Download presentation
Presentation is loading. Please wait.

1
Biology

2
1. Discovery of Genetic Material 2. DNA Structure 3. The Race to Solve the Puzzle of DNA Structure 4.DNA Replication

3
Biologists knew that genes are located on chromosomes (made of DNA and protein) – DNA and protein were the candidates for the genetic material – Until the 1940s, the case for proteins seemed stronger because proteins appeared to be more structurally complex and functionally specific. – Biologists finally established the role of DNA in heredity through studies involving bacteria and the viruses that infect them.

4
1928; British medical officer Griffith was studying two strains of a bacterium: – a pathogenic (disease-causing) strain that cause pneumonia – a harmless strain.
 strain that cause pneumonia – a harmless strain.")
5
Found that when he mixed a dead version of the pathogenic bacteria and harmless bacteria, some living bacterial cells were converted to the disease-causing form. – Furthermore, all of the descendants of the transformed bacteria inherited the newly acquired ability to cause disease.

6
Clearly, some chemical component of the dead bacteria could act as a “transforming factor” that brought about a heritable change.

7
1952; American biologists Experiments showed that DNA is the genetic material of a virus (bacteriophage or phage, for short) called T2, which infects E.coli T2 consists solely of DNA and protein; DNA-containing head and a hollow tail with six jointed fibers extending from it.
 called T2, which infects E.coli T2 consists solely of DNA and protein; DNA-containing head and a hollow tail with six jointed fibers extending from it.")
8
T2 infects bacteria by attaching to the surface with its fibers and injecting its hereditary material. Raised the question: Is it DNA being passed on or protein? To answer protein or DNA question, they devised an experiment to determine what kinds of molecules the phage transferred to E.coli during infection

9
Used a few relatively simple tools: Chemicals containing radioactive isotopes To label the DNA and protein in T2 Used radioactive sulfur and phosphorous Sulfur is in proteins. Phosphorous is in DNA. A radioactivity detector A kitchen blender And a centrifuge (device that spins test tubes to separate particles of different weights.

10
The Experiment 1. First they grew T2 with E.Coli with radioactive sulfur. 2. Then, they grew a separate batch of phages in a solution containing radioactive phosphorous. 3. They allowed the two batches of T2 to infect separate samples of nonradioactive bacteria 4. Shortly after the onset of infection, they agitated the cultures in a blended to shake loose any parts of the phages that remained outside the bacterial cells. 5. They then spun the mixtures in a centrifuge. The cells were deposited as a pellet at the bottom of the centrifuge tubes, but phages and parts of phages being lighter, remained suspended in the liquid. 6. The researchers then measured the radioactivity in the pellet and the liquid.

11
The Results Found that when the bacteria has been infected with T2 phages containing labeled protein, the radioactivity ended up in the liquid but not bacteria. Result suggested that the phage protein did not enter the cells. But when the bacteria had been infected with phages whose DNA was tagged, then most of the radioactivity was in the pellet, made up of bacteria. When these bacteria were returned to liquid growth medium, they soon died and lysed and released new phages that contained radioactive phosphorous in their DNA but no radioactive sulfur.

12
The Conclusions They concluded that T2 injects its DNA into the host cell, leaving virtually all its protein outside. They demonstrated that it is the injected DNA molecules that cause cells to produce additional phage DNA and proteins, making new complete phages. This indicated that DNA contained the instructions for making proteins ***THESE RESULTS CONVINCED MOST SCIENTISTS THAT DNA IS THE HEREDITARY MATERIAL!!!***

14
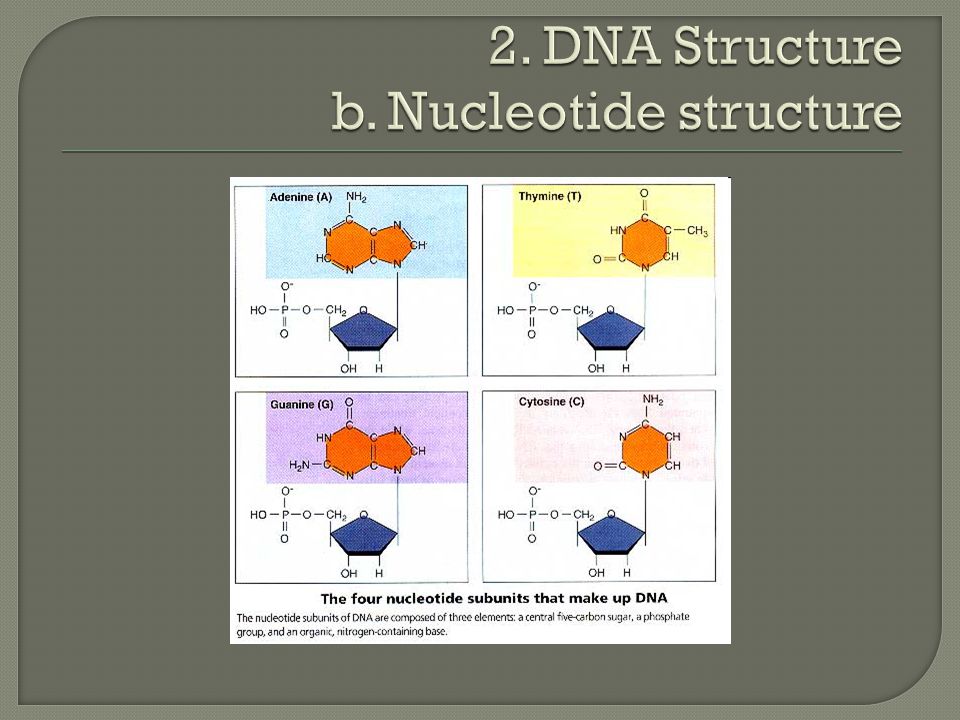
Deoxyribonucleic Acid (DNA) Monomers made up of nucleotides: Nucleotides consist of: A five carbon sugar, deoxyribose Phosphate group Nitrogenous base (Adenine, Guanine, Cytosine, Thymine) Double helix consists of: Sugar-phosphate backbone held by covalent bonds Nitrogen bases are hydrogen bonded together; A pairs with T and C pairs with G
 Monomers made up of nucleotides: Nucleotides consist of: A five carbon sugar, deoxyribose Phosphate group Nitrogenous base (Adenine, Guanine, Cytosine, Thymine) Double helix consists of: Sugar-phosphate backbone held by covalent bonds Nitrogen bases are hydrogen bonded together; A pairs with T and C pairs with G")
16
Nucleotides consist of: A five carbon sugar, deoxyribose Missing one oxygen when compared to ribose Phosphate group Nitrogenous base Adenine, Guanine, Cytosine, Thymine Double ring= purines (A and G) Single ring= pyrimidines (T and C)
 Single ring= pyrimidines (T and C)")
18
DNA’s sugar-phosphate backbones run in opposite directions. Each strands has a 3’ end and a 5’ end. The primed number is referring to the carbon atoms of the nucleotide sugars. At one end of each DNA strand, the sugar’s 3’ carbon atom is attached to an – OH group, at the other end, the sugar’s 5’ carbon has a phosphate group.

21
A few scientists working on the puzzle trying to determine the 3-D structure: Franklin, Watson and Crick Rosalind Franklin observed an X-ray crystallography image of the basic shape of DNA

22
Watson saw this image, and with just a glance deduced the basic shape of DNA to be a helix with a uniform diameter of 2 nm, with its nitrogenous bases stacked about one-third of a nanometer apart. The diameter of the helix suggested it was made up of two polynucleotide strands=> DOUBLE HELIX!

23
Watson and Crick began trying to construct a double helix that would conform both Franklin’s data and what was currently known about the chemistry of DNA. Franklin had concluded that the sugar- phosphate backbones must be on the outside of the double-helix, forcing the nitrogenous bases to swivel to the interior of the molecule.

24
Watson and Crick found that Adenine always paired with Thymine, and Guanine and Cytosine, to ensure a uniform diameter. Complementary base pairing was explained both by the physical attributes and chemical bonding of DNA, along with data obtained by Chargaff

25
Chargaff’s rules: A always pairs with T and G always pairs with C. Only apply to base pairing, not the sequence of nucleotides The sequence of bases can vary in countless ways, and each gene has a unique order of nucleotides, or base sequence

26
1962, Watson and Crick received the Nobel Prize for their work (Franklin would have received it as well, but she died from cancer in 1958; Nobel Prizes are never awarded to the deceased)
")
27
“It has not escaped our notice that the specific pairing we have postulated immediately suggests a possible copying mechanism for the genetic material” ~Watson and Crick

28
Logic behinds Watson-Crick’s proposal for how DNA is copied Can be seen by covering one of the strands in the parental DNA molecule with a piece of paper: you can determine the bases of the covered strand by applying the base-pairing rules: A pair with T, and G pairs with C. Watson and Crick predicted that a cell applies the same rules when copying its genes.

29
Figure 10.4A Template model for DNA replication 1. First, the two strands of parental DNA separate, and each becomes a template for the assembly of a complementary strand from a supply of free nucleotides. 2. The nucleotides line up one at a time along the template strand in accordance with the base- pairing rules 3. Enzymes link the nucleotides to form the new DNA strands. 4. Completed new molecules, identical to the parental molecule, are known as daughter DNA.

30
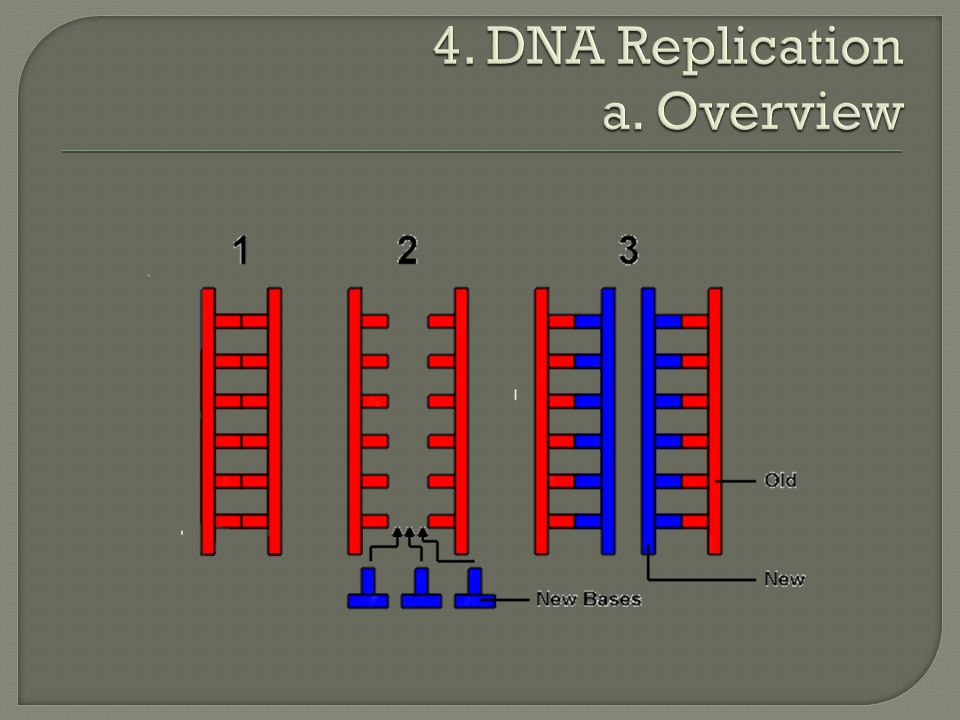
Semi-conservative model Watson and Crick’s model predicts that when a double helix replicates, each of the two daughter molecules will have one old strand, which was part of the parental molecule, and one new made strand. Known as semi-conservative model because half of the parental molecule is maintained (conserved) in each daughter molecule. Confirmed by experiments performed in the 1950s.
 in each daughter molecule. Confirmed by experiments performed in the 1950s..")
32
The opposite orientation of the strands is important in DNA replication. DNA polymerases link DNA nucleotides to a growing daughter strand, only to the 3’ end of the strand, never to the 5’ end. Thus, a daughter DNA strand can only grow in the 5’ 3’ direction.

33
We will be looking at three steps in DNA Repication: 1. Initiation 2. Elongation 3. Termination

34
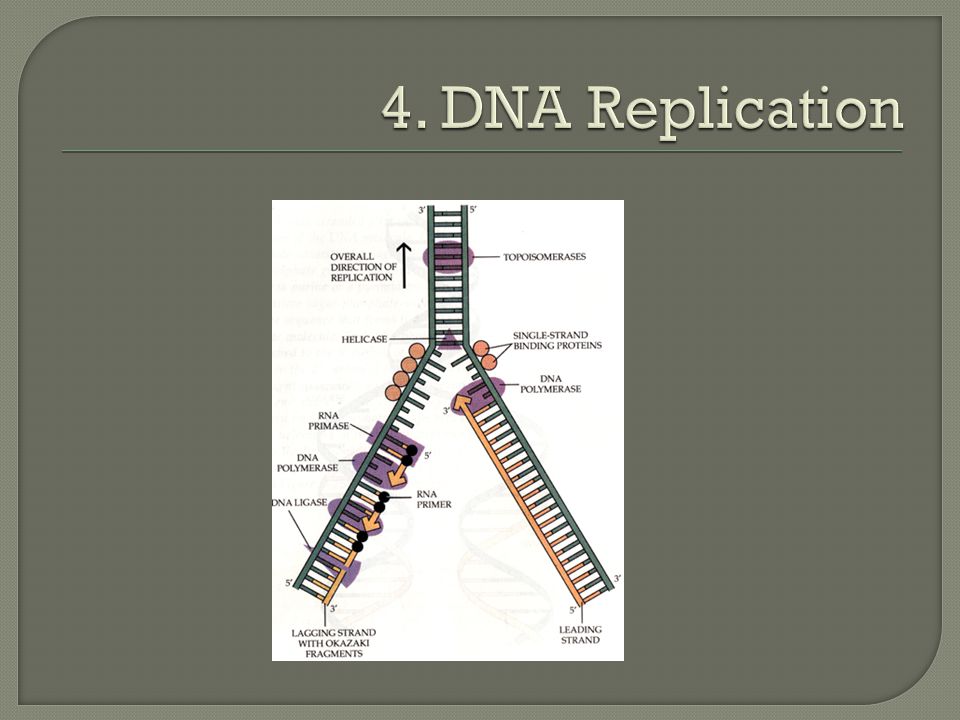
Initiation: DNA replication begins at specific sites on the double helix referred to as origins of replication. An enzyme called helicase binds to DNA and separates the strands.

35
Initiation (continued) Replication proceeds in both directions, creating replication “bubbles” DNA has many origins of replication that can start simultaneously, for time efficiency. Thousands of bubbles can be present, and eventually all the bubbles merge, yielding two completed daughter DNA molecules.

36
Elongation: Replication fork forms: Partial opening of a DNA helix to form two single strands that has a fork appearance. DNA polymerase adds DNA nucleotides in the 5’ 3’ direction

37
Elongation (continued): Leading strand: one of the daughter strands can be synthesized in one continuous fashion working toward the forking point of the parental DNA.
: Leading strand: one of the daughter strands can be synthesized in one continuous fashion working toward the forking point of the parental DNA.")
38
Elongation (continued): Lagging strand: other daughter strand polymerase molecules must work outward from the forking point, It is synthesized in short pieces (called Okazaki fragments) as the fork opens up in a discontinous fashion DNA ligase links (ligates) the pieces together into a single DNA strand.
: Lagging strand: other daughter strand polymerase molecules must work outward from the forking point, It is synthesized in short pieces (called Okazaki fragments) as the fork opens up in a discontinous fashion DNA ligase links (ligates) the pieces together into a single DNA strand.")
39
Termination: At the completion DNA replication, you end up with two identical strands of DNA. DNA ligase and DNA polymerase- proofread the DNA and repair any mistakes Each DNA molecule contains one original parent strand, and one new daughter strand. Semi conservative replication

41
http://www.wiley.com/college/pratt/047 1393878/student/animations/dna_replica tion/index.html

42
Key enzymes: Helicase: unwinds the double helix DNA polymerase- adds DNA nucleotides DNA ligase- joins the ends of DNA segments; DNA repair

43
Process is not only fast but also amazingly accurate Typically, only about one DNA nucleotide per billion is incorrectly paired Ensure that all somatic cells in a multicellular organism carry the same genetic information.

Similar presentations