Download presentation
Presentation is loading. Please wait.

1
Lecture 27 Exam on Friday Up to glyoxalate cycle

2
For the exam Metabolism Glycolysis Fermentation Gluconeogensis
Glycogen Pentose Phosphate Pathway TCA Mechanisms! Regulation of pathways!

3
Page 851

4
Glyoxylate cycle The glyoxylate cycle results in the net conversion of two acetyl-CoA to succinate instead of 4 CO2 in citric acid cycle. Succinate is transferred to mitochondrion where it can be converted to OAA (TCA) Can go to cytosol where it is converted to oxaloacetate for gluconeogenesis. Net reaction 2Ac-CoA + 2NAD+ + FAD OAA + 2CoA + 2NADH +FADH2 + 2H+ Plants are able to convert fatty acids to glucose through this pathway
 Can go to cytosol where it is converted to oxaloacetate for gluconeogenesis. Net reaction. 2Ac-CoA + 2NAD+ + FAD OAA + 2CoA + 2NADH +FADH2 + 2H+ Plants are able to convert fatty acids to glucose through this pathway.")
5
Page 798

6
Electron transport and oxidative phosphorylation
Complete oxidation of glucose by molecular oxygen can be described as: C6H12O6 + 6O2 6CO2 + 6H2O Go’=-2823 kJ/mol Can be broken down into two half-reactions with the transfer of electrons C6H12O6 + 6H2O 6CO2 + 24H+ +24e- 6O2 + 24H+ + 24e- 12H2O 12e- from the oxidation of glucose are not transferred directly to O2, go to NAD+ and FAD to form 10NADH and 2FADH2 These are reoxidized, passing their electrons to the electron- transport chain to reduce O2 to H2O causing the mitochondrion to create a proton gradient. This pH gradient is used to drive the synthesis of ATP via oxidative phosphorylation.

7
Chemiosmotic Theory Electron Transport: Electrons carried by reduced coenzymes are passed through a chain of proteins and coenzymes to drive the generation of a proton gradient across the inner mitochondrial membrane Oxidative Phosphorylation: The proton gradient runs downhill to drive the synthesis of ATP Electron transport is coupled with oxidative phosphorylation It all happens in or at the inner mitochondrial membrane

8
Outer Membrane – Freely permeable to small molecules and ions
Outer Membrane – Freely permeable to small molecules and ions. Contains porins with 10 kDa limit Inner membrane – Protein rich (4:1 protein:lipid). Impermeable. Contains ETR, ATP synthase, transporters. Cristae – Highly folded inner membrane structure. Increase surface area. Matrix- “cytosol” of the mitochondria. Protein rich (500 mg/ml) Contains TCA cycle enzymes, pyruvate dehydrogenase, fatty and amino acid oxidation pathway, DNA, RNA, ribosomes Intermembrane Space – composition similar to cytosol
. Impermeable. Contains ETR, ATP synthase, transporters. Cristae – Highly folded inner membrane structure. Increase surface area. Matrix- cytosol of the mitochondria. Protein rich (500 mg/ml) Contains TCA cycle enzymes, pyruvate dehydrogenase, fatty and amino acid oxidation pathway, DNA, RNA, ribosomes. Intermembrane Space – composition similar to cytosol.")
9
Mitochondrial transport
The inner membrane is impermeable to hydrophilic substances. Has special transport systems for the following: Glycolytically produced cytosolic NADH. Mitochondrially produced metabolites (OAA, acetyl-CoA) for cytosolic glucose formation and fatty acid biosynthesis. Mitochondrially produced ATP must go to cytosol where ATP-utilizing reactions take place. Example: cytoplasmic shuttle systems transport NADH across inner membrane.
 for cytosolic glucose formation and fatty acid biosynthesis. Mitochondrially produced ATP must go to cytosol where ATP-utilizing reactions take place. Example: cytoplasmic shuttle systems transport NADH across inner membrane.")
10
“Transport” of NADH across inner mito membrane

11
Malate-aspartate shuttle
2 phases Phase A Cytosolic NADH reduces OAA to malate (malate DH). Malate--ketoglutarate carrier transports malate from cytosol to mitochondrial matrix, exchanged for -KG In the matrix, NAD+ reoxidizes malate to make OAA and yield NADH. Phase B 4. Transaminase converts OAA to Asp and Glu to -KG. The glutamate-apartate carrier transports Asp from the matrix in exchange for Glu. Transaminase in cytosol converts Glu to Asp. 3 ATPs for every NADH but loses 0.3 ATP because each NADH enters matrix with proton (yield 2.7 ATP).
. Malate--ketoglutarate carrier transports malate from cytosol to mitochondrial matrix, exchanged for -KG. In the matrix, NAD+ reoxidizes malate to make OAA and yield NADH. Phase B. 4. Transaminase converts OAA to Asp and Glu to -KG. The glutamate-apartate carrier transports Asp from the matrix in exchange for Glu. Transaminase in cytosol converts Glu to Asp. 3 ATPs for every NADH but loses 0.3 ATP because each NADH enters matrix with proton (yield 2.7 ATP).")
12
Glycerol phosphate shuttle

13
Glycerophosphate shuttle
Simpler but less energy efficient than the malate-aspartate shuttle. Supplies electrons in a manner similar to succinate dehydrogenase Approx. 2 ATP per NADH, about 0.7 ATP less than the malate-aspartate shuttle. Advantage-irreversible so it operates efficiently even when cytoplasmic NADH is low relative to NAD+. Malate-aspartate shuttle is reversible - driven by concentration gradients.

15
Reduction Potentials High Eo' indicates a strong tendency to be reduced Crucial equation: Go' = -nF Eo' Eo' = Eo'(acceptor) - Eo'(donor) NADH + ½ O2 + H+ NAD++ H+ + H2O n=# of e- transferred per mol reactants, F=Faraday constant NAD++ H+ + 2e- NADH Eo’ = -0.32 ½ O2 + 2e- + 2H+ H2O Eo’ = V Go‘= -nF(Eo'(O2) - Eo'(NADH)) Go‘= -nF(0.82 –(-0.32)) = -nF(1.14) = -2(96.5 kJ mol-1V-1)(1.136) = -220 kJ mol-1
 - Eo (donor) NADH + ½ O2 + H+ NAD++ H+ + H2O. n=# of e- transferred per mol reactants, F=Faraday constant. NAD++ H+ + 2e- NADH Eo’ = ½ O2 + 2e- + 2H+ H2O Eo’ = V. Go‘= -nF(Eo (O2) - Eo (NADH)) Go‘= -nF(0.82 –(-0.32)) = -nF(1.14) = -2(96.5 kJ mol-1V-1)(1.136) = -220 kJ mol-1.")
16
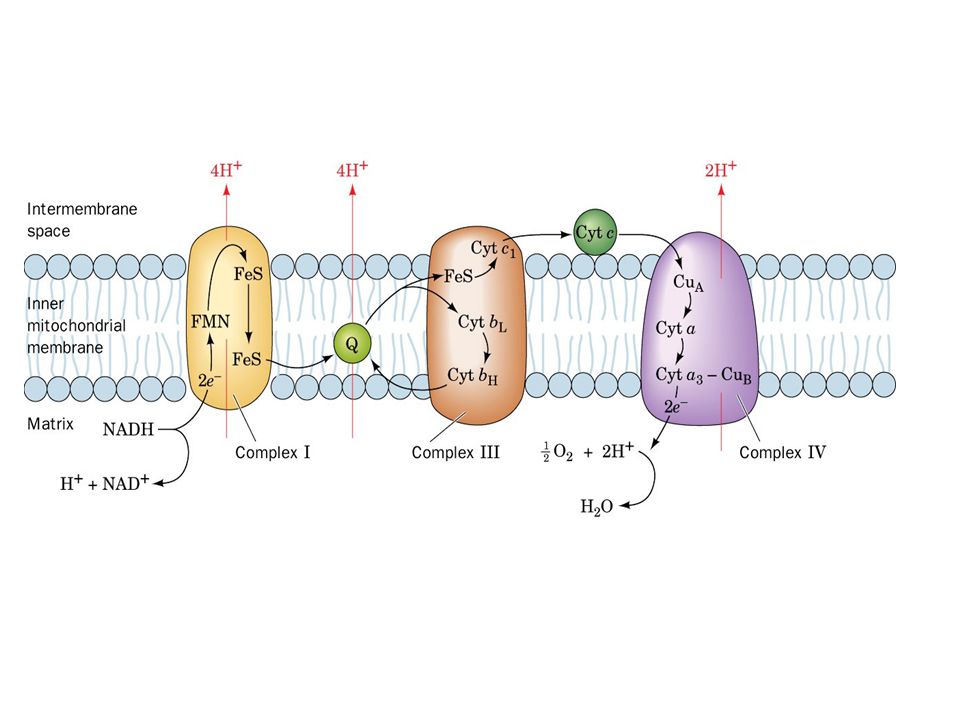
Figure 22-9 The mitochondrial electron-transport chain.
Page 803

17
Electron Transport Four protein complexes in the inner mitochondrial membrane A lipid soluble coenzyme (UQ, CoQ) and a water soluble protein (cyt c) shuttle between protein complexes Electrons generally fall in energy through the chain - from complexes I and II to complex IV
 and a water soluble protein (cyt c) shuttle between protein complexes. Electrons generally fall in energy through the chain - from complexes I and II to complex IV.")
18
Standard reduction potentials of the major respiratory electron carriers.

19
Cofactors of the electron transport chain
Fe-S clusters Coenzyme Q (ubiquinone) Flavin mononucleotide FAD Cytochrome a Cytochrome b Cytochrome c CuA CuB
 Flavin mononucleotide. FAD. Cytochrome a. Cytochrome b. Cytochrome c. CuA. CuB.")
20
Iron-sulfur clusters 4 main types of iron sulfur clusters
[2Fe-2S] and [4Fe-4S] cluster coordinated by 4 Cys SH [3Fe-4S] is a [4Fe-4S] lacking one Fe atom. [Fe-S] is only found in bacteria, liganded to 4 Cys Rieske iron-sulfur proteins [2Fe-2S] cluster but 1 Fe is coordinated by 2 His. Oxidized and reduced states of all Fe-S clusters differ by one formal charge.

21
Figure 22-15a. Structures of the common iron–sulfur clusters
Figure 22-15a Structures of the common iron–sulfur clusters. (a) [Fe–S] cluster. Page 808
 [Fe–S] cluster. Page 808.")
22
Figure 22-15b. Structures of the common iron–sulfur clusters
Figure 22-15b Structures of the common iron–sulfur clusters. (b) [2Fe–2S] cluster. Page 808
 [2Fe–2S] cluster. Page 808.")
23
Figure 22-15c. Structures of the common iron–sulfur clusters
Figure 22-15c Structures of the common iron–sulfur clusters. (c) [4Fe–4S] cluster. Page 808
 [4Fe–4S] cluster. Page 808.")
24
Figure 22-16 X-Ray structure of ferredoxin from Peptococcus aerogenes.
Page 809

25
Figure 22-17a Oxidation states of the coenzymes of complex I. (a) FMN.
Can accept or donate 1 or 2 e- Page 810

26
Figure 22-17b Oxidation states of the coenzymes of complex I. (b) CoQ.
Coenzyme Q’s hydrophobic tail allows it to be soluble in the inner membrane lipid bilayer. Page 810

27
Figure 22-21a. Visible absorption spectra of cytochromes
Figure 22-21a Visible absorption spectra of cytochromes. (a) Absorption spectrum of reduced cytochrome c showing its characteristic a, b, and g (Soret) absorption bands. Page 813
 Absorption spectrum of reduced cytochrome c showing its characteristic a, b, and g (Soret) absorption bands. Page 813.")
28
Figure 22-21b The three separate bands in the spectrum of beef heart mitochondrial membranes indicate the presence of cytochromes a, b, and c. Page 813

29
Figure 22-22a Porphyrin rings in cytochromes. (a) Chemical structures.
Page 813

30
Figure 22-22b. Porphyrin rings in cytochromes
Figure 22-22b Porphyrin rings in cytochromes. (b) Axial liganding of the heme groups contained in cytochromes a, b, and c are shown. Page 813
 Axial liganding of the heme groups contained in cytochromes a, b, and c are shown. Page 813.")
32
Complex I NADH-CoQ Oxidoreductase (NADH dehydrogenase)
Electron transfer from NADH to CoQ More than 30 protein subunits - mass of 850 kD 1st step is 2 e- transfer from NADH to FMN FMNH2 converts 2 e- to 1 e- transfer 6-7 FeS clusters. Four H+ transported out per 2 e- NADH + H+ FMN Fe2+S CoQ NAD+ FMNH2 Fe3+S CoQH2

33
Complex II Succinate-CoQ Reductase
Contains the succinate dehydrogenase (from TCA cycle!) four subunits Two largest subunits contain 2 Fe-S proteins Other subunits involved in binding succinate dehydrogenase to membrane and passing e- to Ubiquinone FAD accepts 2 e- and then passes 1 e- at a time to Fe-S protein No protons pumped from this step Succinate FAD Fe2+S CoQ Fumarate FADH2 Fe3+S CoQH2
 four subunits. Two largest subunits contain 2 Fe-S proteins. Other subunits involved in binding succinate dehydrogenase to membrane and passing e- to Ubiquinone. FAD accepts 2 e- and then passes 1 e- at a time to Fe-S protein. No protons pumped from this step. Succinate. FAD. Fe2+S. CoQ. Fumarate. FADH2. Fe3+S. CoQH2.")
34
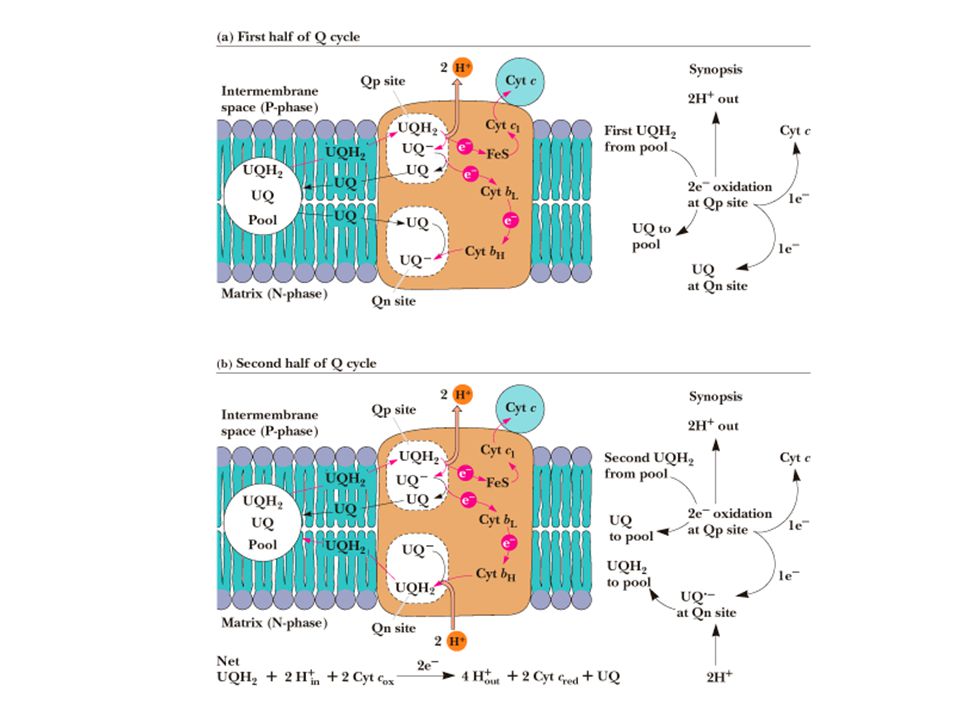
Q-Cycle Transfer from the 2 e- carrier ubiquinone (QH2) to Complex III must occur 1 e- at a time. Works by two single electron transfer steps taking advantage of the stable semiquinone intermediate Also allows for the pumping of 4 protons out of mitochondria at Complex III Myxothiazol (antifungal agent) inhibits electron transfer from UQH2 and Complex III. UQ UQ.- UQH2
 inhibits electron transfer from UQH2 and Complex III. UQ. UQ.- UQH2.")
Similar presentations







 Each.>")

 Electron transport. Chemiosmotic Theory Electron Transport: Electrons carried by reduced coenzymes are passed through a chain of.>")

 are finalled in stepwise manner to oxygen.>")








