Download presentation
Presentation is loading. Please wait.

1
ELECTRON TRANSPORT CHAINS & OXIDATIVE PHOSPHORYLATION
CONTENTS: DEFINING ETC/ETP (ELECTRON TRANSPORT CHAIN/PATHWAY) DEFINING REDOX (OXIDATION-REDUCTION REACTIONS) COMPONENTS OF ETC BIOENERGETICS & UNDERSTANDING ETC ROLE OF MITOCHONDRIA IN ETC CHEMIOSMOTIC THEORY OXIDATIVE STRESS MITOCHONDRIAL PATHWAYS OF ETC WHAT IS OXIDATIVE PHOSPHORYLATION? ASSOCIATION WITH KREB’S CYCLE OR CITRIC ACID CYCLE CHEMIOSMOTIC COUPLING SUMMARY
 DEFINING REDOX (OXIDATION-REDUCTION REACTIONS) COMPONENTS OF ETC. BIOENERGETICS & UNDERSTANDING ETC. ROLE OF MITOCHONDRIA IN ETC. CHEMIOSMOTIC THEORY. OXIDATIVE STRESS. MITOCHONDRIAL PATHWAYS OF ETC. WHAT IS OXIDATIVE PHOSPHORYLATION ASSOCIATION WITH KREB’S CYCLE OR CITRIC ACID CYCLE. CHEMIOSMOTIC COUPLING. SUMMARY.")
2
ELECTRON TRANSPORT CHAIN
Electron transport chains (also called electron transfer pathways- ETP) are biochemical reactions that produce ATP, which is the energy currency of life. An ETP is a series of linked membrane-embedded electron carrier molecules that transfer electrons, during a regulated process of redox reactions, from one electron carrier molecule to another releasing and capturing energy for cellular work.
 are biochemical reactions that produce ATP, which is the energy currency of life. An ETP is a series of linked membrane-embedded electron carrier molecules that transfer electrons, during a regulated process of redox reactions, from one electron carrier molecule to another releasing and capturing energy for cellular work.")
3
ETC contd. 3. Only two sources of energy are available to living organisms: oxidation-reduction (redox) reactions to produce ATP in chemotrophs and using sunlight (photosynthesis) for energy production in phototrophs. 4. Both chemotrophs and phototrophs utilize electron transport chains to convert energy into ATP.
 for energy production in phototrophs. 4. Both chemotrophs and phototrophs utilize electron transport chains to convert energy into ATP.")
4
REDOX REACTIONS Cells generate energy by using redox reactions
Redox reactions are chemical reactions in which electrons are transferred from a oxidised donor molecule (reducing agent) e.g. CH3 COOH (acetic acid) to an electron-deficient acceptor molecule (oxidising agent) e.g. CH3 CH2OH (ethyl alcohol). The two processes always occur simultaneously. The reducing agent that donates their electrons becomes oxidised and the oxidising agent that that accept electrons become reduced.
 e.g. CH3 COOH (acetic acid) to an electron-deficient acceptor molecule (oxidising agent) e.g. CH3 CH2OH (ethyl alcohol). The two processes always occur simultaneously. The reducing agent that donates their electrons becomes oxidised and the oxidising agent that that accept electrons become reduced.")
5
REDOX contd. When electrons are transferred energy is lost and cells can capture this energy to do cellular work such as synthesis of ATP – the energy carrier molecule that directly supplies energy used to maintain highly organised cellular structures and function. The underlying force driving these reactions is the Gibbs free energy of the reactants and products. The Gibbs free energy is the energy available (“free”) to do work. Any reaction that decreases the overall Gibbs free energy of a system will proceed spontaneously
 to do work. Any reaction that decreases the overall Gibbs free energy of a system will proceed spontaneously.")
6
SOME BASIC DEFINITIONS
In an electron transfer reaction, an element undergoing oxidation loses electrons, whereas an element gaining Electrons undergoes reduction. Remember the formula: OIL RIG where OIL refers to Oxidation Is Loss and RIG Reduction Is Gain in electrons.

7
Basic concepts When you take hydrogen ions or electrons
away from a molecule, you “oxidize” that molecule. When you give hydrogen ions or electrons to a molecule, you “reduce” that molecule. When you give phosphate molecules to a molecule, you “phosphorylate” that

8
Oxidation-reactions Reactions of metals or any other organic compounds with oxygen to give oxides are labelled as oxidation. In other words, oxidation is addition of oxygen to a compound or removal of hydrogen from a compound Oxidation increases C-O bonds.

9
Reduction reactions The removal of oxygen from metal oxides to give the metals in their elemental forms is labeled as reduction. Whereas, reduction is addition of hydrogen or removal of oxygen; it increases C-H bonds. The more reduced is a molecule, the more H+ atoms and more energy it contains. Fatty acids have more hydrogen atoms than sugars; hence yield more energy when oxidised.

10
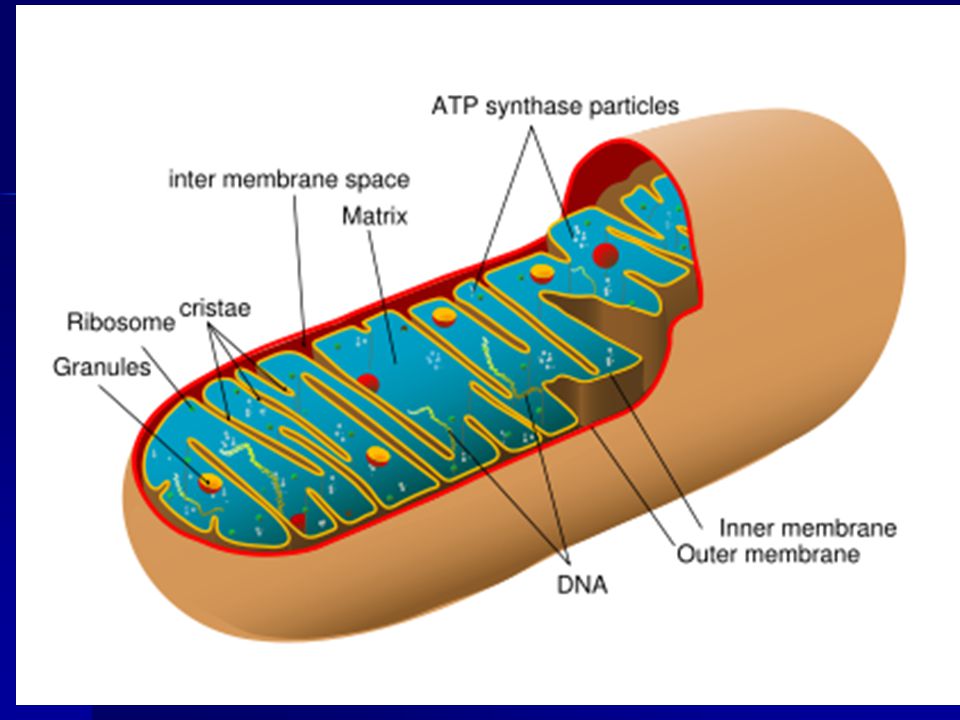
COMPONENTS OF ETC/ETP MITOCHONDRIA ELECTRONS AND PROTONS OXYGEN
NAD+/NADH (redox coenzymes) FLAVIN NUCLEOTIDES (redox coenzymes) COENZYME Q (UBIQUINONE) IRON-SULFUR PROTEINS
 FLAVIN NUCLEOTIDES (redox coenzymes) COENZYME Q (UBIQUINONE) IRON-SULFUR PROTEINS.")
11
BIOENERGETICS The study of bioenergetics involves the processes which reduce nicitinamides and flavin nucleotides, generated from oxidation of carbohydrates and lipids by molecular oxygen via mitochondrial electron-transport chain (ETC) and the mechanism (oxidative phosphorylation) where oxidation is coupled to ATP synthesis. Oxidative phosphorylation is central to metabolism because the free energy of hydrolysis of the ATP generated is used in the synthesis of biomolecules and biological activities such as muscle contraction and transmission of nerve impulses.
 and the mechanism (oxidative phosphorylation) where oxidation is coupled to ATP synthesis. Oxidative phosphorylation is central to metabolism because the free energy of hydrolysis of the ATP generated is used in the synthesis of biomolecules and biological activities such as muscle contraction and transmission of nerve impulses.")
12
ATP synthase & ETC ATP is made by an enzyme called ATP synthase. The structure of this enzyme and its underlying genetic code is remarkably similar in all known forms of life. ATP synthase is powered by a transmembrane electrochemical potential gradient, usually in the form of a proton gradient. The function of the electron transport chain is to produce this gradient. In all living organisms, a series of redox reactions is used to produce a transmembrane electrochemical potential gradient.

13
UNDERSTANDING THE ELECTRON TRANSFER CHAIN
The transfer of electrons from a high-energy molecule (the donor) to a lower-energy molecule (the acceptor) can be spatially separated into a series of intermediate redox reactions. This is an electron transport chain. Electron transport chains produce energy in the form of a transmembrane electrochemical potential gradient. This energy is used to do useful work. The gradient can be used to transport molecules across membranes. It can be used to produce ATP and NADH, high-energy molecules that are necessary for growth. A small amount of ATP is available from substrate-level phosphorylation (for example, in glycolysis). Some organisms can obtain ATP exclusively by fermentation. In most organisms, however, the majority of ATP is generated by electron transport chains.
 to a lower-energy molecule (the acceptor) can be spatially separated into a series of intermediate redox reactions. This is an electron transport chain. Electron transport chains produce energy in the form of a transmembrane electrochemical potential gradient. This energy is used to do useful work. The gradient can be used to transport molecules across membranes. It can be used to produce ATP and NADH, high-energy molecules that are necessary for growth. A small amount of ATP is available from substrate-level phosphorylation (for example, in glycolysis). Some organisms can obtain ATP exclusively by fermentation. In most organisms, however, the majority of ATP is generated by electron transport chains.")
14
ETC IN MITOCHONDRIA The cells of all eukaryotes (all animals, plants, fungi, algae – in other words, all living things except bacteria and archaea) contain intracellular organelles called mitochondria that produce ATP. Energy sources such as glucose are initially metabolized in the cytoplasm. The products are imported into mitochondria. Mitochondria continue the process of catabolism using metabolic pathways including the Krebs cycle, fatty acid oxidation and amino acid oxidation. The end result of these pathways is the production of two energy-rich electron donors, NADH and FADH2.
 contain intracellular organelles called mitochondria that produce ATP. Energy sources such as glucose are initially metabolized in the cytoplasm. The products are imported into mitochondria. Mitochondria continue the process of catabolism using metabolic pathways including the Krebs cycle, fatty acid oxidation and amino acid oxidation. The end result of these pathways is the production of two energy-rich electron donors, NADH and FADH2.")
16
ETC IN MITOCHONDRIA- contd.
Electrons from these donors are passed through an electron transport chain to oxygen, which is reduced to water. This is a multi-step redox process that occurs on the mitochondrial inner membrane. The enzymes that catalyze these reactions have the remarkable ability to simultaneously create a proton gradient across the membrane, producing a thermodynamically unlikely high-energy state with the potential to do work

17
CHEMIOSMOTIC THEORY Peter Mitchell, a Britiah biochemist, in 1961, proposed a mechanism by which the free energy generated during electron transport drives ATP synthesis As electrons pass through the ETC, protons ae transported from the matrix and released into the inter membrane space As a result, an electrical potential and proton gradient (pH) arise across the inner membrane and this elecrochemical proton gradient is often referred as protonmotive force
 arise across the inner membrane and this elecrochemical proton gradient is often referred as protonmotive force.")
18
CHEMIOSMO THEORY contd.
Protons, present in the intermembrane in excess can pass through the inner membrane and back into the matrix down their concentration gradient only through special channels as the inner membrane is impermeable to ions (protons) As the themodynamically favorable flow of protons occur through a channel, each of which contains an ATP synthase activity, an ATP synthesis occurs.
 As the themodynamically favorable flow of protons occur through a channel, each of which contains an ATP synthase activity, an ATP synthesis occurs.")
19
Chemiosmo theory contd.
Uncouplers: A variety of molecules such as, dinitrophenol (DNP) and gramicidin can collapse the proton gradient by equalising the proton concentration on both sides of the membrane and according to the chemiosmotic theory, a disrupted proton gradient dissipates the energy (derived from food) as heat
 and gramicidin can collapse the proton gradient by equalising the proton concentration on both sides of the membrane and according to the chemiosmotic theory, a disrupted proton gradient dissipates the energy (derived from food) as heat.")
21
OXIDATIVE STRESS Oxygen can accept single electrons to form unstable derivatives, referred as reactive oxygen species (ROS) e.g. Hydrogen peroxide Because ROS are so reactive, they can seriously damage living cells if formed in significant ammounts They can be kept to the minimum by antioxidants e.g. Alpha-tocopherol (Vitamin E), beta-Carotene (Vitamin A) that inhibit the reaction of molecules with oxygen radicals.
, beta-Carotene (Vitamin A) that inhibit the reaction of molecules with oxygen radicals.")
22
MITOCHONDRIAL REDOX CARRIERS
Four membrane-bound complexes have been identified in mitochondria. Each is an extremely complex transmembrane structure that is embedded in the inner membrane. Three of them are proton pumps. The structures are electrically connected by lipid-soluble electron carriers and water-soluble electron carriers

23
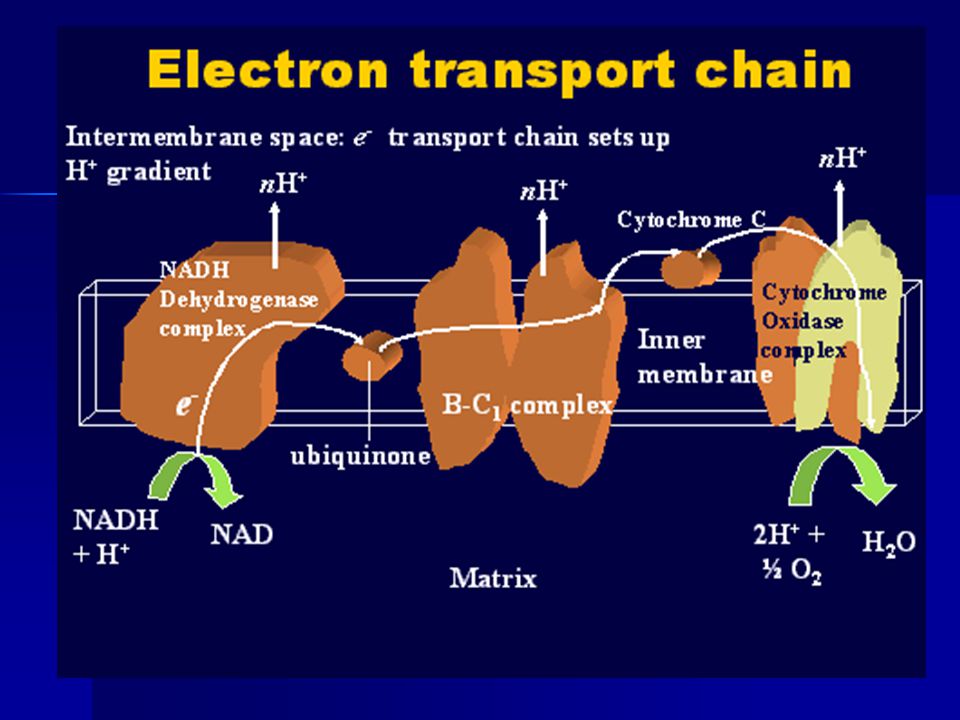
PATHWAYS (COMPLEXES) OF ETC
The overall electron transport chain is: NADH → Complex I → Q → Complex III →cytochrome c → Complex IV → O ↑ Complex II

24
Complex I (NADH dehydrogenase) removes two electrons from NADH and transfers them to a lipid-soluble carrier, ubiquinone (Q). The reduced product, ubiquinol (QH2) is free to diffuse within the membrane. At the same time, Complex I moves four protons (H+) across the membrane, producing a proton gradient. Complex I is also called NADH:ubiquinone oxidoreductase.

26
COMPLEX I NADH (nicotinamide adenine dinucleotide reduced form) is oxidized to NAD+, reducing FMN (flavin mononucleotide) to FMNH2 in one two-electron site The next electron carrier is a Fe-S cluster, which can only accept one electron at a time to reduce the ferric ion into a ferrous ion. Conveniently, FMNH2 can only be oxidized in two one-electron steps, through a semiquinone intermediate.
 is oxidized to NAD+, reducing FMN (flavin mononucleotide) to FMNH2 in one two-electron site. The next electron carrier is a Fe-S cluster, which can only accept one electron at a time to reduce the ferric ion into a ferrous ion. Conveniently, FMNH2 can only be oxidized in two one-electron steps, through a semiquinone intermediate.")
27
COMPEX I contd. The electron thus travels from the FMNH2 to the Fe-S cluster, then from the Fe-S cluster to the oxidized Q to give the free-radical (semiquinone) form of Q. This happens again to reduce the semiquinone form to the ubiquinol form, QH2. During this process, four protons are translocated across the inner mitochondrial membrane, from the matrix to the intermembrane space This creates a proton gradient that will be later used to generate ATP through oxidative phosphorylation.
 form of Q. This happens again to reduce the semiquinone form to the ubiquinol form, QH2. During this process, four protons are translocated across the inner mitochondrial membrane, from the matrix to the intermembrane space. This creates a proton gradient that will be later used to generate ATP through oxidative phosphorylation.")
28
COMPLEX II Complex II (succinate dehydrogenase) is not a proton pump. It serves to funnel additional electrons into the quinone pool (Q) by removing electrons from succinate and transferring them (via FAD) to Q. Other electron donors (e.g. fatty acids and glycerol 3-phosphate) also funnel electrons into Q (via FAD), again without producing a proton gradient.
 is not a proton pump. It serves to funnel additional electrons into the quinone pool (Q) by removing electrons from succinate and transferring them (via FAD) to Q. Other electron donors (e.g. fatty acids and glycerol 3-phosphate) also funnel electrons into Q (via FAD), again without producing a proton gradient.")
29
COMPLEX III Complex III (cytochrome bc1 complex) removes in a stepwise fashion two electrons from QH2 and transfers them to two molecules of cytochrome c, a water-soluble electron carrier located on the outer surface of the membrane. At the same time, it moves four protons across the membrane, producing a proton gradient. When electron transfer is hindered (by a high membrane potential, point mutations or respiratory inhibitors such as antimycin A), Complex III may leak electrons to oxygen resulting in the formation of a superoxide.
 removes in a stepwise fashion two electrons from QH2 and transfers them to two molecules of cytochrome c, a water-soluble electron carrier located on the outer surface of the membrane. At the same time, it moves four protons across the membrane, producing a proton gradient. When electron transfer is hindered (by a high membrane potential, point mutations or respiratory inhibitors such as antimycin A), Complex III may leak electrons to oxygen resulting in the formation of a superoxide.")
30
COMPLEX IV Complex IV (cytochrome c oxidase) removes four electrons from four molecules of cytochrome c and transfers them to molecular oxygen (O2), producing two molecules of water (H2O). At the same time, it moves four protons across the membrane, producing a proton gradient.
 removes four electrons from four molecules of cytochrome c and transfers them to molecular oxygen (O2), producing two molecules of water (H2O). At the same time, it moves four protons across the membrane, producing a proton gradient.")
31
PROTON PUMP MECHNISMS There are three proton pumps: I, III and IV. The resulting transmembrane proton gradient is used to make ATP via ATP synthase. The trans membrane electrochemical gradient acts as the intermediate in the transfer of energy to ATP. Oxidation of NADH results in the production of about 3 molecules per O reduced to water. Oxidation of FADH2 yields 2 ATP’s.

32
WHAT IS OXIDATIVE PHOSPHORYLATION?
When you give phosphate molecules to a molecule, you “phosphorylate” that molecule. So, oxidative phosphorylation (very simply) means the process that couples the removal of hydrogen ions from one molecule and giving phosphate molecules to another molecule.
 means the process that couples the removal of hydrogen ions from one molecule and giving phosphate molecules to another molecule.")
33
MITOCHONDRIA IN OXIDATIVE PHOSPHORYLATION
Why do we need mitochondria? The whole idea behind this process is to get as much ATP out of glucose (or other food products) as possible. If we have no oxygen, we get only 4 molecules of ATP energy packets for each glucose molecule (in glycolysis). However, if we have oxygen, then we get to run the Kreb’s cycle to produce many more hydrogen ions that can run those ATP pumps.
 as possible. If we have no. oxygen, we get only 4 molecules of ATP. energy packets for each glucose molecule. (in glycolysis). However, if we have oxygen, then we get. to run the Kreb’s cycle to produce many. more hydrogen ions that can run those ATP. pumps.")
34
WHY MITOCHONDRIA? From the Kreb’s cycle we get 24-28 ATP
molecules out of one molecule of glucose converted to pyruvate (plus the 4 molecules we got out of glycolysis). So, you can see how much more energy we can get out of a molecule of glucose if our mitochondria are working and if we have oxygen.
. So, you can see how much more energy we. can get out of a molecule of glucose if our. mitochondria are working and if we have. oxygen.")
35
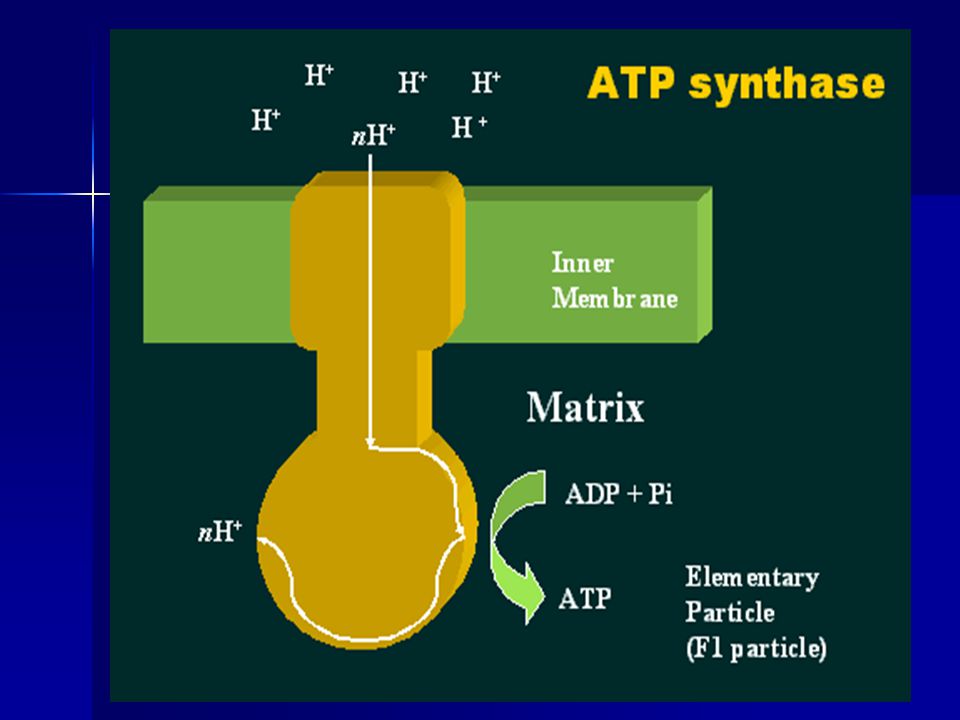
ATP synthase complex This complex (sometimes termed as COMPLEX V of ETC) is found in all energy transducing membranes including that of the mitochondria. It contains a proton transport channel, the only way for the protons to reenter the mitochondrial matrix. The energy proton potential gradient is used in the synthesis of ATP from ADP and Pi
 is found in all energy transducing membranes including that of the mitochondria. It contains a proton transport channel, the only way for the protons to reenter the mitochondrial matrix. The energy proton potential gradient is used in the synthesis of ATP from ADP and Pi.")
37
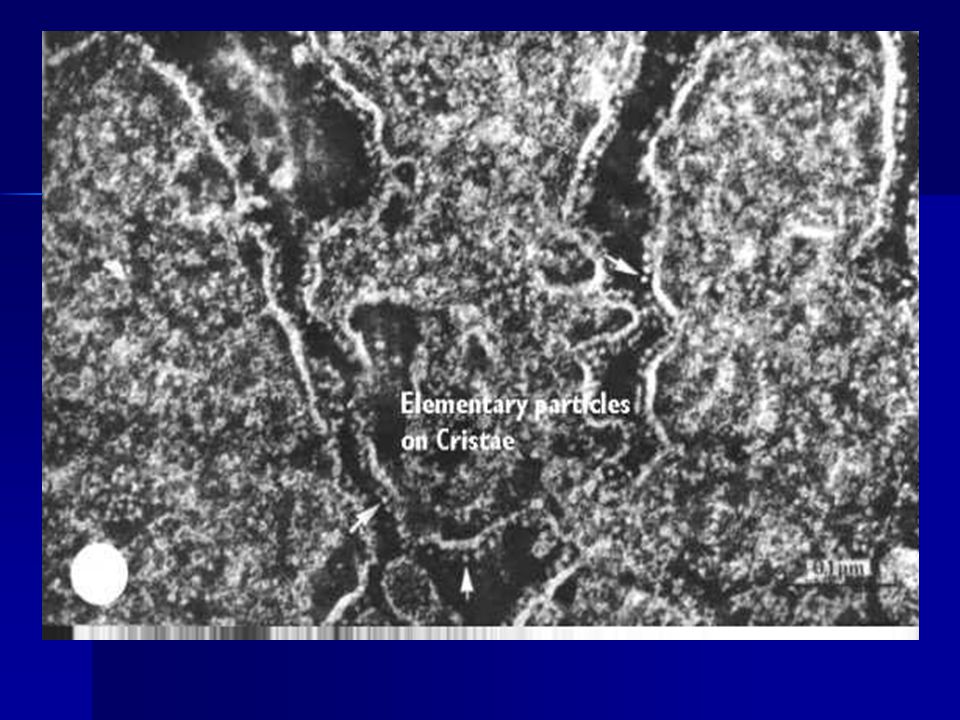
ETC ON CRISTAE

38
IMPORTANCE OF CRISTAE You can now appreciate the importance of the cristae....not only do they contain and organize the electron transport chain and the ATP pumps, they also serve to separate the matrix from the space that will contain the hydrogen ions, allowing the gradient needed to drive the pump

40
ASSOCIATION WITH KREB’S CYCLE OR CITRIC ACID CYCLE
As the Kreb’s cycle runs, hydrogen ions (or electrons) are donated to the two carrier molecules in 4 of the steps. They are picked up by either NAD or FAD and these carrier molecules become NADH and FADH (because they now are carrying a hydrogen ion). The NADH and FADH essentially serve as a ferry in the lateral plane of the membrane diffusing from one complex to the next. At each site is a hydrogen (or proton) pump which transfers hydrogen from one side of the membrane to the other.
 are donated to the two carrier molecules in 4 of the steps. They are picked up by either NAD or FAD and these carrier molecules become NADH and FADH (because they now are carrying a hydrogen ion). The NADH and FADH essentially serve as a ferry in the lateral plane of the membrane diffusing from one complex to the next. At each site is a hydrogen (or proton) pump which transfers hydrogen from one side of the membrane to the other.")
41
KREB’S CYCLE contd. This creates a gradient across the inner membrane with a higher concentration of Hydrogen ions in the inter cristae space (this is the space between the inner and outer membranes). The electrons are carried from complex to complex by ubiquinone and cycochrome C.
. The electrons are carried from complex to complex by ubiquinone and cycochrome C.")
42
OXIDATIVE PHOSPHORYLATION
The chemiosmotic coupling hypothesis, as proposed by Nobel Prize in Chemistry winner Peter D. Mitchell explains that the electron transport chain and oxidative phosphorylation are coupled by a proton gradient across the inner mitochondrial membrane. The efflux of protons creates both a pH gradient and an electrochemical gradient This proton gradient is used by the F0F1 ATP synthase complex to make ATP via oxidative phosphorylation. ATP synthase is sometimes regarded as complex V of the electron transport chain.

43
OXYDAPHOS-contd. The F0 component of ATP synthase acts as an ion channel for return of protons back to mitochondrial matrix. During their return, the free energy produced during the generation of the oxidized forms of the electron carriers (NAD+ and FAD) is released. This energy is used to drive ATP synthesis, catalyzed by the F1 component of the complex. Coupling with oxidaive phosphorylation is a key step for ATP production.
 is released. This energy is used to drive ATP synthesis, catalyzed by the F1 component of the complex. Coupling with oxidaive phosphorylation is a key step for ATP production.")
44
SUMMARY The mitochondrial electron transport chain removes electrons from an electron donor (NADH or FADH2) and passes them to a terminal electron acceptor (O2) via a series of redox reactions. These reactions are coupled to the creation of a proton gradient across the mitochondrial inner membrane. There are three proton pumps: I, III and IV. The resulting transmembrane proton gradient is used to make ATP via ATP synthase.
 and passes them to a terminal electron acceptor (O2) via a series of redox reactions. These reactions are coupled to the creation of a proton gradient across the mitochondrial inner membrane. There are three proton pumps: I, III and IV. The resulting transmembrane proton gradient is used to make ATP via ATP synthase.")
45
Summary contd. The reactions catalyzed by Complex I and Complex III exist roughly at equilibrium. The steady-state concentrations of the reactants and products are approximately equal. This means that these reactions are readily reversible, simply by increasing the concentration of the products relative to the concentration of the reactants (for example, by increasing the proton gradient).
.")
46
Summary contd. ATP synthase is also readily reversible. Thus ATP can be used to make a proton gradient, which in turn can be used to make NADH. This process of reverse electron transport is important in many prokaryotic electron transport chains

47
THANKS The whole art of teaching is only the art of awakening the natural curiosity of young minds for the purpose of satisfying it afterwards.

Similar presentations



 Electron transport. Chemiosmotic Theory Electron Transport: Electrons carried by reduced coenzymes are passed through a chain of.>")
















