Download presentation
Presentation is loading. Please wait.

1
14-3-3 proteins, what are they really? 1.A little bit about the family 2.Where they are found 3.Structure 4.Cell Apoptosis 5.Nerve cells 6.Nerve cell degeneration 7.The future of 14-3-3

2
A little bit about the family 30kDa acidic proteins 70% of the proteins is conserved on the amino acid level Each monomer is denoted by a Greek character: β, γ, ε, ζ, η, σ, and τ In vivo they form homo and heterodimers Phosphoryalted or nonphosphorylated?

4
These proteins make up 1% of all the soluble proteins in the brain 14-3-3 ε is expressed in the spermatogonia and 14-3-3 ζ is expressed in primary and secondary spermatocytes. 14-3-3 σ is found in the epithelial cells that are part of the dermal layer of the tongue. Where 14-3-3 proteins are found

5
14-3-3 Structure

6
14-3-3 S136

7
Apoptosis R18 BAD Bcl-2 Bcl-X 14-3-3 Cell Death

8
Nerve cells: Serotonin N- Acetyltransferase Complex AANAT is an enzyme that controls the biological clock. Research has found that 14-3-3 ζ binds with AANAT, which enhances the affinity for AANAT to bind with other substrates. Energy measurements were made after 14-3-3 ζ bound with AANAT and then bound with acetyl- Co-A. In terms of Gibbs free energy, it bound with a ΔG of ~0.8 kcal/mol, as opposed to the large negative values with just AANAT by itself.

9
14-3-3 :Serotonin N-Acetyltransferase Complex

11
Nerve Cells: Parathyroid hormone receptor It has been researched the C-terminal tail of PTHR has been identified as RSGSSSYS this strongly correlates with the 14-3-3 binding motif of RSXpSXP. Two binding sites on PTHR for 14-3-3 It is hypothesized 14-3-3 proteins may act as mediators between steroids and PTHR. This also could have effects on calcium regulation; the exact function of the binding is undetermined.

12
Neurodegenerative disease: Kainic acid (Ka) In rats the build up of Kainic acid (Ka), a neurotoxin, induces the expression of 14-3-3 ζ mRNA. Up regulation of 14-3-3 ζ and p53 The association with p53 for this experiment may suggest that 14-3-3 ζ may be a mediator to this type of cell death. It is hypothesized when 14-3-3 ζ binds to p53 it may change its affinity to bind with DNA sequences.

13
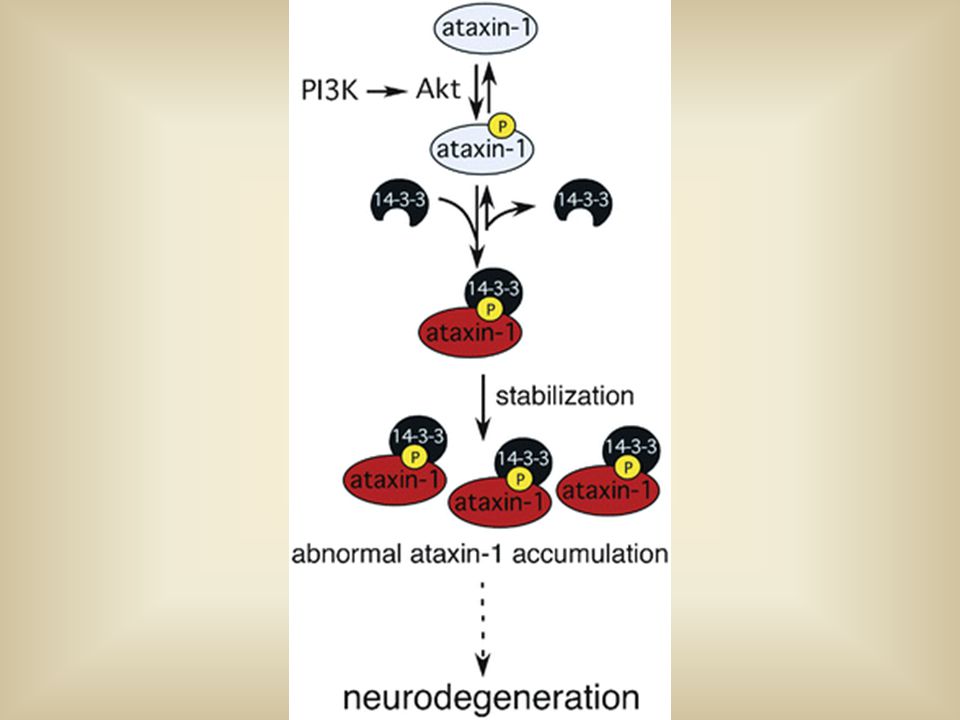
Neurodegenerative disease: Spinocerebellar Ataxia type 1 (SCA1) A neurodegenerative disease that is caused by an unusual long polyglutamine track, 82Q, in the ataxin-1 protein. In vivo Akt phosphorylates S776 on mutant ataxin 1 this allows for 14-3-3 binding. By binding ataxin 1 to 14-3-3 mutant ataxin 1 becomes more toxic to nerve cells because neurdegeneration cannot occur, this causes a build up of mutant ataxin 1and causes degeneration of nerve cells. the longer the polyglutamine track the greater the binding affinity for 14-3-3 isotypes.

15
Neurodegenerative disease: Multiple system atrophy (MSA) This disease causes glial cytoplasmic inclusions (GCI), which are aggregates of α-synulein, found in the nucleus and the presynaptic the cell bodies of oligodendrites and Lewy bodies One major component of the GCIs is α-synuclein, 140 amino acids long. α-synuclein can bind to 14-3-3 by the phosphorylation S129. It is hypothesized that in MSA 14-3-3 may mediate the interaction between α-synuclein and protein kinase C (PKC). This means that 14-3-3 would be a possible cofactor for GCI formation.
. This means that would be a possible cofactor for GCI formation..")
16
J.E Galvin, “Glial Cytoplasmic Inculsions”

17
The future of 14-3-3 In E. tenella 14-3-3 proteins are used in a mannitol cell cycle pathway, this is believed to be a part of the sporulation process. A major enzyme in the pathway is mannitol-1-phosphate dehydrogenase (M1PD) Sm14-3-3—1 Sm14-3-3—1 interacts with a phosphorylated s. mansoni receptor kinase-1domian, which is homologous to the mammalian transforming growth factor β receptor-1 (TGFβ) 26 to 45% effective in preventing in mice infected with S. mansoni
 Sm14-3-3—1 Sm14-3-3—1 interacts with a phosphorylated s. mansoni receptor kinase-1domian, which is homologous to the mammalian transforming growth factor β receptor-1 (TGFβ) 26 to 45% effective in preventing in mice infected with S. mansoni.")
18
For Further Reading: [1] M.B. Yaffe, How do 14-3-3 proteins work? Gatekeeper phosphorylation and the molecular anvil hypothesis. FEBS Lett. 513 (2002), pp. 53– 57. [2] H. Fu, R.R. Subramanian, S.C Masters, 14-3-3 Proteins: Structure, Function, and Regulation. Annu. Rev. Pharmacol. Toxicol 40 (2000), pp. 614-47. [3] T. Obsil, “Structural Biology of 14-3-3 protiens” www.xray.cz/xray/csca/nh/obsil.htm (2001)www.xray.cz/xray/csca/nh/obsil.htm [4] M.B Yaffe, A.E.H. Elia, Phosphoserine/threonine-binding domains. Curr. Opin. Cell Biol. 2 (2001), pp. 131-138 [5] H. Yang, S.C. Masters, H. Wang and H. Fu. Biochim. Biophys. Acta 1547 (2001), pp. 313–319. [6] H. Ibelgaufts “COPE: Apoptosis” www.copewithcytokines.de/cope.cgi?000638 (2003)www.copewithcytokines.de/cope.cgi?000638 [7] R.R. Subramanian, S.C. Masters, H. Zhang and H. Fu. Exp. Cell Res. 271 (2001), pp. 142–151. [8] M. Adachi, Y.B. Zhang, K. Imai, Mutation of BAD with in the BH3 domain impairs its phosphorylation-mediated regulation. FEBS lett. 551 (2003), pp. 147-152. [9] S.C. Masters, H. Fu, 14-3-3 Proteins Mediate an Essential Anti-apoptotic Signal. J. Biol. Chem. 276 (2001), pp. 45193-45200. [10] A.J. Muslin and H. Xing, 14-3-3 proteins: regulation of subcellular localization by molecular interference. Cell. Signal. 19 (2000), pp. 703– 709. [11] K. Toyooka et al., Isolation and the structure of the mouse 14-3-3 η chain gene and the distribution of 14-3-3 η mRNA in the mouse brain. Mole. Brain Res. 100 (2002) pp. 13-20. [12] H. Tazawa, S. Takahashi, J. Zilliacus, Interaction of the parathyroid hormone receptor with the 14-3-3 protien. Biochim. Biophys. Acta 160 (2003) pp. 32-38. [13] T. Obsil, R. Ghirlando, D.C. Klein, S. Gangly, F. Dyda, Crystal Structure of the 14-3-3ζ: Serotonin N-Acetyltransferase Complex: A Role for Scaffolding in Enzyme Regulation. Cell. 105 (2001), pp. 257-267. [14] T. Komori, K. Ishizawa, N. Arai, Immunoexpression of 14-3-3 proteins in glial cytoplasmic inclusions of multiple system atrophy. Acta Neuropath. 106 (2003) pp.66-70. [15] M.P. Brug, S. Goodenough, P. Wilce, Kainic acid induces 14-3-3 ζ expression in distinct regions of rat brain. Brain Res. 956 (2002) pp. 110- 115. [16] X. Monatano, Correspondence: Common amino acid motifs in p53, 14-3-3 and Akt protein families. FEBS Lett. 507 (2001), pp. 237-240. [17] H.K. Chen et al., Interaction of Akt-Phosphorylated Ataxin-1 with 14-3-3 Mediates Neurodegeneration in Spinocerebellar Ataxia Type-1. Cell 113 (2003), pp. 457-468. [18] K.K. Dev, K. Hofele, S. Barbieri, V.L. Buchman, H. Putten, Part II: α-synuclein and its molecular pathophysiological role in neurodegenerative disease. Neuropharm. 45 (2003) pp. 14-44. [19] M.M. Siles-Lucas, B. Gottstein, The 14-3-3 protein: a key molecule in parasites as in other organisms. Tren. Parasite. 12 (2003) pp. 575-581. [20] D. Schectman et al., The 14-3-3 protein as a vaccine candidate against schistosomiasis. Parasite Immunol. 23 (2001), pp. 213-217. [21] J.E Galvin, “Glial Cytoplasmic Inculsions”.http://www.neuro.wustl.edu/people/galvin.html. (2000)http://www.neuro.wustl.edu/people/galvin.html [22] H. Ibelgaufts “COPE: Raf” http://www.copewithcytokines.de/cope.cgi?008043 (2002)http://www.copewithcytokines.de/cope.cgi?008043
![For Further Reading: [1] M.B. Yaffe, How do proteins work.](http://images.slideplayer.com/10/2810723/slides/slide_18.jpg "Gatekeeper phosphorylation and the molecular anvil hypothesis. FEBS Lett. 513 (2002), pp. 53– 57. [2] H. Fu, R.R. Subramanian, S.C Masters, Proteins: Structure, Function, and Regulation. Annu. Rev. Pharmacol. Toxicol 40 (2000), pp [3] T. Obsil, Structural Biology of protiens (2001) [4] M.B Yaffe, A.E.H. Elia, Phosphoserine/threonine-binding domains. Curr. Opin. Cell Biol. 2 (2001), pp [5] H. Yang, S.C. Masters, H. Wang and H. Fu. Biochim. Biophys. Acta 1547 (2001), pp. 313–319. [6] H. Ibelgaufts COPE: Apoptosis (2003) [7] R.R. Subramanian, S.C. Masters, H. Zhang and H. Fu. Exp. Cell Res. 271 (2001), pp. 142–151. [8] M. Adachi, Y.B. Zhang, K. Imai, Mutation of BAD with in the BH3 domain impairs its phosphorylation-mediated regulation. FEBS lett. 551 (2003), pp [9] S.C. Masters, H. Fu, Proteins Mediate an Essential Anti-apoptotic Signal. J. Biol. Chem. 276 (2001), pp [10] A.J. Muslin and H. Xing, proteins: regulation of subcellular localization by molecular interference. Cell. Signal. 19 (2000), pp. 703– 709. [11] K. Toyooka et al., Isolation and the structure of the mouse η chain gene and the distribution of η mRNA in the mouse brain. Mole. Brain Res. 100 (2002) pp [12] H. Tazawa, S. Takahashi, J. Zilliacus, Interaction of the parathyroid hormone receptor with the protien. Biochim. Biophys. Acta 160 (2003) pp [13] T. Obsil, R. Ghirlando, D.C. Klein, S. Gangly, F. Dyda, Crystal Structure of the ζ: Serotonin N-Acetyltransferase Complex: A Role for Scaffolding in Enzyme Regulation. Cell. 105 (2001), pp [14] T. Komori, K. Ishizawa, N. Arai, Immunoexpression of proteins in glial cytoplasmic inclusions of multiple system atrophy. Acta Neuropath. 106 (2003) pp [15] M.P. Brug, S. Goodenough, P. Wilce, Kainic acid induces ζ expression in distinct regions of rat brain. Brain Res. 956 (2002) pp [16] X. Monatano, Correspondence: Common amino acid motifs in p53, and Akt protein families. FEBS Lett. 507 (2001), pp [17] H.K. Chen et al., Interaction of Akt-Phosphorylated Ataxin-1 with Mediates Neurodegeneration in Spinocerebellar Ataxia Type-1. Cell 113 (2003), pp [18] K.K. Dev, K. Hofele, S. Barbieri, V.L. Buchman, H. Putten, Part II: α-synuclein and its molecular pathophysiological role in neurodegenerative disease. Neuropharm. 45 (2003) pp [19] M.M. Siles-Lucas, B. Gottstein, The protein: a key molecule in parasites as in other organisms. Tren. Parasite. 12 (2003) pp [20] D. Schectman et al., The protein as a vaccine candidate against schistosomiasis. Parasite Immunol. 23 (2001), pp [21] J.E Galvin, Glial Cytoplasmic Inculsions . (2000) [22] H. Ibelgaufts COPE: Raf (2002)")
Similar presentations




, or other advanced Molecular/Cell.>")
















