Download presentation
Presentation is loading. Please wait.

1
AFM application: Beyond Imaging Probing Nano-Scale Forces Probing Nano-Scale Forces with the Atomic Force Microscope

2
How did the AFM system measure the force between tip and sample in contact mode?

4
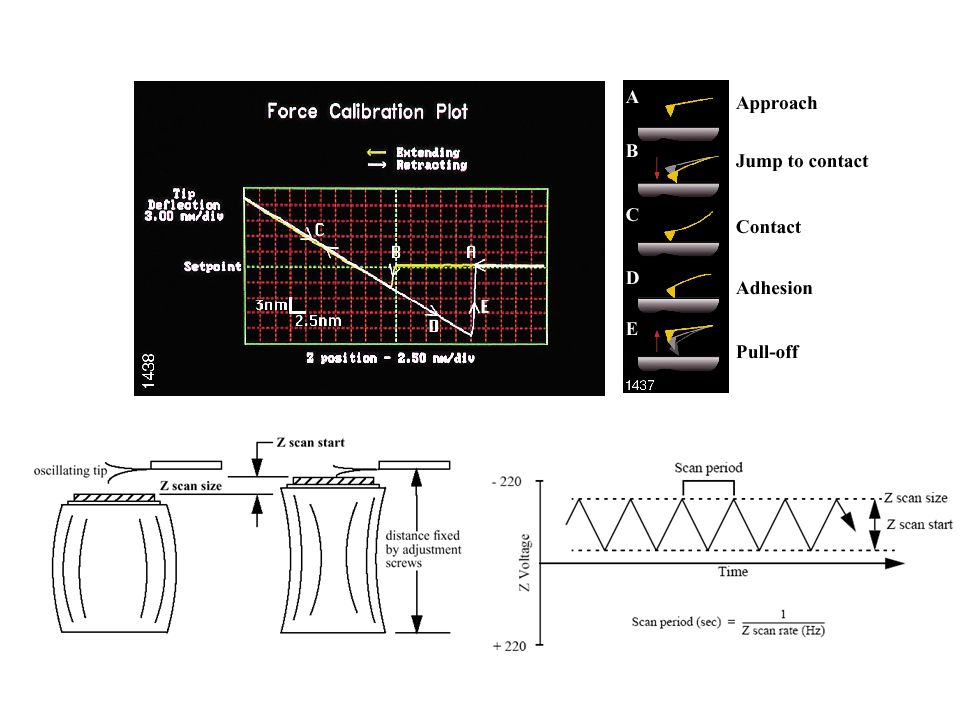
The cantilever starts (point A) not touching the surface. In this region, if the cantilever feels a long-range attractive (or repulsive) force it will deflect downwards (or upwards) before making contact with the surface. In the case shown, there is minimal long range force, so this “noncontact”part of the force curve shows no deflection. As the probe tip is brought very close to the surface, it may jump into contact (B) if it feels sufficient attractive force from the sample. Once the tip is in contact with the surface, cantilever deflection will increase (C) as the fixed end of the cantilever is brought closer to the sample. During the process, adhesion or bond forms between tip and sample. If the cantilever is sufficiently stiff, the probe tip may indent into the surface at this point. In this case, the slope or shape of the contact part of the force curve (C) can provide information about the elasticity of the sample surface. As the cantilever is withdrawn, adhesion or bonds formed during contact with the surface may cause the tip to adhere to the sample (section D) some distance past the initial contact point on the approach curve (B). A key measurement of the AFM force curve is the point (E) at which the adhesion is broken and the cantilever comes free from the surface. This can be used to measure the rupture force required to break the bond or adhesion.
 force it will deflect downwards (or upwards) before making contact with the surface. In the case shown, there is minimal long range force, so this noncontact part of the force curve shows no deflection. As the probe tip is brought very close to the surface, it may jump into contact (B) if it feels sufficient attractive force from the sample. Once the tip is in contact with the surface, cantilever deflection will increase (C) as the fixed end of the cantilever is brought closer to the sample. During the process, adhesion or bond forms between tip and sample. If the cantilever is sufficiently stiff, the probe tip may indent into the surface at this point. In this case, the slope or shape of the contact part of the force curve (C) can provide information about the elasticity of the sample surface. As the cantilever is withdrawn, adhesion or bonds formed during contact with the surface may cause the tip to adhere to the sample (section D) some distance past the initial contact point on the approach curve (B). A key measurement of the AFM force curve is the point (E) at which the adhesion is broken and the cantilever comes free from the surface. This can be used to measure the rupture force required to break the bond or adhesion..")
5
Details of force measurements

6
Photodiode sensitivity measurement ( convert Volts To cantilever deflection(nm) Hard surface Calculating forces F = - k x k: Spring constant of the cantilever; x: deflection of the cantilever Scanner Z position Volts Deflection
 Hard surface Calculating forces F = - k x k: Spring constant of the cantilever; x: deflection of the cantilever Scanner Z position Volts Deflection")
7
Spring constant measurement

8
Cleveland method: Rev. Sci. Instrum. 1993, 64, 403 This method is based upon measuring the resonance frequency of the cantilever in air, as a function of applied mass. When an end mass M is added, the resonant frequency is given by

9
http://www.asylumresearch.com/Applications/CantileverCal/ AddedMass.pdf Nondestructive Added Mass Spring Calibration with the MFP-3D™

12
Some typical force curves

13
Beginnings — Controlling Surface Adhesion Measurement of the forces between an AFM tip and a mica surface in air and water. The measurement in air shows a large adhesive force due to capillary forces from the liquid contaminant layer on the surface. Once the whole cantilever probe is immersed in water, capillary forces are mostly eliminated and adhesion is substantially diminished. From A.L. Weisenhorn, Appl. Phys. Lett. 1989, 54, 2651

14
Sharpened tip Imaging in vacuum Imaging in some solvent to reduce the tip-sample adhesion ( for example, alcohol, remove organic contamination) Ozone or UV treat the tip Tip modification Improve image quality
 Ozone or UV treat the tip Tip modification Improve image quality")
15
The AFM can measure forces on colloidal particles. In this case, a silica bead was glued to an AFM cantilever. Colloidal Forces The forces between colloidal particles dominate the behavior of a great variety of materials, including paints, paper, soils, clays and, in some circumstances, biological cells.” Colloid forces also play a critical role in the formation of ceramics and composite materials. Even in Pharmacy, Lotion, cream ( TiO2 and PbO)
.")
16
Science 1994, 264, 415

17
AFM Imaging Mode Revolution Contact ModeTappingModeForce-VolumePeakForce Tapping First introduced 1986 (original mode)1992~19942009 Tip-sample interaction Tip scans in constant contact with the sample Cantilever is oscillated at its resonance, so the tip intermittently contacts or “taps” the sample Whole probe is ramped linearly up and down, so the tip intermittently contacts the sample Whole probe is ramped sinusoidally, so the tip intermittently contacts or “taps” the sample Tip oscillation Not applicable At cantilever resonance (typ. 10-1000’s kHz) with typ. amplitude of 1-10’s nm Far below cantilever resonance (typ. <100 Hz) with typical amplitude of 100- 1000’s nm Below cantilever resonance (typ. <10 kHz) with typical amplitude of 10-100’s nm Imaging feedback Constant force (cantilever deflection) Constant tapping amplitude No feedback. Each curve individually triggered. Constant peak force Sketch Off surface On surface F0F0 F SP A free A SP F0F0 F0F0 F0F0 F peak F0F0 F0F0 F0F0 F trig Signature Feature from Bruker AFM Peak Force Tapping 5/8/201517 Bruker Confidential
 with typ. amplitude of 1-10’s nm Far below cantilever resonance (typ. <100 Hz) with typical amplitude of ’s nm Below cantilever resonance (typ. <10 kHz) with typical amplitude of ’s nm Imaging feedback Constant force (cantilever deflection) Constant tapping amplitude No feedback. Each curve individually triggered. Constant peak force Sketch Off surface On surface F0F0 F SP A free A SP F0F0 F0F0 F0F0 F peak F0F0 F0F0 F0F0 F trig Signature Feature from Bruker AFM Peak Force Tapping 5/8/ Bruker Confidential.")
18
5/8/201518Bruker Confidential Most Direct and Fastest way to conduct Quantitative Nanomechanical Mapping through force curves, no sample dependent empirical parameters required. Simultaneously obtain quantitative modulus, adhesion, dissipation, and deformation data while imaging topography at high resolution Direct force control keeps the imaging force low, which limits indentation depths to deliver non- destructive, high-resolution imaging Material properties can be characterized over a very wide range to address samples in many different research applications. Peak repulsive force can be as small as tens of pN Peak Force

19
PeakForce QNM can quantitatively and unambiguously identify modulus and adhesion variations. Phase imaging and multifrequency imaging techniques cannot. Comparison of the adhesion and phase images clearly shows that the phase contrast is primarily due to adhesion, whereas one might more commonly assume that it reflects modulus variations Section plot illustrates ability to measure the modulus across the polymer layers PFQNM-Height PFQNM-AdhesionPFQNM-Modulus Tapping-Height Tapping-Phase Multilayered polymer film, 10 µm scans Left: PeakForce QNM Right: TappingMode Capabilities Unlike Anything Else 5/8/201519 Bruker Confidential

21
Choose the Right Cantilever/Tip assemblies

23
Early detection of aging cartilage and osteoarthritis in mice and patient samples using atomic force microscopy

24
Local Nanomechanical Motion of the Cell Wall of Saccharomyces cerevisiae Science, 2004, 305, 1147 k=-F/x k: the spring constant of the cell surface

25
A B After the MDCK fixed on the surface, is it becoming Softer or harder?

26
Effect of Dendrimer Generation on the Assembly and Mechanical Properties of DNA/Phosphorus Dendrimer Multilayer Microcapsules Macromolecules 2006, 39, 5479-5483

27
Figure 1 (A) Schematic of the cell compression experiment. (B) Confocal micrograph reveals the typical AFM probe position, that is, above the center of the designated cell. (C) Confocal microscopy snapshots of a single cell under the compression cycle of AFM force curve acquisition. Cell membrane deformation (blue boundaries) followed by the flattening of the nucleus (see pink regions) was captured. At high deformation, the Newton diffraction ring is clearly visible, which demonstrates the microscopic precision in the alignment. Liu, G.-Y. Langmuir 2006, 22, 8151-8155
 Confocal micrograph reveals the typical AFM probe position, that is, above the center of the designated cell. (C) Confocal microscopy snapshots of a single cell under the compression cycle of AFM force curve acquisition. Cell membrane deformation (blue boundaries) followed by the flattening of the nucleus (see pink regions) was captured. At high deformation, the Newton diffraction ring is clearly visible, which demonstrates the microscopic precision in the alignment. Liu, G.-Y. Langmuir 2006, 22,")
28
Figure 2 Typical force versus deformation curves for living (green) and dead (blue) T cells. Insets show the two types of cells upon the addition of 10 L of 4% trypan blue solution, where dead cells turn blue under optical microscopy.

29
Figure 4 (A) Least-squares fitting of the force curves of living cells according to eq 3. (B) Least-squares fitting of the force curves of dead cells using eq 6. (C) Fitting of the cell compression curves for fixed cells using eq 6. The corresponding cell deformation mechanisms are illustrated on the right column, revealing a fluid-filled balloon (A'), a soft and permeable sphere (B'), and a hard and permeable rubber ball model (C'), respectively.
 Least-squares fitting of the force curves of dead cells using eq 6. (C) Fitting of the cell compression curves for fixed cells using eq 6. The corresponding cell deformation mechanisms are illustrated on the right column, revealing a fluid-filled balloon (A ), a soft and permeable sphere (B ), and a hard and permeable rubber ball model (C ), respectively..")
30
For Living cells

33
For Dead Cells and Fixed Cells The bending of the membrane and cell interior compression (e.g., cytoskeleton and nuclei) are the main contributions to the force- deformation correlation. The small bending of the spherical membrane can be estimated following the example of living cells (see eq 1): Is also very small
: Is also very small.")
34
Stretching and breaking duplex DNA by chemical force microscopy Chemistry & Biology 1997, Vol4, 519

35
A schematic illustration of the chemical force microscopy setup. The sample is mounted on a piezoceramic transducer (green) that can move the sample vertically with subangstrom precision. The probe tip is attached to a flexible cantilever (yellow) and the laser beam (red) is reflected off the cantilever back onto a photodiode to monitor cantilever deflection. The inset shows a cartoon representation of interactions between two complementary strands immobilized on the tip and sample surfaces. Both the tip and the sample are coated with gold; self-assembled monolayers of lkanethiols (blue) are then formed on the gold surface. The DNA shown in the inset corresponds to the relaxed B-DNA conformation.
 that can move the sample vertically with subangstrom precision. The probe tip is attached to a flexible cantilever (yellow) and the laser beam (red) is reflected off the cantilever back onto a photodiode to monitor cantilever deflection. The inset shows a cartoon representation of interactions between two complementary strands immobilized on the tip and sample surfaces. Both the tip and the sample are coated with gold; self-assembled monolayers of lkanethiols (blue) are then formed on the gold surface. The DNA shown in the inset corresponds to the relaxed B-DNA conformation..")
36
Mixed monolayers of thiohexadecanol and hexadecanethiol functionalized with oligonucleotides were formed by spontaneous self-assembly from 3:l H2O:EtOH solutions. The resulting hydroxylterminated monolayer has a hydrophilic surface from which the DNA strands protrude.

37
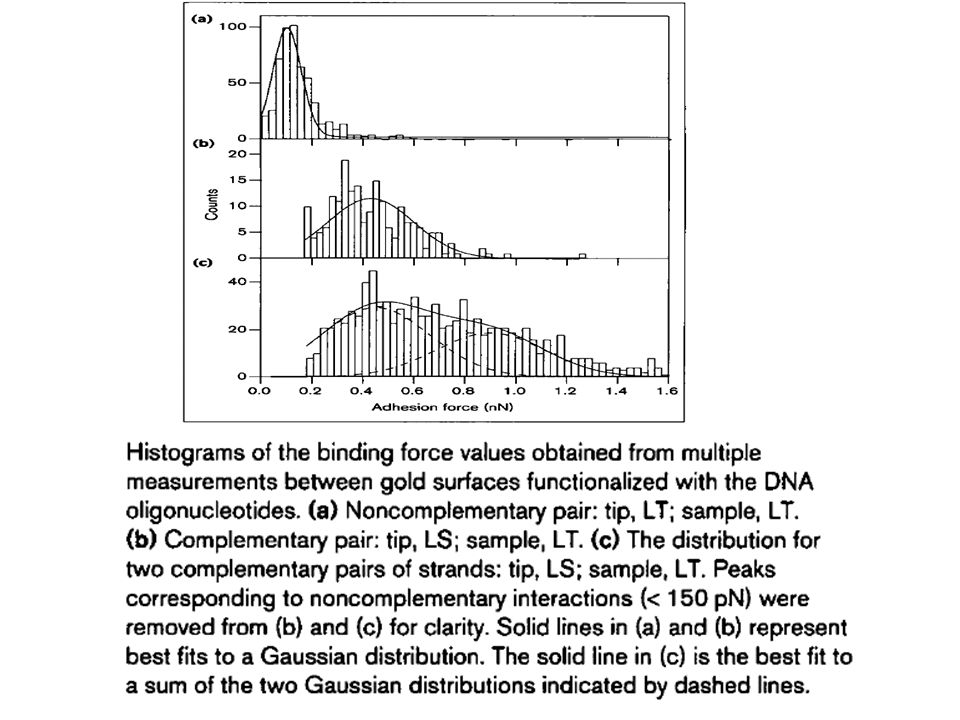
Trace I: noncomplementary pair in which the tip and surface were functionalized with LS. Trace II: complementary pair in which the tip was functionalized with LT and the surface was functionalized with LS.

41
Force versus separation plots obtained from individual approach withdrawal traces for interactions between complementary DNA strands.

43
Sequence-dependent mechanics of single DNA molecules Gaub H. E. Nat. Struct. Biol.1999, 6 346 1 Force-induced melting transition in - DNA. a, Stretching (red) and relaxation (blue) curve of a 1.5- m-long segment of a -BstE II digested DNA molecule. The superimposed black curve is a force curve taken on ssDNA. Because the ssDNA strand has a different contour length (~300 nm), the curves were scaled to the same contour length before superposition. Inset: low-force range of a relaxation curve of - DNA previously split in the melting transition (blue curve). The black curve is a fit according to the freely jointed chain model with an additional segment elasticity using the same parameters that have been determined for ssDNA12 (Kuhn length 8 Å, molecular spring constant per unit length 800 pN).
 and relaxation (blue) curve of a 1.5- m-long segment of a -BstE II digested DNA molecule. The superimposed black curve is a force curve taken on ssDNA. Because the ssDNA strand has a different contour length (~300 nm), the curves were scaled to the same contour length before superposition. Inset: low-force range of a relaxation curve of - DNA previously split in the melting transition (blue curve). The black curve is a fit according to the freely jointed chain model with an additional segment elasticity using the same parameters that have been determined for ssDNA12 (Kuhn length 8 Å, molecular spring constant per unit length 800 pN)..")
44
Inset: a sequence of three stretching (red) and relaxation (blue) cycles performed each time on the same molecule (pulling speed: 0.25 m s–1). Partial melting of the double helix occurs during the B-S transition (first trace) and is complete after the melting transition (second and third trace). Reannealing of the two single strands leads to the characteristic shape of the relaxation traces. Superposition of four extension traces of the same molecule taken at different pulling speeds (red, 3 m s–1; green, 1.5 m s–1; blue, 0.7 m s–1; black, 0.15 m s–1).
 and is complete after the melting transition (second and third trace). Reannealing of the two single strands leads to the characteristic shape of the relaxation traces. Superposition of four extension traces of the same molecule taken at different pulling speeds (red, 3 m s–1; green, 1.5 m s–1; blue, 0.7 m s–1; black, 0.15 m s–1)..")
45
Fig. 2 The mechanical compliance of DNA strongly depends on the specific base pairing in the double helix. a, For double-stranded poly(dG-dC) DNA the B-S transition occurs at 65 pN (see arrows), just as in -DNA, while the melting transition is raised to 300 pN, compared to 150 pN in -DNA. b, In duplex poly(dA-dT) DNA the force of the B-S transition is reduced to 35 pN and the strands melt during this transition, so that no distinct melting transition can be observed.
 DNA the B-S transition occurs at 65 pN (see arrows), just as in -DNA, while the melting transition is raised to 300 pN, compared to 150 pN in -DNA. b, In duplex poly(dA-dT) DNA the force of the B-S transition is reduced to 35 pN and the strands melt during this transition, so that no distinct melting transition can be observed..")
46
The self-complementarily of poly(dG-dC) and poly(dA-dT) allows for hairpin formation within the two DNA strands. After force-induced melting of the double helix, base-pairing forces can be directly measured in the strand attached to the tip. Direct measurement of base-pairing forces

47
Three repeated extension (red) and relaxation (blue) curves of one poly(dG- dC) molecule. In the first extension/relaxation cycle, the molecule is extended into the B-S transition. This process is essentially reversible. In the second cycle the molecule is extended partly into the melting transition, which leads to a shortening of the B-S plateau and the appearance of a new plateau at FG-C = 20 pN in the relaxation trace, caused by hairpin formation. This 20 pN plateau reappears as a hairpin- unzipping plateau in the extension trace of the third cycle. Here the molecule is overstretched even more, resulting in further growth of the 20 pN plateau at the expense of the B-S plateau in the relaxation trace.

48
For poly(dA-dT) DNA the zipping/unzipping force is FA-T = 9 pN. The two plots show the stretching (red) and relaxation (blue) of a molecule that had already formed intrastrand hairpins during a previous stretching. The first cycle shows zipping and unzipping of the hairpins. In the second trace the double strand is further converted into intrastrand hairpins. The double helix starts to melt during the B-S transition.
 and relaxation (blue) of a molecule that had already formed intrastrand hairpins during a previous stretching. The first cycle shows zipping and unzipping of the hairpins. In the second trace the double strand is further converted into intrastrand hairpins. The double helix starts to melt during the B-S transition..")
49
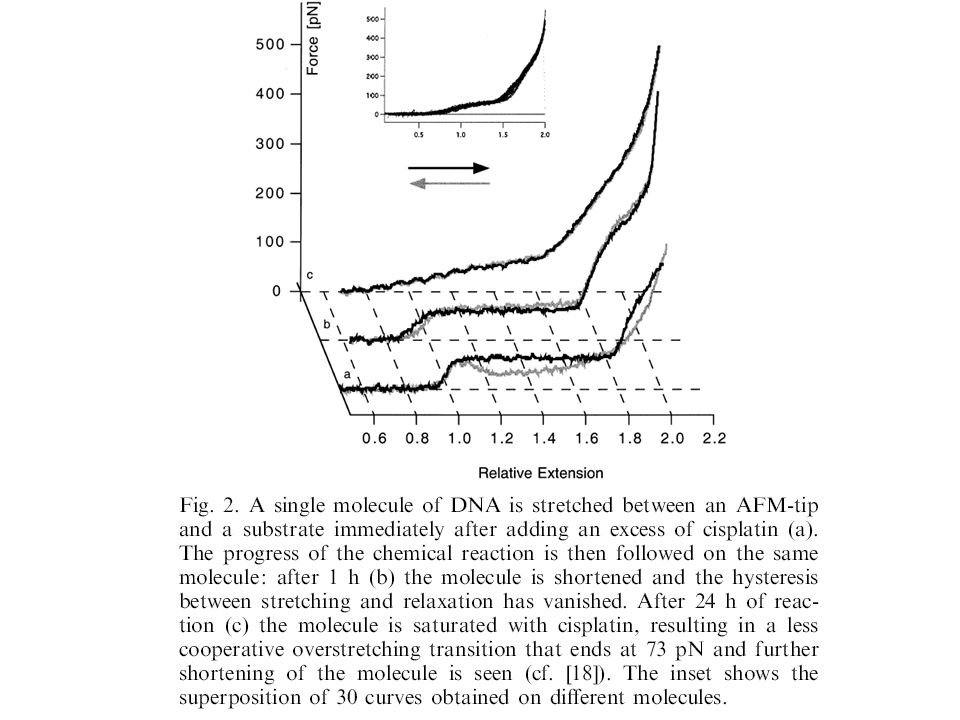
Discriminating small molecule DNA binding modes by single molecule force spectroscopy FEBS Letters 510 (2002) 154 Crosslinker: cisplatin Minor groove binder: Berenil Intercalator: ethidium bromide Drugs may interact with double stranded DNA via a variety of binding modes, each mode giving rise to a specific pharmacological function. This paper demonstrate the ability of single molecule force spectroscopy to discriminate between different interaction modes by measuring the mechanical properties of DNA and their modulation upon the binding of small molecules. Due to the unique topology of double stranded DNA and due to its base pair stacking pattern, DNA undergoes several well-characterised structural transitions upon stretching. In this paper they showed that small molecule binding markedly affects these transitions in ways characteristic to the binding mode and that these effects can be detected at the level of an individual molecule..

54
Homework: How to detect protein/DNA interaction?

Similar presentations




















 AlGaN/GaN quantum well waveguide CD stamper polymer growth surface atoms.>")
 3 History and Definitions in Scanning Probe Microscopy (SPM) History Scanning Tunneling Microscope (STM) Developed.>")

 Will cover all of classes Lec 1-10 plus (qualitatively) on Lec 11–>")
 Real-Space Surface Microscopic Methods.>")





 proteins with Atomic Force Microscopy José L. Toca-Herrera ETSEQ, 02/06/2004.>")
 452-3084 WWW :>")