arnott) C2’-endo sugar puckers High anti glycosidic angles Right handed – 10 base pairs per turn Bases perpendicular to the helix axis and stacked over the axis Overall bending as much as 15 degrees (result of base morphologies – twist and roll) – {machine learning – sequence vs overall conformation?} Over 230 structures 25 with base mis-pairing – only cause local perturbations Strong influence of hydration along spine http://ndbserver.rutgers.edu/index.html
 C2’-endo sugar puckers High anti glycosidic angles Right handed – 10 base pairs per turn Bases perpendicular to the helix axis and stacked over the axis Overall bending as much as 15 degrees (result of base morphologies – twist and roll) – {machine learning – sequence vs overall conformation } Over 230 structures 25 with base mis-pairing – only cause local perturbations Strong influence of hydration along spine")
62
Pharm201 Lecture 2 201062 Neidle, Stephen. Nucleic Acid Structure and Recognition. Oxford University Press, 2002, p. 36. A DNA

63
Pharm201 Lecture 2 201063 Canonical A DNA C3’-endo sugar puckers – brings consecutive phosphates closer together 5.9A rather than 7.0 Glycosidic angle from high anti to anti Base pairs twisted and nearly 5A from helix axis Helix rise 2.56A rather than 3.4A Helix wider and 11 base pairs per repeat Major groove now deep and narrow Minor grove wide and very shallow

64
Pharm201 Lecture 2 201064 Z-DNA

65
Pharm201 Lecture 2 201065 Z-DNA Helix has left-handed sense Can be formed in vivo, given proper sequence and superhelical tension, but function remains obscure. Narrower, more elongated helix than A or B. Major "groove" not really groove Narrow minor groove Conformation favored by high salt concentrations, some base substitutions, but requires alternating purine-pyrimidine sequence. N2-amino of G H-bonds to 5' PO: explains slow exchange of proton, need for G purine. Base pairs nearly perpendicular to helix axis GpC repeat, not single base-pair – P-P distances: vary for GpC and CpG – GpC stack: good base overlap – CpG: less overlap. Zigzag backbone due to C sugar conformation compensating for G glycosidic bond conformation Conformations: – G; syn, C2'-endo – C; anti, C3'-endo

66
Pharm201 Lecture 2 201066 Z-DNA Convex major groove Deep minor groove Alternate C then G Spine of hydration

67
Pharm201 Lecture 2 201067 Quadruplex DNA 1NP9 Jmol

68
Pharm201 Lecture 2 201068 Saenger, Wolfram. Principles of Nucleic Acid Structure. Springer-Verlag New York Inc., 1984, p. 333. tRNA 1EVV jmol Invariant L-shape

69
Pharm201 Lecture 2 201069 Neidle, Stephen. Nucleic Acid Structure and Recognition. Oxford University Press, 2002, p. 148. tRNA H bonds between distant regions

70
Pharm201 Lecture 2 201070 The Ribosome Complex of protein and RNA Small 30S subunit – controls interactions between mRNA and tRNA Large 50S subunit – peptide transfer and formation of the peptide bond

71
Nucleic acid structure databases Primary: UniProt http://www.uniprot.org/http://www.uniprot.org/ Tertiary: PDB http://rcsb.org contains all of http://ndbserver.rutgers.edu/http://rcsb.org http://ndbserver.rutgers.edu/

72
Molecular viewers Pymol Rasmol Jmol Molmol VMD Swiss PDB etc… (please note, only freeware are listed)
")
73
Transcription and translation (DNA RNA Protein)
")
74
Transcription and translation (DNA Protein)
")
Similar presentations








 Nucleic Acids. Information encoded in a DNA molecule is transcribed via synthesis of an RNA molecule The sequence of the RNA molecule.>")





 Dr. Sumbul Fatma>")




