Download presentation
Presentation is loading. Please wait.

1
Yaniv Loewenstein Computational Biology Seminar, HUJI November 2006 Nuclear Organization

2
The nucleus - introduction The nucleus defines eukaryotes. –No unequivocal evolutionary origin Discrete membrane bound compartment –transcription –RNA processing –splicing

3
Prokaryotes vs. eukaryotes. Multiple linear chromosomes Single cyclic “chromosome” Transcription & translation physically separated Endomembrane compartments (ER)
.")
4
Origin of Eukaryote nucleus Invagination of plasmatic membrane –Nucleus connected to ER Probable endosymbiotic origin –Partially incompatible w. known cell biology –Cytoskeleton proteins based phylogeny Orthopoxvirus ancestor suggested (DNA-pol) Selective force –Splicing\translation decoupling –metabolic compartmentalization (anabolic/catabolic) Still under much debate…(and numerous papers)
 Selective force –Splicing\translation decoupling –metabolic compartmentalization (anabolic/catabolic) Still under much debate…(and numerous papers).")
5
Mitotic nucleus (by the textbook) “chromatin spaghetti” shaped chromosomes nucleus lost nuclei regained
 chromatin spaghetti shaped chromosomes nucleus lost nuclei regained")
6
Talk overview The nucleus - compartments overview –Nuclear domains (D. Spector) –DNA loops and rosettes –Transcription factories? (P. Cook’s) –Nuclear pores & expression regulation (P. Silver) Transcriptional regulation. –Review of experimental results & models (T. Cremer) –A taste of recently published work 3D FISHing – Bolzer et al. Challenge oversimplified text book dogmas.
 –DNA loops and rosettes –Transcription factories. (P. Cook’s) –Nuclear pores & expression regulation (P. Silver) Transcriptional regulation. –Review of experimental results & models (T. Cremer) –A taste of recently published work 3D FISHing – Bolzer et al. Challenge oversimplified text book dogmas..")
7
Nuclear compartments 1.Nuclear envelope & lamina 2.Nuclear Pore Complexes (NPCs) 3.Chromosomal Territories (CT) –DNA is not a random spaghetti 4.Interchromatin Granule Clusters (IGC) –Splicing factor compartments - Speckles (D. Spector) 5.Nucleolus and sub-compartments 6.Others –Cell-type or condition specific.
 5.Nucleolus and sub-compartments 6.Others –Cell-type or condition specific..")
8
The nuclear lamina Nuclear shape maintenance & NPC spacing. Organization of heterochromatin. –Often anchors interphase heterochromatin. DNA replication. Regulation of transcription factors. A scaffolding structure at the nuclear periphery.

9
Nuclear lamina (II) Made of lamins A/B/C intermediate filaments. RNPs involved –RNase somehow disrupts nuclear matrix. Lamins phosphorylated in mitosis - nucleus breaks. –Dephosphorylation promotes chromosome vesicles fusion.

10
Nuclear Pore Complex Exports: –mostly mRNA. Imports –Nuclear proteins & snRNPs –Viruses. –Interacts with importins (karyopherins) Lamina associated cytosol nucleoplasma “I’m an importer/exporter”*
 Lamina associated cytosol nucleoplasma I’m an importer/exporter *.")
11
NPC components (S.cerevisiae). Bapteste et al. Genome Biology 2005 6:R85 A dynamic 50 MDa complex of more than 30 multiple “Nup”s (I'm sorry, the correct answer is “Moops”!)
.")
12
Nuclear Pore Complex (II) Nucleoplasm Cytoplasm Small molecules diffuse Active transport of large macromolecules. Extends 95 nm into the nucleoplasm Suntharalingam and Wente, Dev. Cell 2003.

13
Mammalian Nucleus - EM white – euchromatin (open) black – heterochromatin (condensed) Heterochromatin excluded from pores Heterochromatin concentrates near lamina
 black – heterochromatin (condensed) Heterochromatin excluded from pores Heterochromatin concentrates near lamina")
14
Foisner, R. J Cell Sci 2001;114:3791-3792 Integral membrane protein RFBP interacts with RUSH, a SWI/SNF chromatin remodeling TF. Lamin (+Chromatin) Binding Proteins
 Binding Proteins.")
15
Genome-Wide Localization of the Nuclear Transport Machinery Couples Transcriptional Status and Nuclear Organization Cosalry J, … Silver PA, Cell 2004 Genes relocate from the nucleoplasm to the nuclear pore upon transcriptional induction.

16
GAL genes localize to periphery upon induction Green – GAL loucs (FISH) Red – NPC (periphery) Casolari J et al (Silver PA) Cell 117 (2004) 427-439 % Counts (120 cells) at nuclear periphery
 Red – NPC (periphery) Casolari J et al (Silver PA) Cell 117 (2004) % Counts (120 cells) at nuclear periphery")
17
Casolari J et al (Silver PA) Cell 117 (2004) 427-439 Known NPC components (used by Casolari 2004) Arrows depict known physical interactions
 Cell 117 (2004) Known NPC components (used by Casolari 2004) Arrows depict known physical interactions")
18
NPC binds induced GAL genes GalactoseGlucose 0%100% 0% 100% GAL1,2,7,10 Casolari J et al (Silver PA) Cell 117 (2004) 427-439 Nuclear basket Myosin like (non pore) …...
 Cell 117 (2004) Nuclear basket Myosin like (non pore) …...")
19
Genomic localization (microarray) (in a nutshell) Nuclear transport subcomplexes show similar genome occupancy specificities
 (in a nutshell) Nuclear transport subcomplexes show similar genome occupancy specificities")
20
Correlated Expression & NPC binding

21
RAP1 & NPC binding co-localizes … Rap1 - DNA binding protein associated with Telomeres, Silent mating-type loci Many active genes Boundary activity.

22
Levels of transcription regulation Epigenetics - DNA methylation, nucleosome modifications, insulators etc. DNA sequence alone - can't explain orchestrated activity of thousands of genes. Nucleus architecture - a higher topological level of regulation. ? ?

23
2 nm 11nm 30 nm 300 nm 700 nm 1400 nm 30 nm fiber of packed nucleosomes Chromosomal loops attached to nuclear scaffold Condensed section of metaphase chromosome Entire metaphase chromosome “Beads-on-a-string” Chromatin Packing Double helix 10 5 m 5-10 m ~x10 4 ~x7 ~x100

24
One rosette

25
DNA Rosettes Labrador and Corces, 2002. Cell 111, 151 -154. Nuclear lamina CT Chromosomal Territory (long term silencing) Heterchromain Euchromatin ?
 Heterchromain Euchromatin .")
26
A | Linear layout of interphase chromatin. Yellow - open chromatin Blue – highly condensed chromatin Red –Domains with regulatable insulators under cell differentiation. Loop regulation Labrador and Corces, 2002. Cell 111, 151-154. B | During development, domains of higher-order chromatin structure are organized by active insulators (purple). Inactive insulators and the domain they flank (green) remain in the heterochromatin compartment. C | In a particular tissue, a chromatin domain becomes open after activation of the flanking insulators and vice versa.
. Inactive insulators and the domain they flank (green) remain in the heterochromatin compartment. C | In a particular tissue, a chromatin domain becomes open after activation of the flanking insulators and vice versa..")
27
Looped domains -splicing-proteins green staining (light micrography) Granules may represent splicing machineries.
 Granules may represent splicing machineries.")
28
Loops

29
DNA-loop regulation Back to the text book… Scaffold rich in topoisomerases RNase sensitive lamina.

32
The nucleolus Production and assembly of ribosome components –various small RNA –telomerase function modulation –oncogene regulation A non membranous compartment. Thompson et al, (2003) Science 302 1399-1401 Do tRNA genes affect chromosomes positions?
 Science Do tRNA genes affect chromosomes positions .")
33
tRNAs on yeast chromosomes

34
Nucleolar tRNA localization

35
tRNA genes nucleolar colocalization Thompson et al, (2003) Science 302 1399-1401
 Science")
36
The nucleolus How hundreds of tRNA genes found in many chromosomes are arranged and clustered in the nuclear space?

37
Legend: SUP3 - tRNA gene, URA3 (red) - adjacently inserted gene (non-RNA probe). U14 (green) - nucleolar probe (87% of 715 cells) (52% of 440 cells) tRNA gene localization depends on Pol III complex formation
 - nucleolar probe (87% of 715 cells) (52% of 440 cells) tRNA gene localization depends on Pol III complex formation.")
38
Lessons from the nucleolus Inactivation of the promoter at a single locus removes its nucleolar association. => Nucleolar localization requires tRNA gene transcription-complex formation. Organization of tRNA genes profoundly affects the spatial genome packaging. Are gene types coordinated in 3D to regulate transcription? –Nuclear structure prediction from gene activity?

39
P Cook experiment D. Spector – Speckles & others.

40
Spector, D. L. J Cell Sci 2001;114:2891-2893

41
Splicing factor granules - speckles Differential distribution of factors involved in pre-mRNA processing in the yeast cell nucleus. Potashkin, J.A., …, Spector, D.L. 1990. MCB. 10: 3524-3534. Associations between distinct pre-mRNA splicing components and the cell nucleus. Spector, D.L., …, Maniatis, T. 1991. EMBO J. 10: 3467-3481. Nascent pre-mRNA transcripts are associated with nuclear regions enriched in splicing factors. Huang, S. and Spector, D.L. 1991. Genes & Dev., 5: 2288-2302. U1 and U2 snRNAs are present in nuclear speckles. Huang, S. and Spector, D.L. 1992. PNAS 89: 305-308. … ….. …….. 2006

42
Speckles in the IC space Green – splicing factors snRNPs ABs Blue (DAPI) DNA Speckled pattern Cajal bodies diffused in the nucleoplasm Speckles occur in nuclear specific regions containing little or no DNA. Lamond & Spector. 2003. Nature Rev. Mol. Cell Biol. 4, 605-612

43
Nuclear sub-compartments Nucleolus - rRNA synthesis (various subcomp.) Cajal bodies –snRNP biogenesis (e.g. U1,2, 4-6). –Trafficking to speckles (snRNPs) or nucleoli (snoRNPs). Gems – snRNP maturation. Cleavage bodies – cleavage & poly-A proteins foci. Perinucleolar compartment (PNC) –small RNAs –Predominantly found in cancer cells. PML bodies – associated with various cancers. Spector DL. J Cell Sci. 2001, 114(Pt 16):2891-3.
. –Trafficking to speckles (snRNPs) or nucleoli (snoRNPs). Gems – snRNP maturation. Cleavage bodies – cleavage & poly-A proteins foci. Perinucleolar compartment (PNC) –small RNAs –Predominantly found in cancer cells. PML bodies – associated with various cancers. Spector DL. J Cell Sci. 2001, 114(Pt 16):")
44
Cell type specific domains GATA-1 nuclear bodies (GATA TF) –cell type specific to murine haemopoietic cells –not active in transcription Heat Shock Factor 1 (HSF1 TF) foci –physiological state specific for HS cells –not in HSP70/90 or HSP90 transcription sites Additional levels of transcriptional regulation? Spector DL. J Cell Sci. 2001, 114(Pt 16):2891-3.
:")
45
P Cook experiment P Cook’s experiment

46
Transcription factories & fixed pol? Permeabilized human nucleus of HeLa cells Red - TOTO-3 stains DNA, Green - bromo-UTP nascent RNA transcripts. Cook P. 2002. Nature Genetics 32, 347–52

47
Transcription factories Cook P. 2002. Nature Genetics 32, 347–52 Foci concentrated transcripts. # foci << # active genes # foci << # polymerases ==> “Transcription factories” Similar to bacteria nucleoids. Pol aggregates + RNA interactions (inhibited by RNase).
..")
48
A fixed polymerase? Cook P. 2002. Nature Genetics 32, 347–52 Experimental result Kinetics consistent with the existence of loops of 7.5−175 kb The “untwining problem” – no known mechanism.

49
Lamond & Spector. 2003. Nature Rev. Mol. Cell Biol. 4, 605-612 Speckles created by PPI of pre-mRNA splicing factors. Basal level of factor exchange with nucleoplasmic pool, regulated by phosphorilation. Cell-type-specific (de)phosphorylation. Phosphorylation level modulation of speckle proteins results in an increased release and recruitment to transcription sites. Regulated-exchange model.
phosphorylation. Phosphorylation level modulation of speckle proteins results in an increased release and recruitment to transcription sites. Regulated-exchange model..")
50
P Cook experiment Cremer T’s CT-IC model (review)
")
51
Gathers a large-body of previous experimental work. Understanding gene reg. at the topological level: –Reviews several testable models. –Offers the CT-IC model.

52
Chromosome Territories (CT) Chromosomes occupy discrete territories in the cell nucleus (evidence since the 70s). Methods: FISH detects specific DNA seqs in single cells. –3D positioning of individual (in)active genes –Using various fluorochromes in conjuction. –Secondary coloring (antibodies etc.). S-phase DNA labeling persists in daughter cells –Can be analyzed in EM. –One patch per chromatid
active genes –Using various fluorochromes in conjuction. –Secondary coloring (antibodies etc.). S-phase DNA labeling persists in daughter cells –Can be analyzed in EM. –One patch per chromatid.")
53
Cremer T et al. Nat Rev Genet. 2001 Apr;2(4):292-301. Sponge-like CT structure. Accessible interchromatin invaginations.

55
Silver PA Present the model + predictions

56
CT–IC model – supporting structural features Cremer T et al. Nat Rev Genet. 2001 Apr;2(4):292-301. a | A giant chromatin loop with several active genes (red) expands from the CT surface into the IC space. b | Top, actively transcribed genes (white) are located on a chromatin loop that is remote from centromeric heterochromatin. Bottom, recruitment of the same genes (black) to the centromeric heterochromatin leads to their silencing Short (p) arm A living HeLa cell nucleus. long (q) arm
: a | A giant chromatin loop with several active genes (red) expands from the CT surface into the IC space. b | Top, actively transcribed genes (white) are located on a chromatin loop that is remote from centromeric heterochromatin. Bottom, recruitment of the same genes (black) to the centromeric heterochromatin leads to their silencing Short (p) arm A living HeLa cell nucleus. long (q) arm.")
57
CT–IC model – supporting structural features Cremer T et al. Nat Rev Genet. 2001 Apr;2(4):292-301. c | CTs have variable chromatin density (dark brown, high density; light yellow, low density). Loose chromatin expands into the IC, whereas the most dense chromatin is remote from the IC. A living HeLa cell nucleus.
: c | CTs have variable chromatin density (dark brown, high density; light yellow, low density). Loose chromatin expands into the IC, whereas the most dense chromatin is remote from the IC. A living HeLa cell nucleus..")
58
CT–IC model – supporting structural features Cremer T et al. Nat Rev Genet. 2001 Apr;2(4):292-301. A living HeLa cell nucleus. d | CT showing early-replicating chromatin domains (green) and mid-to-late-replicating chromatin domains (red). Each domain comprises 1 Mb. Gene-poor chromatin (red), is preferentially located at the nuclear periphery and in close contact with the nuclear lamina (yellow), as well as with infoldings of the lamina and around the nucleolus (nu). Gene-rich chromatin (green) is located between the gene-poor compartments.
: A living HeLa cell nucleus. d | CT showing early-replicating chromatin domains (green) and mid-to-late-replicating chromatin domains (red). Each domain comprises 1 Mb. Gene-poor chromatin (red), is preferentially located at the nuclear periphery and in close contact with the nuclear lamina (yellow), as well as with infoldings of the lamina and around the nucleolus (nu). Gene-rich chromatin (green) is located between the gene-poor compartments..")
59
CT–IC model – supporting structural features Cremer T et al. Nat Rev Genet. 2001 Apr;2(4):292-301. A living HeLa cell nucleus. e | Higher-order chromatin structures built up from a hierarchy of chromatin fibres. Inset: this topological view of gene regulation indicates that active genes (white dots) are at the surface of convoluted chromatin fibres. Silenced genes (black dots) may be located towards the interior of the chromatin structure. f | The CT–IC model predicts that the IC (green) contains complexes (orange dots) and larger non-chromatin domains (aggregations of orange dots) for transcription, splicing, DNA replication and repair.
: A living HeLa cell nucleus. e | Higher-order chromatin structures built up from a hierarchy of chromatin fibres. Inset: this topological view of gene regulation indicates that active genes (white dots) are at the surface of convoluted chromatin fibres. Silenced genes (black dots) may be located towards the interior of the chromatin structure. f | The CT–IC model predicts that the IC (green) contains complexes (orange dots) and larger non-chromatin domains (aggregations of orange dots) for transcription, splicing, DNA replication and repair..")
60
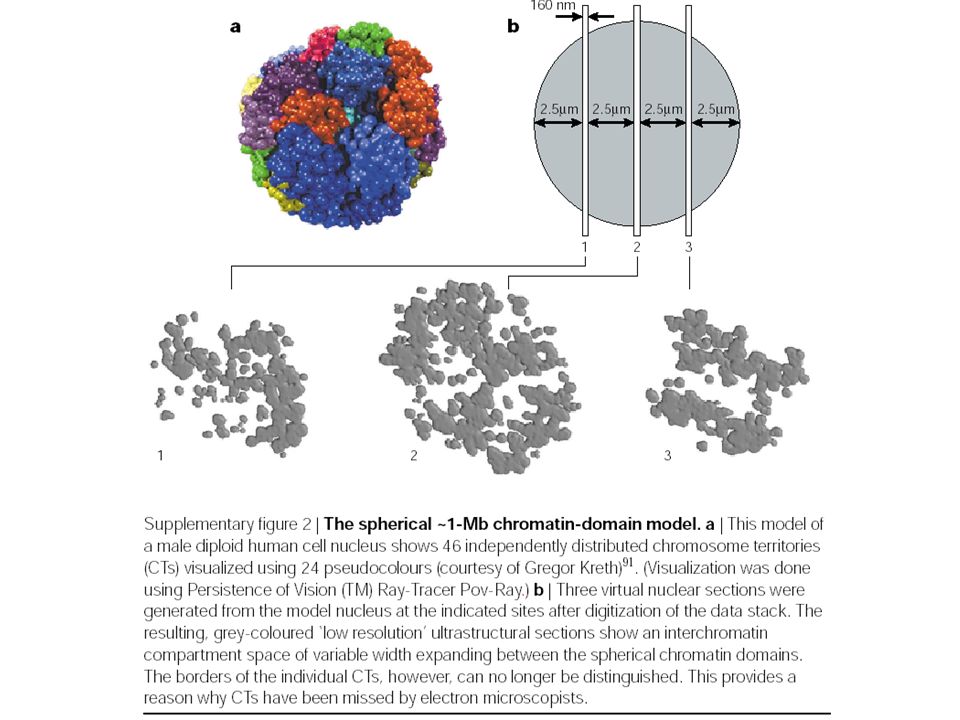
CT–IC model – supporting structural features Cremer T et al. Nat Rev Genet. 2001 Apr;2(4):292-301. A living HeLa cell nucleus. g | CT with 1-Mb chromatin domains (red) and IC (green) expanding between these domains. Inset: the topological relationships between the IC, and active and inactive genes. The finest branches of the IC end between 100-kb chromatin domains. Top: active genes (white dots) are located at the surface of these domains, whereas silenced genes (black dots) are located in the interior. Bottom: alternatively, closed 100-kb chromatin domains with silenced genes are transformed into an open configuration before transcriptional activation.
: A living HeLa cell nucleus. g | CT with 1-Mb chromatin domains (red) and IC (green) expanding between these domains. Inset: the topological relationships between the IC, and active and inactive genes. The finest branches of the IC end between 100-kb chromatin domains. Top: active genes (white dots) are located at the surface of these domains, whereas silenced genes (black dots) are located in the interior. Bottom: alternatively, closed 100-kb chromatin domains with silenced genes are transformed into an open configuration before transcriptional activation..")
61
P Cook experiment Bolzer & Cremer T – Multi-FISH PNAS 2005

62
FISHing in 3D – Bolzer et al A probabilistic 3D order of chromosomes & CTs in quiescent and cycling human fibroblast nuclei. Methods Differential coloring of all 24 chromosome types. –FISH with combinatorial labeling. –Confocal microscopy. –Conditions preserve 3D nucleus shape. 54 nuclei analyzed by “goldFISH”. –A lot of technical details. Bolzer A et al. (Cremer T) PLoS Biol. 2005 May;3(5):e157
 PLoS Biol May;3(5):e157.")
63
Combinatorial coloring (2001) Cremer T et al. Nat Rev Genet. 2001 Apr;2(4):292-301. DAPI color combinations Mutually exclusive CTs (chicken fibroblast) karyotype
: DAPI color combinations Mutually exclusive CTs (chicken fibroblast) karyotype.")
64
Pseudo-coloring (Human – 2005) Bolzer A et al. PLoS Biol. 2005 May;3(5):e157
 Bolzer A et al. PLoS Biol May;3(5):e157")
65
3D image processing & simulations Bolzer A et al. PLoS Biol. 2005 May;3(5):e157
:e157")
66
Multi FISH analysis scheme Promethaphase chromosome “flowers” Bolzer A et al. PLoS Biol. 2005 May;3(5):e157
:e157.")
67
3D reconstruction Bolzer A et al. (Cremer T) PLoS Biol. 2005 May;3(5):e157
 PLoS Biol May;3(5):e157")
68
Radial positions for 54 nuclei Bolzer A et al. PLoS Biol. 2005 May;3(5):e157 CN periphery Nuclei shape and size normalizions North/South etc. can’t be distinguished. Results: 1.short chromosomes prefer the nucleus center. 2.No gene-richness correlation.
:e157 CN periphery Nuclei shape and size normalizions North/South etc. can’t be distinguished. Results: 1.short chromosomes prefer the nucleus center. 2.No gene-richness correlation..")
69
Significantly probabilistic positioning “Chromosome positioning patterns are statistical representations … positions can be described, but contain significant uncertainty”. [Parada, TCB 2003] –Bolzer’s findings agree. –Disagree w. reported fibroblasts precise positioning. Does’nt exclude some extent of determinism – Gene-poor chromatin located beneath the nuclear envelope of all reported cell types. –Nucleoli positioning combines probabilistic and deterministic patterns. Bolzer A et al. PLoS Biol. 2005 May;3(5):e157
:e157.")
70
Bolzer PLoS 2005 - discussion Findings in fibroblasts with flat ellipsoidal nuclei contradict previous studies in cells with rounder nuclei. Central positioning of gene-rich CTs in lymphocytes. Cell-type-specific can’t be explained solely by geometrical constrains of nucleus shape. –3D Simulations. A common rule relating GE to nuclear position: –Gene-poor chromosomes located on nuclear envelope. Questions for future experiments –Does nucleus shape enforce CT arrangements or vice versa? –Nuclear shape causally connected with expression changes? Bolzer A et al. PLoS Biol. 2005 May;3(5):e157
:e157.")
71
Nat Genet. 2006 Aug;38(8):936-41.
:")
72
RNAi & chromatin insulators RNAi required for establishing centromeric chromatin in S. pombe and D. melanogaster. –Nuclear positioning of silent chromatin. Defects in telomere clustering in S. pombe Insulator proteins physical interaction lost when RNase added. RNAi mutants show large phenotypic loss. –“Insulator bodies” lost. Chromatin condensation affected. –Transgenes introduction => phenotypic rescue.

73
CP190 RM62

74
Transcription regulation - summary Chromosomal territories: Probabilistic but non-random chromosome location. chromatin condensation regulated by nuclear location. –Sequence based? –Regulatory insulators Sequence and factor dependent? RNA interactions (and RNAi) Exposure to nucleic factors in interchromatin –DNA topology - Rosettes and loops –Differential factor concentrations. –Transcription factories. Can they be predicted from sequence? mRNA nuclear export regulation (not covered)
 Exposure to nucleic factors in interchromatin –DNA topology - Rosettes and loops –Differential factor concentrations. –Transcription factories. Can they be predicted from sequence. mRNA nuclear export regulation (not covered).")
75
transcription regulation (revisited) Epigenetics - DNA methylation, nucleosome modifications, insulators etc. DNA sequence alone - can't explain orchestrated activity of thousands of genes. Nucleus architecture - a higher topological level of regulation. ? ?

Similar presentations







 genes, and duplicated or diverged genes in gene families.>")













