Download presentation
Presentation is loading. Please wait.

1
Huck-Hui Ng Genome Institute of Singapore 17 July 2010 Stem cells and epigenetics

2
I. Stem cells II. Epigenetics

3
Ralston and Rossant, 2005; Chazaud et al., 2006; Ralston and Rossant, 2008 1-cell2-cell4-cell8-cell uncompacted 8-cell compacted 16-24 cell morula Cavitating morula E3.5 Blastocyst E4.5 Blastocyst TE Mouse pre-implantation development

4
http://stemcells.nih.gov/ Mouse pre-implantation development

5
Embryonic stem cells http://stemcells.nih.gov

6
Stem cells from blastocyst lineages Trophoblast stem (TS) cells are derived from the trophectoderm lineage Embryonic stem (ES) cells represent the epiblast lineage Extraembryonic endoderm (XEN) cells derive from the primitive endoderm lineage Ralston A, Rossant J. Genetic regulation of stem cell origins in the mouse embryo. Clin Genet 2005: 68: 106–112.

7
The stem-cell hierarchy Eckfeldt CE et al. Nat Rev Mol Cell Biol. (2005)
")
8
Waddington’s epigenetic landscape model

9
Embryonic stem cells 1)First isolated from mouse embryos in 1981. Evans and Kaufman. Nature. (1981) 292:154-6. Martin. Proc Natl Acad Sci U S A. (1981) 78:7634-8. 2)Gene targeting / Generation of transgenic mice (Animal model for the study of gene functions in vivo) 3)Human ES cells Thomson et al. Science. (1998) 282:1145-7 Reubinoff et al. Nat Biotechnol. (2000) 18:399-404.
 292: Martin. Proc Natl Acad Sci U S A. (1981) 78: )Gene targeting / Generation of transgenic mice (Animal model for the study of gene functions in vivo) 3)Human ES cells Thomson et al. Science. (1998) 282: Reubinoff et al. Nat Biotechnol. (2000) 18:")
10
http://images.crinet.com How does the cell read genetic information? Sequence-specific transcription factor recruits multi-subunit complexes that can modulate transcription and chromatin structure. Taatjes et al. (2004). Nat Rev Mol Cell Biol. 5(5):403-10
. Nat Rev Mol Cell Biol. 5(5):")
11
Oct4 A POU transcription factor expressed by early embryo cells and germ cells (Schöler et al, 1990. Nature). Required for the formation of pluripotent stem cells in the mammalian embryo (Nichols et al, 1998. Cell). Required for the maintenance of pluripotency of ES cells and controls lineage commitment (trophectoderm) (Niwa et al, 2000. Nat Genet).
. Required for the formation of pluripotent stem cells in the mammalian embryo (Nichols et al, Cell). Required for the maintenance of pluripotency of ES cells and controls lineage commitment (trophectoderm) (Niwa et al, Nat Genet)..")
12
Chromatin Immunoprecipitation (ChIP) Formaldehyde: relatively non-specific high resolution crosslinker (2 Å) covalent crosslink is reversible (by heating in the presence of Tris) fixation is extremely rapid Cells are frozen in native state “snap shot” HCHO crosslinking in living cells Sonication Immunoprecipitation to enrich for binding sites
 Formaldehyde: relatively non-specific high resolution crosslinker (2 Å) covalent crosslink is reversible (by heating in the presence of Tris) fixation is extremely rapid Cells are frozen in native state snap shot HCHO crosslinking in living cells Sonication Immunoprecipitation to enrich for binding sites")
13
advantages The advantages of Chromatin IP allows one to probe the direct physical relationship between DNA binding proteins and their DNA targets in vivo measurement of physical occupancy (crosslinking in living cells) physiologically relevant targets (wild type context)
 physiologically relevant targets (wild type context)")
14
Mapping transcription factor binding sites Aims: 1) How is the ES cell genome wired? 2) Are there cross-talks between the key signaling pathways and the other transcription factors? 3) Can we infer the composition of multi-protein complexes assembled on the chromatin?
 Are there cross-talks between the key signaling pathways and the other transcription factors. 3) Can we infer the composition of multi-protein complexes assembled on the chromatin .")
15
Where do transcription factors bind in the genomic space? * * * * * * * * core factors * signaling effectors * self-renewal regulator

16
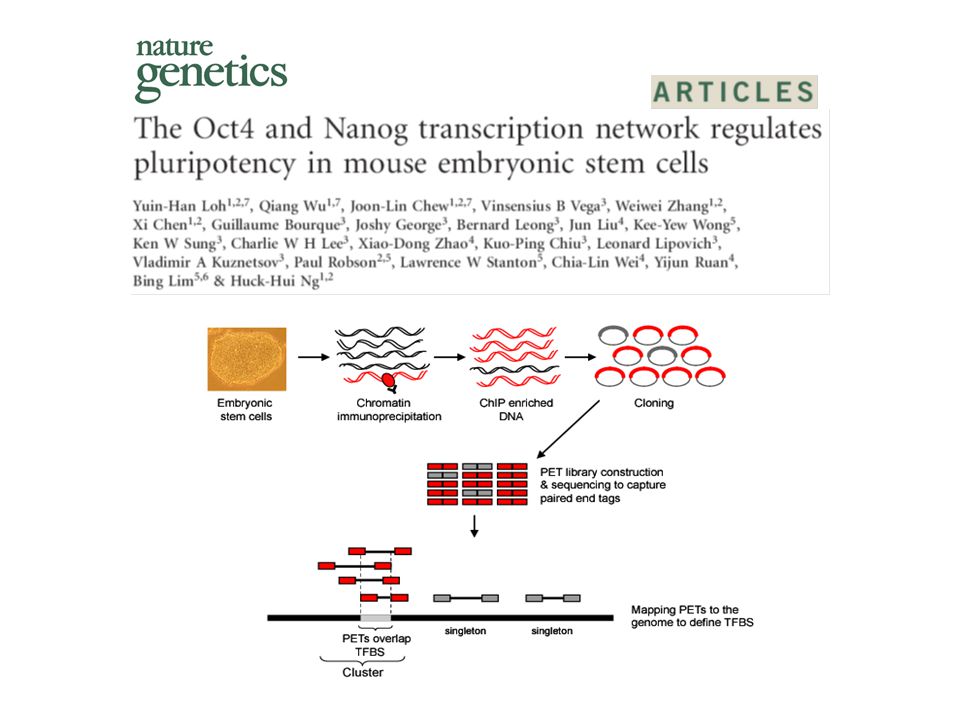
High resolution localization of sites using ChIP-seq method Oct4 gene (4.7 kb) Oct4 binding
 Oct4 binding")
17
Binding profiles of 13 sequence specific transcription factors at Oct4 and Nanog loci

18
Colocalization of transcription factor binding sites at Oct4 and Nanog enhancers

19
Transcription factor relationship at multiple transcription factor binding loci (MTL) Oct4-centric MTL Myc-centric MTL
 Oct4-centric MTL Myc-centric MTL")
20
Oct4-centric MTL can enhance transcription

21
ES cell-specific enhanceosomes: 1)Regions densely bound by multiple transcription factors (include Oct4, Sox2, Nanog, Smad1, STAT3 and others) 2)These sites are not commonly found at proximal promoter regions (-500bp, +2,000bp) 3)Function as enhancers 4)Bound by co-activators (p300, CBP, NcoA3)
Regions densely bound by multiple transcription factors (include Oct4, Sox2, Nanog, Smad1, STAT3 and others) 2)These sites are not commonly found at proximal promoter regions (-500bp, +2,000bp) 3)Function as enhancers 4)Bound by co-activators (p300, CBP, NcoA3)")
22
Clustering based on transcription factor binding sites reveals five classes of genes Nanog, Pou5f1, Sox2, Esrrb, Klf4, c-Myc, n-Myc, Rif1, Sall4, Tbx3, Tcf3, Tcfcp2l1, Zic3 Poorly expressed / silenced Genes expressed in ES cells ESC-specific expression constitutive expression Suz12 bound genes Myc bound genes

23
Summary Design principles of ES cell TF network 1)Regulatory loops for key transcription factors Loh et al (2006). Nat Genet; Jiang et al (2008). Nat Cell Biol 2)Highly connected network Hotspots for transcription factor co-binding - ES cell-specific enhanceosomes Nexus that integrate extracellular signaling and intrinsic pathways Chen et al (2008). Cell 3)Downstream targets of key TFs are important for ES cells Rif1, Esrrb, Klf2, Klf4, Klf5 Loh et al (2006). Nat Genet; Jiang et al (2008). Nat Cell Biol
. Nat Cell Biol 2)Highly connected network Hotspots for transcription factor co-binding - ES cell-specific enhanceosomes Nexus that integrate extracellular signaling and intrinsic pathways Chen et al (2008). Cell 3)Downstream targets of key TFs are important for ES cells Rif1, Esrrb, Klf2, Klf4, Klf5 Loh et al (2006). Nat Genet; Jiang et al (2008). Nat Cell Biol.")
24
How can you de-differentiate a somatic cell? Yamanaka, 2007. Cell Stem Cell. 1(1):39-49 Takahashi and Yamanaka, 2006. Cell
:39-49 Takahashi and Yamanaka, Cell.")
25
Oct4, Sox2, Klf4, c-Myc Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006 Aug 25;126(4):663-76. Epub 2006 Aug 10. Transcription factors can specify ES cell identity in non-stem cells
: Epub 2006 Aug 10. Transcription factors can specify ES cell identity in non-stem cells.")
26
Are different somatic cells amenable to reprogramming? Hochedlinger and Plath. Development 136, 509-523 (2009)
.")
27
The path towards induced pluripotency Hochedlinger and Plath. Development 136, 509-523 (2009)
")
28
II. Epigenetics

29
What is epigenetics?

30
References: Li, E. (2002). Nature Reviews Genetics. Chromatin modification and epigenetic reprogramming in mammalian development. Bird, A. (2002). Genes & Development. DNA methylation patterns and epigenetic memory. Bird, A. (2007). Nature. Perceptions of epigenetics.
. Genes & Development. DNA methylation patterns and epigenetic memory. Bird, A. (2007). Nature. Perceptions of epigenetics..")
31
Epigenetics: 'outside conventional genetics' The study of mitotically and / or meiotically heritable changes in gene function that cannot be explained by changes in DNA sequence. Russo, V.E.A., Martienssen, R.A., and Riggs, A.D. 1996. Epigenetic mechanisms of gene regulation. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. Bird. 2002. DNA methylation patterns and epigenetic memory. Genes Dev. 16: 6- 21. There are two epigenetic systems that affect animal development and fulfill the criterion of heritability: DNA methylation and the Polycomb-trithorax group (Pc- G/trx) protein complexes. (Histone modification has some attributes of an epigenetic process, but the issue of heritability has yet to be resolved.)
 protein complexes. (Histone modification has some attributes of an epigenetic process, but the issue of heritability has yet to be resolved.).")
32
yeast chromosome telomere ADE2 (wildtype) gene at normal location on chromosome centromere wildtype white colony telomere ADE2 (wildtype) gene moved to location near telomere centromere red sectored colony Yeast position effect variegation: sectored colonies
 gene at normal location on chromosome centromere wildtype white colony telomere ADE2 (wildtype) gene moved to location near telomere centromere red sectored colony Yeast position effect variegation: sectored colonies")
33
Yeast position effect variegation Metastable states (on and off) Single cell Grown up colony Epigenetic properties: 1)Mitotically heritable 2)Cannot be explained by changes in DNA sequence
 Single cell Grown up colony Epigenetic properties: 1)Mitotically heritable 2)Cannot be explained by changes in DNA sequence")
34
Unifying definition of epigenetic events: The structural adaptation of chromosomal regions so as to register, signal or perpetuate altered activity states. Bird (2007). Nature. 447, 396-398.
. Nature. 447,")
35
Nature 2003 Jan 23;421(6921):448-53 Controlling the double helix. Felsenfeld G, Groudine M. Expanded meaning of “epigenetics”

36
Special features of nuclear architecture in embryonic stem cells

37
Nuclear architecture in ES cells and differentiating ES-derived cells Meshorer E, Misteli T. Nat Rev Mol Cell Biol. 2006 Jul;7(7):540-6
:")
38
Nature 2003 Jan 23;421(6921):448-53 Controlling the double helix. Felsenfeld G, Groudine M. Histone H3 Lys 9 methylation (repressive mark) Histone H3 acetylation (active mark) Histone H4 acetylation (active mark) Histone H3 Lys 4 methylation (active mark)
 Histone H3 acetylation (active mark) Histone H4 acetylation (active mark) Histone H3 Lys 4 methylation (active mark).")
39
Breathing Chromatin of ES cells

40
Meshorer E, Misteli T. Nat Rev Mol Cell Biol. 2006 Jul;7(7):540-6 Open chromatin architecture permissive for gene expression and pluripotency in ES cells ? UndifferentiatedDifferentiated
:540-6 Open chromatin architecture permissive for gene expression and pluripotency in ES cells . UndifferentiatedDifferentiated.")
41
Working model: The crosstalk between the transcription factor network and the epigenetic mechanism in the maintenance of pluripotency Oct4 chromatin structure in ES cells ? Genetic / biochemical interactions with chromatin modifiers?

42
1. The roles of histone H3K9 demethylases in ES cells 2. Oct4 and histone H3K9 methylase The roles of histone modifiers in ES cells

44
Oct4’s target genes Mapping of transcription factor binding sites in living cells and study how the targets relate to ES cell properties

45
Oct4 ChIP-PET clusters mapped to Jmjd1a and Jmjd2c Jmjd1a Jmjd2c

46
Histone methylation is reversible H3K36me2 demethylase

47
Zhang and Reinberg (2001) Genes Dev. Different states of lysine methylation Me1Me2Me3 histone H3 lysine methylases histone H3 lysine demethylases

48
Cell. 2006 May 5;125(3):483-95. Cell. 2006 May 5;125(3):467-81. Nature. 2006 Jul 20;442(7100):312-6. K9 Jmjd1a Demethylation of H3K9 me2 K9 Jmjd2c Demethylation of H3K9 me3 K9 Jmjd2c K9 Jmjd1a Histone H3 lys 9 demethylases
: K9 Jmjd1a Demethylation of H3K9 me2 K9 Jmjd2c Demethylation of H3K9 me3 K9 Jmjd2c K9 Jmjd1a Histone H3 lys 9 demethylases.")
49
Control +RA Jmjd1a -LIF H3K9Me2 H3 Differentiation of ES cells leads to reduction of Jmjd1a and increase in H3K9Me2 β-tubulin Western

50
Control +RA Jmjd2c -LIF β-tubulin H3K9Me3 H3 Differentiation of ES cells leads to reduction of Jmjd2c and increase in H3K9Me3 Western

51
Jmjd1a and Jmjd2c are induced in reprogrammed fibroblasts Takahashi K, Yamanaka S. Cell. 2006 Aug 25;126(4):663-76
:")
52
Oct4 directly regulates Jmjd1a and Jmjd2c A B

53
Oct4 binds to intronic sequences of Jmjd1a and Jmjd2c

54
Depletion of Jmjd1a induces H3K9Me2 but not H3K9Me3 K9 Jmjd1a K9 Western

55
Depletion of Jmjd2c induces H3K9Me3 but not H3K9Me2 Western K9 Jmjd2c

56
Depletion of Jmjd1a and Jmjd2c induces ES cell differentiation

57
Changes in expression of ES cell and differentiation markers upon Jmjd1a or Jmjd2c depletion

58
histone modifiers Global effectsLocalized effects Actions of JmjC histone demethylases and their roles in ES cells Large scale regulation e.g. heterochromatization Regulation of specific promoters e.g. targeted recruitment gene coating transient interactions TF

59
Tcl1 is regulated by Jmjd1a Tcl1: T-cell lymphoma breakpoint 1 enhances Akt kinase activity and induces its nuclear translocation a self-renewal regulator in ES cells (Ivanova (2006). Nature; Matoba (2006). PLoS ONE)
. PLoS ONE).")
60
Jmjd1a regulates the expression and H3K9Me2 of Tcl1 AB C D

61
Tcl1 promoter Oct4 Oct4 binding at Tcl1 promoter is dependent on Jmjd1a

62
Jmjd2c regulates the expression and H3K9Me3 of Nanog

63
Jmjd2c binds to Nanog promoter ABC

64
Increased H3K9Me3 at Nanog promoter leads to increased binding of co-repressors (HP1 and KAP1) ABC
 ABC")
65
Oct4 Tcl1Nanog Tcl1Nanog Jmjd1aJmjd2c Maintenance of ES cells guardian of self-renewal genes Interface between genetic and epigenetic regulation

66
Model: role of histone demethylases in maintaining ES cells Open chromatin Low H3K9 methylation Self-renewal Oct4 Sox2 Nanog Condensed chromatin High H3K9 methylation Jmjd1a Jmjd2c Jmjd1a Jmjd2c Differentiation

67
Summary 1.Oct4 directly regulates Jmjd1a and Jmjd2c, which are upregulated in ES cells. 2.Depletion of Jmjd1a and Jmjd2c leads to global increase in H3K9Me2 and H3K9Me3, respectively. 3.Depletion of Jmjd1a leads to ES cell differentiation. 4.Novel regulatory pathways used by Oct4 to maintain the expression of its downstream targets (Tcl1 and Nanog).
..")
68
Histone H3 lys 9 methylase Mol Cell. 2003 Aug;12(2):475-87. Methylation of H3K9 me3 K9 Jmjd2c K9 Jmjd2c Eset Knockout of Eset Mol Cell Biol. 2004 Mar;24(6):2478-86.
:")
69
Depletion of Eset induces differentiation

70
Tcfap2a, Tcfap2c and Cdx2 are repressed by Eset H3K9me3 H3K9me2 Eset occupancy

71
Eset depleted cells express Cdx2 and Cdh3

72
Eset interacts with Oct4 and recruitment of Eset is dependent on Oct4

73
Model for regulation of cell fate by Oct4 through Eset

74
Summary 1.Eset interacts with Oct4. 2.Depletion of Eset leads to ES cell differentiation. 3.Eset regulates H3K9Me2 and H3K9Me3 of Tcfap2a and Tcfap2c. 4.Oct4 may control pluripotency through Eset (selective recruitment of Eset by Oct4 to lineage specific genes).
..")
75
Overview 1.Oct4 directly controls the expression of histone H3 lysine 9 demethylases (Jmjd1a and Jmjd2c). 2.Oct4 recruits Eset (histone H3 lysine 9 methylase) to repress trophectoderm lineage and genes. Oct4 Jmjd1aJmjd2c Eset Suppress H3K9 methylations and promote expression of self-renewal genes Mediates H3K9 methylations and repress trophectoderm lineage
 to repress trophectoderm lineage and genes. Oct4 Jmjd1aJmjd2c Eset Suppress H3K9 methylations and promote expression of self-renewal genes Mediates H3K9 methylations and repress trophectoderm lineage.")
76
Cellular potency in development and reprogramming Hemberger et al (2009). Nat. Rev. MCB
. Nat. Rev. MCB")
Similar presentations