Download presentation
Presentation is loading. Please wait.

1
Molecular mechanism of plant flowering time control
第十章 基因和发育 第二讲 植物开花时间控制的分子机理 Molecular mechanism of plant flowering time control By Hongwei Guo, Peking University,

2
When to flower is a matter of the survival of species
Transition to reproduction When to flower is a matter of the survival of species ? Main inflorescence shoot in middle, that can branch into lateral shoot At end of each shoot – place to make flower Each inflorescence shoot can make leaves and lateral organs What regulates transition from vegetative growth (main function to increase biomass of plant) to making plant sexually ready Vegetative phase Reproductive phase
 to making plant sexually ready. Vegetative phase. Reproductive phase.")
3
Two major pathways regulating floral transition
Photoperiodic flowering (光周期开花): day-length dependent flowering time control Vernalization (春化): cold-promoted flowering
: day-length dependent flowering time control. Vernalization (春化): cold-promoted flowering.")
4
(“Maryland Mammoth” cultivar of tobacco)
")
8
Floral induction of leaves of Short Day plant Perrila crispa
“Something” must be produced in leaves and “move” to the meristem Long journey to identify “Florigen” began (Lang and Zeevart)
")
9
Multiple grafting experiment with Perilla
Only single leaf from one of the plant treated – all 5 plants can make flower Something made in leaf can be transferred Flowering signal is grafting transmittable Long distance transport---- thru vascular tissue

10
The flowering signal: florigen
vegetative or reproductive growth? the flowering signal is generated in the leaf the signal goes one way: from the leaf to the apex Grafting transmittable SAM Florigen Florigen can be transmitted to other plants thru grafting Flower locus T protein claimed to be florigen – but some disputes If you find signal or molecule that can greatly change flowering time – agriculture revolutionized Florigen Florigen

11
Photoperiodic flowering
Two essential questions: 1. How does the leaf measure day-length? 2. What is the florigen?

12
Genetics provides the answers
Arabidopsis: Long Day Plant Flowering is induced by Long Day (LD) Certain late flowering mutants are blind to photoperiod cryptochrome 2 (cry2) phytochrome A (phyA) contstans (co) flowering locus t (ft) Cry2: blue light receptor PhyA: Far-red light receptor CO: transcriptional co-activator FT: transcriptional co-activator (?)
 Certain late flowering mutants are blind to photoperiod. cryptochrome 2 (cry2) phytochrome A (phyA) contstans (co) flowering locus t (ft) Cry2: blue light receptor. PhyA: Far-red light receptor. CO: transcriptional co-activator. FT: transcriptional co-activator ( )")
14
3 wavelengths (Blue, Red, Far-red) are important in regulating plant growth and development
 are important in regulating plant growth and development")
15
phyA also absorbs blue light
Phytochromes (光敏色素):Red/Far-red light receptors Cryptochromes (隐花色素):Blue light receptors phyA also absorbs blue light
:Red/Far-red light receptors. Cryptochromes (隐花色素):Blue light receptors. phyA also absorbs blue light.")
16
Signal Trans-duction

17
(originally proposed by Boenning, 1936)
Light has a dual role in this model: - entrains the circadian oscillation of light- and dark-sensitive phases - directly required for the production of the signal.

18
PRR: photoperiod response regulator

19
PRR: photoperiod response regulator

22
CO is essential for photoperiodic flowering, as co mutant is late flowering and almost a day-neutral plant. It encodes a transcriptional regulator.

24
Is co the PRR that measures daylength?
LFY A, B, C, E class genes (春化) Is co the PRR that measures daylength?
 Is co the PRR that measures daylength")
26
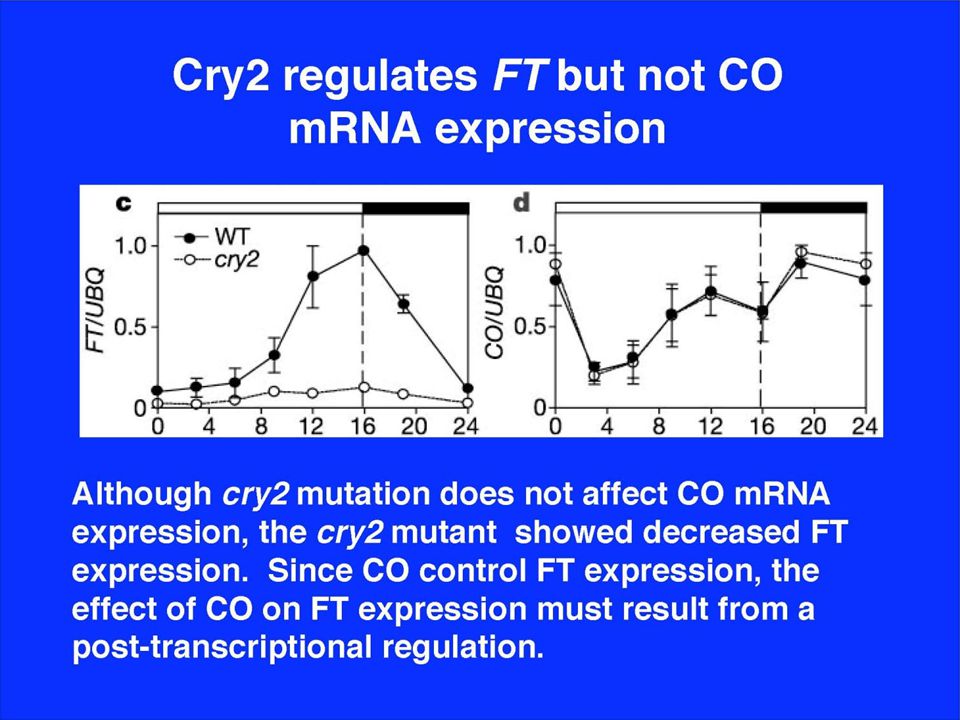
CO is required for FT expression

27
FT mRNA levels determine flowering time
LD: early flowering SD: late flowering phyBDE: early flowering cry2: late flowering

33
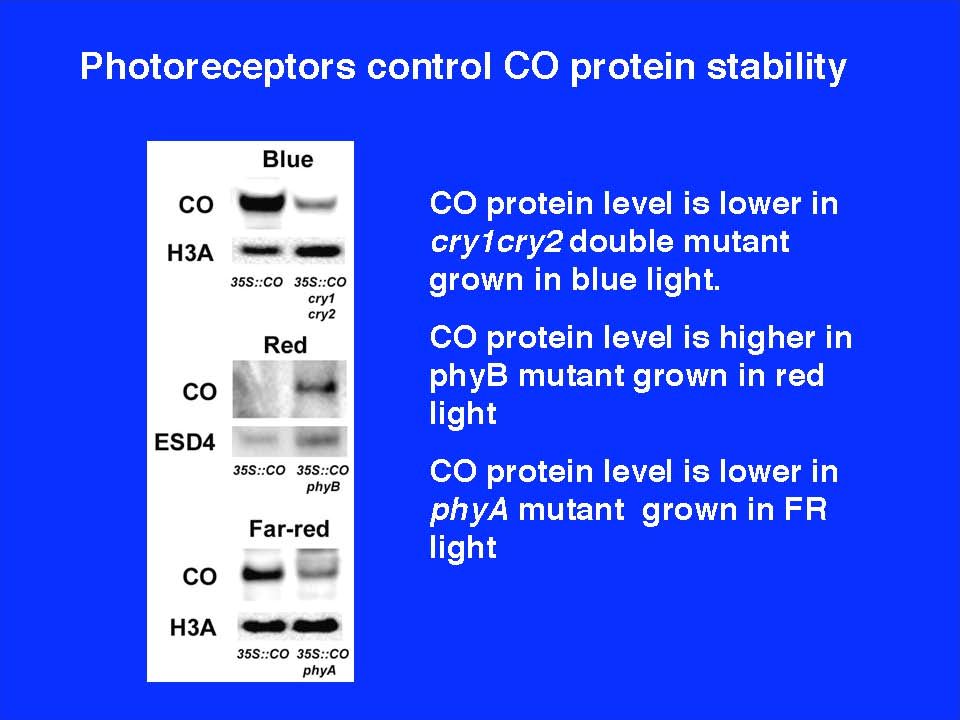
Cry inhibits CO protein degradation in light
When CO mRNA peaks at midnight in SDs, COP1 is predominantly localized in the nucleus. CRY interacts with COP1 but is not able to repress its activity. COP1–CO interaction results in ubiquitination and degradation of CO (A), whereas when CO mRNA peaks in the afternoon in LDs, light activation of CRY during the daytime might mediate translocation of COP1 from nucleus to cytoplasm. Consequently, CO is able to accumulate and activate the transcription of FT to promote flowering (B). U, ubiquitin.
, whereas when CO mRNA peaks in the afternoon in LDs, light activation of CRY during the daytime might mediate translocation of COP1 from nucleus to cytoplasm. Consequently, CO is able to accumulate and activate the transcription of FT to promote flowering (B). U, ubiquitin.")
34
Molecular mechanism of photoperiodic flowering

35
Photoperiod-dependent activation of CO protein and FT mRNA
CDF1: circadian dependent factor CO protein measures day-length FT mRNA level determines flowering time CO mRNA is regulated by circadian clock. CO protein is stabilized by light Therefore, FT expression is activated only in long day

36
FT (or Hd3a) is a floral activator both in LD and SD plants
Long Day Plant Short Day Plant (Kobayashi & Weigel, 2007) CO in Long Day plants and similar proteins (Hd1) in Short Day plants are regulated in opposite ways
 CO in Long Day plants and similar proteins (Hd1) in Short Day plants are regulated in opposite ways.")
38
2005 retr

39
Heating the leaf of pHSP::FT transgenic plants can promote flowering, and FT mRNA can be detected in SAM

41
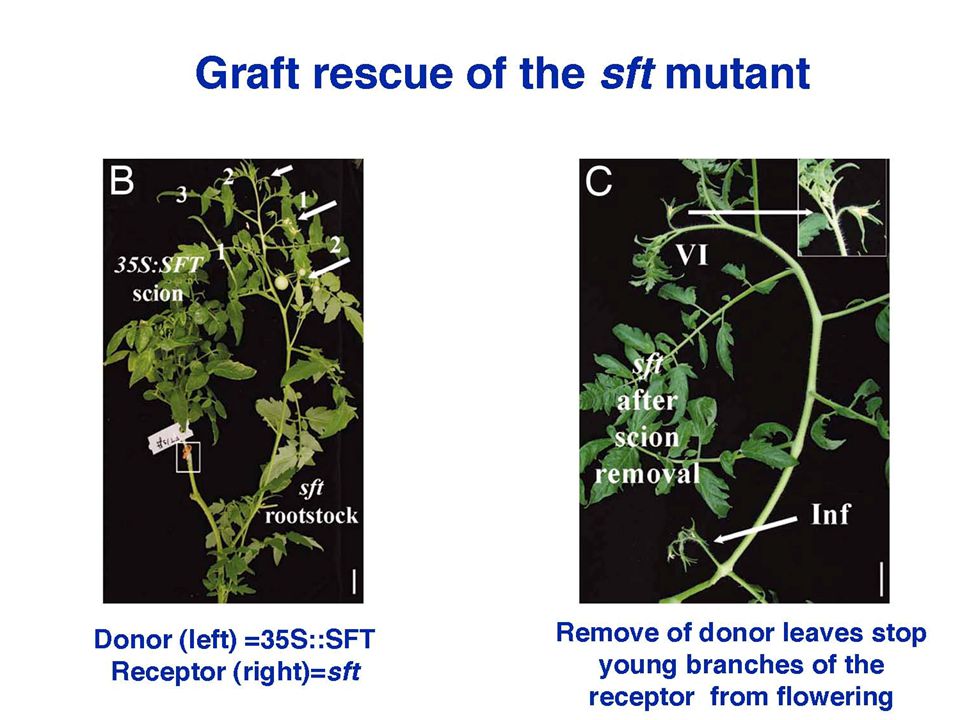
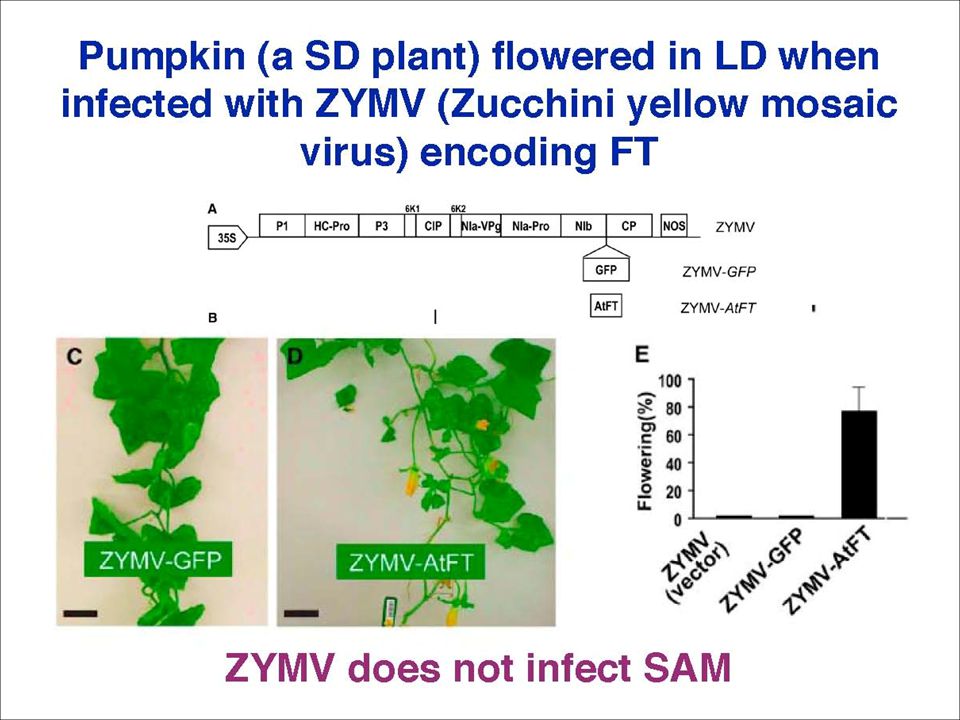
However, the mRNA hypothesis was challenged in 2006 by a PNAS paper, the original Science paper was retracted in more papers have been published in 2007 – all argued that FT protein is the florigen, in at least 5 different plant species: Arabidopsis, rice, tomato, tobacco, pumpkin. Yuval Eshed‘s lab in Israel cloned tomato FT gene from a tomato mutant, sft. They overexpressed SFT in tomato, resulting in early flowering, but they found little transgenic SFT mRNA in the apical meristem。

44
SAM leaves

58
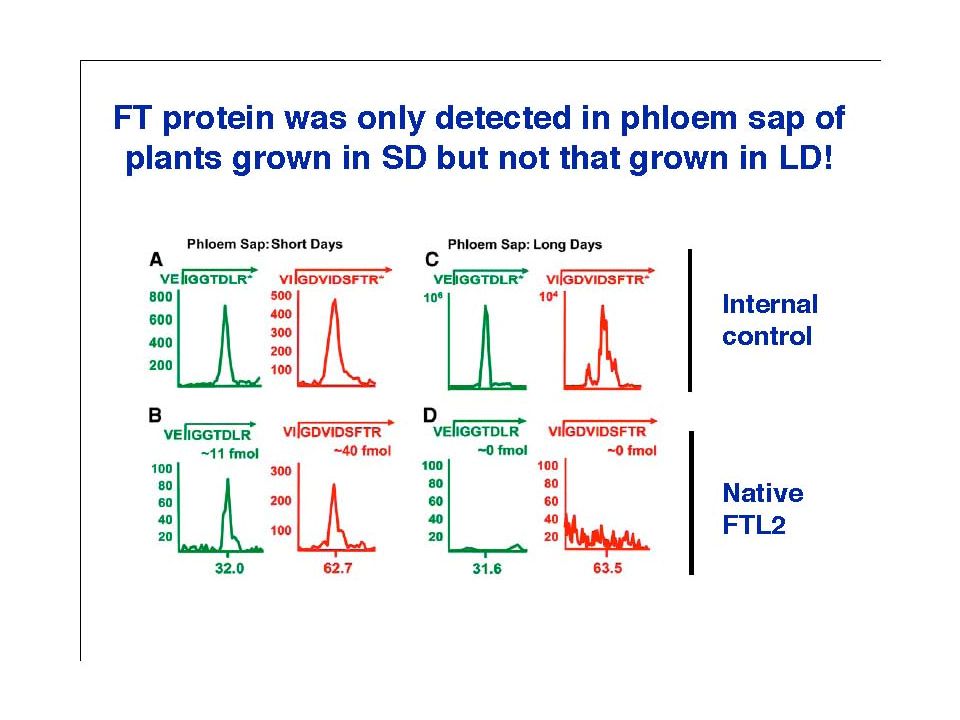
Where in the vascular tissue is FT mRNA or
FT protein locate (and migrate)? Phloem sap: solutes migrating thru phloem (sieve tube)
 Phloem sap: solutes migrating thru phloem (sieve tube)")
64
So…….. FT protein is a “florigen”
CO – transcription factor in leaves, respond to day-length (photoperiod sensor) FT – RAF kinase inhibitor protein, travels in phloem from leaf to SAM, target of CO
 FT – RAF kinase inhibitor protein, travels in phloem from leaf to SAM, target of CO.")
65
Photoperiodic flowering in plants
FT protein GI FT protein Hd3a protein

66
Besides the photoperiod-dependent regulation, floral transition is under controls of many other cues.

67
Vernalization (春化)– Promoting flowering with cold
– Promoting flowering with cold")
68
Some plants need winter to flower
Vernalization: Acquisition of the competence to flower in the spring by exposure to the prolonged cold of winter. Plants are genetically identical No vernalization Vernalization Exposed as a seedling to 4ºC for 40 days.

69
A role of temperature in the plant calendar
Changes in day length are a reliable indicator of seasonal progression, but day length per se is not completely informative of the time of year. Vernalization – the process whereby flowering is promoted by a cold treatment given to a fully hydrated seed (i.e., a seed that has imbibed water) or to a growing plant. - dry seeds do not respond to cold treatment - without cold treatment, plants that require vernalization show delayed flowering or remain vegetative
 or to a growing plant. - dry seeds do not respond to cold treatment. - without cold treatment, plants that require vernalization show delayed flowering or remain vegetative.")
70
Vernalization may involve epigenetic changes in gene expression
Requirements and features of vernalization: - requires cell division and DNA replication - requires stable changes in gene expression in meristem (even after the signal that induced the change, i.e. cold, is removed → epigenetic regulation) Epigenetic: A heritable change in gene expression that is controlled by modifications in DNA methylation and/or chromatin structure. - from yeast to mammals Arabidopsis: gene acting as repressor of flowering: FLOWERING LOCUS C (FLC) FLC - encodes MADS box transcription factor, delaying floral transition - represses transcription of AGAMOUS-LIKE 20 (AGL20)/SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1), encoding MADS box transcription factor accelerating flowering - highly expressed in non-vernalized SAM - after vernaliztion, gene is epigenetically switched off for remainder of plant’s life cycle, permitting flowering in response to long days to occur
 Epigenetic: A heritable change in gene expression that is controlled by modifications in DNA methylation and/or chromatin structure. - from yeast to mammals. Arabidopsis: gene acting as repressor of flowering: FLOWERING LOCUS C (FLC) FLC - encodes MADS box transcription factor, delaying floral transition. - represses transcription of AGAMOUS-LIKE 20 (AGL20)/SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1), encoding MADS box. transcription factor accelerating flowering. - highly expressed in non-vernalized SAM. - after vernaliztion, gene is epigenetically switched off for remainder of plant’s life cycle, permitting flowering in response to long days to occur.")
71
Vernalization blocks the expression of FLC in cold-requiring winter annual Arabidopsis ecotypes
winter annual ecotypes Winter annual without cold Winter annual after 40 days cold Winter annual without cold, but with flc mutation FLC mRNA

72
for vernalization insensitive (vin) mutants
Genetic screening for vernalization insensitive (vin) mutants VRN2: Homolog of Su(z)12 VRN1: Myb DNA binding protein VIN3: PHD finger protein LHP1: LIKE-HETEROCHROMATIN PROTEIN VIL1: PHD finger protein VIN5: Histone arginine methyl transferase VIN7: PAH2 domain protein Genes are involved in the regulation of chromatin structure Fewer;;;
 mutants. VRN2: Homolog of Su(z)12. VRN1: Myb DNA binding protein. VIN3: PHD finger protein. LHP1: LIKE-HETEROCHROMATIN PROTEIN. VIL1: PHD finger protein. VIN5: Histone arginine methyl transferase. VIN7: PAH2 domain protein. Genes are involved in the regulation of chromatin structure. Fewer;;;")
73
Other genes involved in control of flowering by vernalization
VERNALIZATION (VRN) 1 and VRN2 both are required for maintenance of low levels of FLC mRNA that are established by cold treatment once plants are exposed to warmer conditions Wild type Time vernalized Days at 20ºC after vernalization FLC mRNA Role of VRN2 is to maintain the repression of FLC expression. vrn2 Time vernalized Days at 20ºC after vernalization FLC mRNA
 1 and VRN2. both are required for maintenance of low levels of FLC mRNA that are established by cold treatment once plants are exposed to warmer conditions. Wild type. Time vernalized. Days at 20ºC after vernalization. FLC mRNA. Role of VRN2 is to maintain the repression of FLC expression. vrn2. Time vernalized. Days at 20ºC after vernalization. FLC mRNA.")
74
VRN2 encodes a gene related to Drosophila Polycomb-group (PcG) genes
Arabidopsis SU(Z)12 Drosophila In Drosophila, PcG proteins act in large protein complexes. They maintain the repression of transcription of homeotic genes, once the pattern of expression of these genes has been established during early embryo development.
12. Drosophila. In Drosophila, PcG proteins act in large protein complexes. They maintain the repression of transcription of homeotic genes, once the pattern of expression of these genes has been established during early embryo development.")
75
Polycomb-group complexes in Drosophila repress gene expression by modifying histones
Histone 3 is a major target for modifications – those above activate gene expression, those below repress it. Polycomb-group proteins promote the methylation of K9 and K27 (H3K9Me, H3K9Me) – gene repression VRN2 involved in chromatin remodeling → vernalization down-regulates FLC by epigenetic mechanisms
 – gene repression. VRN2 involved in chromatin remodeling → vernalization down-regulates FLC by epigenetic mechanisms.")
76
Repressed chromatin (OFF)
Histone code Active chromatin (ON) Repressed chromatin (OFF) High in: Acetylation, H3K4Me, H3S10P High in: H3K9Me, H3K27Me Modified histone could be recognized by activation/repression complexes and establish stable activation/repression chromatin
 Repressed chromatin (OFF) High in: Acetylation, H3K4Me, H3S10P. High in: H3K9Me, H3K27Me. Modified histone could be recognized by activation/repression complexes and establish stable activation/repression chromatin.")
77
Dynamics of FLC chromatin
Active FLC Chromatin High in Ac; H3K4Me; H3S10P H2A.Z Histone variant Repressed FLC Chromatin H3K9Me; H3K27Me H4R3Me2; LHP1 VIL1, VIN5 WINTER! VIN3, VIL1, VIN5 VRN2, VRN1, etc. ON; Fall OFF; Spring (from Sung and Amasino, 2005)
")
78
Memory of winter can be mitotically stable
From Lang & Melchers memory in Hyocyamus niger

79
FLC repression by vernalization is mitotically stable
VIN3 VIN3 UBQ We recapitulated fall/winter/spring seasons. Prior to vernalization as shown in red here represents fall condition, and blue indicated that plants are in the cold as in winter.

80
Molecular basis of vernalization response
FLC is a potent repressor of flowering. Competence: in Arabidopsis, is determined largely by FLC expression level. Vernalization leads to competence via repression of FLC. Mitotic stability: Vernalization-mediated repression of FLC via histone modifications that are hallmarks of epigenetic silencing

81
No vernalization FLC expression is subject to positive and negative regulation

83
Fall: flowering repressed
FLC is highly expressed and thus represses FT activation Short Days prevents CO-FT activation

84
FLC is repressed and FLC chromatin undergoes changes

85
Stable repression of FLC by chromatin changes eliminates antagonistic effect on FT activation
Long Days promote CO-FT activation

86
FLC resetting During meiosis, FLC is reactivated
(reversing chromatin structure)
")
87
Increase in AGL20 expression during floral induction in SAM
0 h h h d Times after shifting the plants from short days (8h L/16 h D) to long days (16h L/8h D)
 to long days (16h L/8h D)")
88
Multiple flowering control pathways
Many genes have variations even within same species provide flexibilities to adapt to local environments

89
The transition to flowering involves multiple factors and pathways

90
The transition to flowering involves multiple factors and pathways
FT FT

91
The Arabidopsis calendar

Similar presentations






















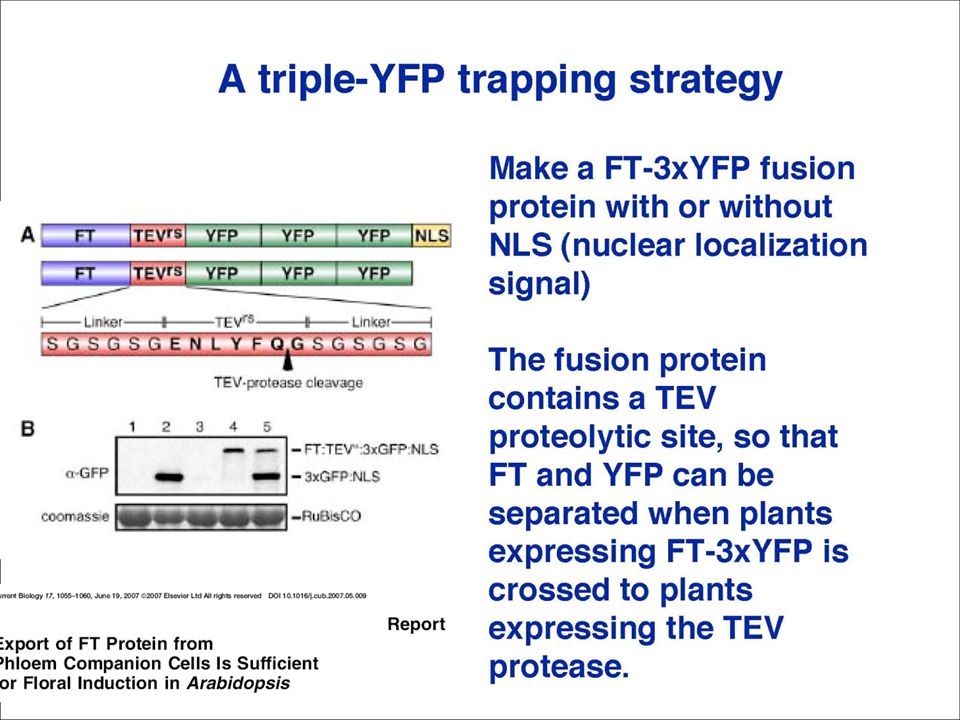










 DNA (gene) mRNA Protein Transcription RNA processing (splicing etc) Translation Folding Post translational modifications Peptides/amino.>")
 have two growth phases – vegetative growth, production of stems and leaves, occurs at the apical meristem,>")












 mRNA expression and CO protein stability. The circadian clock is a master regulator of photoperiod pathway components.>")
![How do plant cells grow? cell division (mitosis) cell expansion Growth rate [flux, water uptake = cell volume increase] depends on [driving force] / resistance.](/19/5757600/big_thumb.jpg "How do plant cells grow? cell division (mitosis) cell expansion Growth rate [flux, water uptake = cell volume increase] depends on [driving force] / resistance.>")
