Download presentation
Presentation is loading. Please wait.

1
Berg • Tymoczko • Stryer
Biochemistry Chapter 8: Enzymes: Basic Concepts and Kinetics Copyright © 2007 by W. H. Freeman and Company

2
The enzyme Aequorin catalalyzes the oxidation of this compd in presence of calcium to release CO2 and light

3
Properties of Enzymes Enzymes are protein catalysts. Increase rate of chemical reaction NOT CONSUMED. Show high catalytic efficiency…enzyme catalyzed reactions from 103 to 106 times faster. Highly specific, catalyze only one type of chemical reaction Some have non-protein cofactor needed for enzymatic activity. Cofactors commonly metal ions like Zn2+ and Fe2+, derivatives of vitamins, NAD+ , FAD, and Coenzyme A.

4
Terminology Holoenzyme = enzyme (protein) + cofactor (non protein)
absence of cofactor, no activity Apoenzyme: protein portion of holoenzyme Zymogen (Proenzyme): catalytically inactive enzyme complex (Ex: Chymotrypsinogen, pepsin) Prosthetic group: tightly bound cofactor that does not dissociate from enzyme (Ex: Biotin).
: catalytically inactive enzyme complex (Ex: Chymotrypsinogen, pepsin) Prosthetic group: tightly bound cofactor that does not dissociate from enzyme (Ex: Biotin).")
5
Enzymes Powerful, Highly Specific Catalysts
Accelerate reactions by six orders of magnitude or more. Carbonic anhydrase catalyzes hydration of carbon dioxide One of fastest enzymes known Catalyzed reaction is 107 times as fast as uncatalyzed one

6
Enzymes Powerful, Highly Specific Catalysts
Precise interaction of substrate with enzyme Three dimensional structure of enzyme Specific in terms of Reaction Substrate Catalyzes single chemical reaction or a set of closely related reactions Eg. Proteases: catalyze hydrolysis of peptide bonds. Most proteolytic enzymes also catalyze a different but related reaction in vitro hydrolysis of an ester bond

7
Enzymes Are Powerful and Highly Specific Catalysts
Substrate specificity Proteolytic enzymes differ markedly in their substrate specificity Papain Wide substrate specificity Cleaves any peptide bond Trypsin specific Cleaves only on the C terminus of Lysine and Arginine residues Thrombin Very Specific Catalyzes hydrolysis of Arg-Gly bonds in a particular sequence only

8
Cofactors Catalytic activity of enzymes depends on presence of small molecules, Cofactors Apoenzyme + cofactor = holoenzyme Cofactors Metals Small organic molecules: coenzymes Derived from vitamins Tightly bound termed prosthetic groups Loosely bound like co-substrates

10
Free energy Useful Thermodynamic Function to Understand Enzymes
Key activity in living systems is ability to convert one form of energy into another Photosynthesis Light energy converted into chemical energy Enzymes play vital roles in energy transformation Play fundamental role in photosynthesis and cellular respiration Other enzymes use chemical bond energy of ATP in diverse ways Myosin converts bond energy of ATP into mechanical energy of contracting muscles

11
Free energy Useful Thermodynamic Function to Understand Enzymes
Thermodynamic properties of reaction Free energy difference between reactants and products Determines if reaction occurs or not Energy to initiate conversion of reactants into products Determines rate of reaction Affected by enzymes

12
Bioenergetics Study of energy changes in biochemical reactions
Why some reactions occur while others do not. Bioenergetics deals with energy: Release Storage Use in biological systems Nonbiological reactions utilize heat energy. Biological systems isothermic, use chemical energy.

13
Free energy Useful Thermodynamic Function to Understand Enzymes
Free energy change (ΔG) of chemical process is measure of energy available to do work. ΔG = Gproducts – Greactants If ΔG <0 , forward reaction favored. Products more stable than reactants. As reaction proceeds, energy released that can do work. If ΔG = 0, reactants and products at equilibrium If ΔG >0, reactants at lower energy than products; energy needed for reaction. ΔG is independent of the path of the transformation ΔG for oxidation of glucose the same whether combustion or enzyme catalyzed reactions ΔG provides no information on rate of reaction. Rate depends on free energy of activation, unrelated to ΔG of reaction
 of chemical process is measure of energy available to do work. ΔG = Gproducts – Greactants. If ΔG <0 , forward reaction favored. Products more stable than reactants. As reaction proceeds, energy released that can do work. If ΔG = 0, reactants and products at equilibrium. If ΔG >0, reactants at lower energy than products; energy needed for reaction. ΔG is independent of the path of the transformation. ΔG for oxidation of glucose the same whether combustion or enzyme catalyzed reactions. ΔG provides no information on rate of reaction. Rate depends on free energy of activation, unrelated to ΔG of reaction.")
14
Free energy Useful Thermodynamic Function to Understand Enzymes
Standard Free Energy Change is related to Equilibrium Constant ΔGo’ is directly dependent on the equilibrium constant K’eq. For a reaction: A B C + D ΔG = ΔGo’ + RT. ln [C] [D]/[A] [B] ΔG depends on the nature of the reactants (expressed in the ΔGo’ term) as well as the concentration of the reactant and products At equilibrium, ΔG = 0, and K’eq = [C] [D]/[A] [B] ΔGo’ = - RT.ln K’eq or ΔGo’ = RT log K’eq since R= x 10-3 kJ.mol-1.deg-1 and T = 298K, ΔGo’ = log K’eq or K’eq = 10 –ΔGo’ /5.71 The free energy change of the forward reaction is equal in magnitude but opposite in sign to that of the backward reaction. A B -5 kcal B A +5 kcal
 as well as the concentration of the reactant and products. At equilibrium, ΔG = 0, and K’eq = [C] [D]/[A] [B] ΔGo’ = - RT.ln K’eq or ΔGo’ = RT log K’eq. since R= x 10-3 kJ.mol-1.deg-1 and T = 298K, ΔGo’ = log K’eq or K’eq = 10 –ΔGo’ /5.71. The free energy change of the forward reaction is equal in magnitude but opposite in sign to that of the backward reaction. A B -5 kcal. B A +5 kcal.")
15
Important Facts Criterion for reaction under specified conditions depends on DG not DG0. Knowing concentrations of reactants allows determination! Standard free energy changes additive in any sequence of consecutive reaction: As long as the sum of DGs of individual reactions is negative, the pathway can potentially proceed as written Even if some of the individual component reactions of the pathway have +DGs.

16
Isomerization of DHAP to GAP
At 250C at equilibrium [GAP]/[DHAP] = ΔGo’ = - RT.ln K’eq = X 10-3 X 298 X ln (0.0475) = 7.53 kJ/mol Reaction is endergonic and will not convert spontaneously For initial concentration For [DHAP]/[GAP] = 0.015 ΔG = ΔGo’ + RT.ln K’eq = 7.53kJ/mol kJ/mol = -2.89kJ/mol ΔG indicates it is exergonic and spontaneous Criterion of spontaneity for a reaction is ΔG; not ΔGo’ Reactions not spontaneous based on ΔG can be spontaneous by adjusting concentrations of reactants and products.
 = 7.53 kJ/mol. Reaction is endergonic and will not convert spontaneously. For initial concentration. For [DHAP]/[GAP] = ΔG = ΔGo’ + RT.ln K’eq. = 7.53kJ/mol kJ/mol = -2.89kJ/mol. ΔG indicates it is exergonic and spontaneous. Criterion of spontaneity for a reaction is ΔG; not ΔGo’ Reactions not spontaneous based on ΔG can be spontaneous by. adjusting concentrations of reactants and products.")
17
Additive property of free energy change
Two or more reactions coupled when products of one reaction are reactants of next reaction. Multiple reactions coupled, become a pathway. Pathway must satisfy minimally two criteria: The individual reactions must be specific, yielding only one particular product or set of products. The entire set of reactions in a pathway must be thermodynamically favored An important thermodynamic fact: the overall free energy change for a chemically coupled series of reactions is equal to the sum of the free-energy changes of the individual steps As long as the sum of ΔG’s of the individual reactions is negative the pathway is thermodynamically favorable. A B + C G0’ = + 5 kcal mol-1 B D G0’ = - 8 kcal mol-1 ******************************* A C + D G0’ = - 3 kcal mol-1

18
Enzymes Only alter reaction rate Cannot alter reaction equilibrium
Eg. Conversion of S to P kF = 10-4 s-1 and kR = 10-6 s-1 K = [P]/[S]=kF/kR = 100 Equilibrium concentration of P is 100 times that of substrate, irrespective of enzyme Might take a very long time to achieve this equilibrium in absence of enzyme Equilibrium attainted rapidly in presence of suitable enzyme Thus, enzymes accelerate attainment of equilibrium but do not shift positions.

19
How Enzymes Work Accelerate reactions by facilitating formation of the transition state Chemical reaction of substrate (S) to form product (P) goes through transition state (X‡) S X‡ P Transition state (X‡): Top of the energy hill; Highest free energy, least stable May go to S or P. Not reaction intermediate Bond breakage, bond formation, charge developments take place. Difference in free energy between transition state and substrate called “Gibbs free energy of activation” or “activation energy” G‡ = Gx‡ - Gs V (reaction rate) proportional to G‡ G‡ does not enter G calculation as energy required to generate this is released during product formation. This is how enzymes alter reaction rate without altering G of reaction Enzymes function to lower activation energy or facilitates formation of transition state
 to form product (P) goes through transition state (X‡) S X‡ P. Transition state (X‡): Top of the energy hill; Highest free energy, least stable. May go to S or P. Not reaction intermediate. Bond breakage, bond formation, charge developments take place. Difference in free energy between transition state and substrate called Gibbs free energy of activation or activation energy G‡ = Gx‡ - Gs. V (reaction rate) proportional to G‡ G‡ does not enter G calculation as energy required to generate this is released during product formation. This is how enzymes alter reaction rate without altering G of reaction. Enzymes function to lower activation energy or facilitates formation of transition state.")
20
How Enzymes Work Enzymes form ES complex.
Existence of ES complexes shown in many ways: At constant concentration of enzyme, reaction rate increases with increase in [S] until a Vmax is reached. Maximal velocity suggests formation of ES complex X-ray crystallography Images of substrate analogs bound to active sites Spectroscopic characteristics of many enzymes and substrate change upon formation of ES complex.

21
Active Site Region that binds substrate and possesses catalytic residues that participate in reaction mechanism. Interaction of enzyme and substrate at active site promotes formation of transition state. Region that lowers activation energy of a reaction, thus speeds up reaction

22
Active Site Although structure, specificity, or mode of catalysis of enzymes differ, their active sites share common features Three dimensional cleft, or crevice, formed by groups from different parts of amino acid sequence Small part of total volume of enzyme Enzymes large three-dimensional structures Serve as scaffold to create 3D active site Constitute regulatory sites Interaction with other proteins Substrate channels Unique Microenvironments Water usually excluded unless a reactant Non polar environment enhances binding of substrates as well as catalysis. Polar residues acquire special properties essential for substrate binding and catalysis in microenvironment.

23
Structure of enzyme-substrate complex

24
Active sites include distant residues

25
Active Site Substrates bound to enzymes by multiple weak reversible interactions Electrostatic Hydrogen bonds Directional character results in high degree of specificity Van der Waals forces Becomes significant when numerous substrate atoms come close to many enzyme atoms which requires enzyme and substrate to have complementary shapes.

26
The specificity of binding depends on the precisely defined arrangement of atoms in an active site.
Since enzymes and substrates interact by means of short range forces that require close contact, a substrate must have a matching shape to fit into the site. Emil Fisher’s Lock and Key model (1890) Enzymes are flexible and shape of the active site can be markedly modified by the binding of substrate Daniel Koshland Jr.’s dynamic induced fit model (1958) Active site assumes a shape that is complementary to that of the substrate only after the substrate has been bound.
 Enzymes are flexible and shape of the active site can be markedly modified by the binding of substrate. Daniel Koshland Jr.’s dynamic induced fit model (1958) Active site assumes a shape that is complementary to that of the substrate only after the substrate has been bound.")
27
Induced-Fit Model In absence of substrate, catalytic and substrate binding groups are far from each other. In presence of substrate, conformational change occurs in the enzyme… results in Aligning groups correctly for substrate binding and catalysis Substrate analogs cause some, not all correct conformational changes. Active site has shape complementary to that of substrate only after substrate is bound.

28
Enzyme-Substrate binding energy
Binding energy: Free energy released upon multiple weak interactions between enzyme and substrate. Maximized when right substrate interacts to form interactions Accounts for substrate specificity Maximal energy released when enzyme facilitates formation of transition state Binding energy lowers activation energy Transition state forms S or P depending reaction ΔG

29
Definition of Terms Substrate: substance acted upon by an enzyme. Activity: amount of substrate converted to product by enzyme per unit time (M/min). Specific activity: activity per quantity of protein (M/min/mg protein). International unit: quantity of enzyme needed to transform 1.0 micromole of substrate to product per minute at 30C and optimal pH.
. Specific activity: activity per quantity of protein (M/min/mg protein). International unit: quantity of enzyme needed to transform 1.0 micromole of substrate to product per minute at 30C and optimal pH.")
30
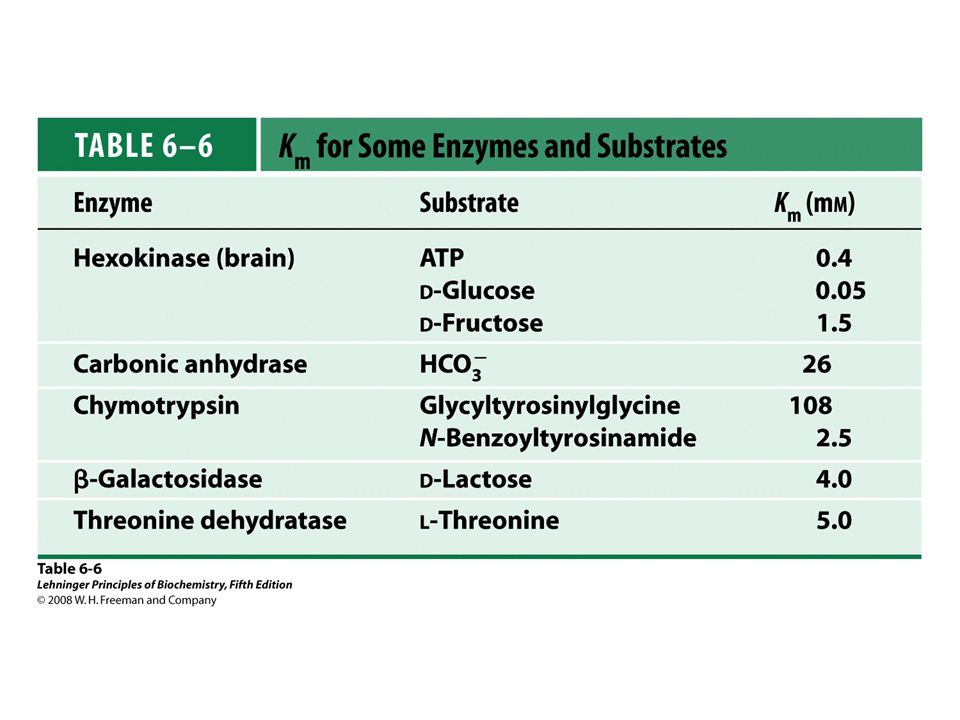
The Michaelis-Menten Equation Describes the Kinetic Properties of Many Enzymes

35
Meaning of KM Two meanings: Measure of strength of ES complex
Substrate concentration when half the active sites filled Dissociation constant of ES complex when k2 is small Measure of strength of ES complex High KM weak binding Low KM strong binding

38
Importance of KM CH3CH2OH + NAD+ CH3CHO + H+ + NADH
Some individuals sensitive to ethanol, exhibit facial flushing, rapid heart rate, after ingesting alcohol. In the liver: Alcohol dehydrogenase CH3CH2OH + NAD+ CH3CHO + H+ + NADH Aldehyde CH3CHO + NAD+ + H2O CH3COO- + NADH + 2H+

39
Most people have 2 forms of acetaldehyde dehydrogenase:
low Km, mitochondrial form high Km, cytosolic form In susceptible persons: the mitochondrial enzyme less active because single amino acid substituted and acetaldehyde processed only by cytosolic enzyme. Cytosolic enzyme has high Km, less acetaldehyde converted into acetate, increased acetaldehyde goes to blood, and symptoms appear.

40
kcat Turnover number: Number of S molecules converted into product by enzyme molecule in a unit time when the enzyme fully saturated with substrate. It is equal to the k3. k3= Vmax/ET

42
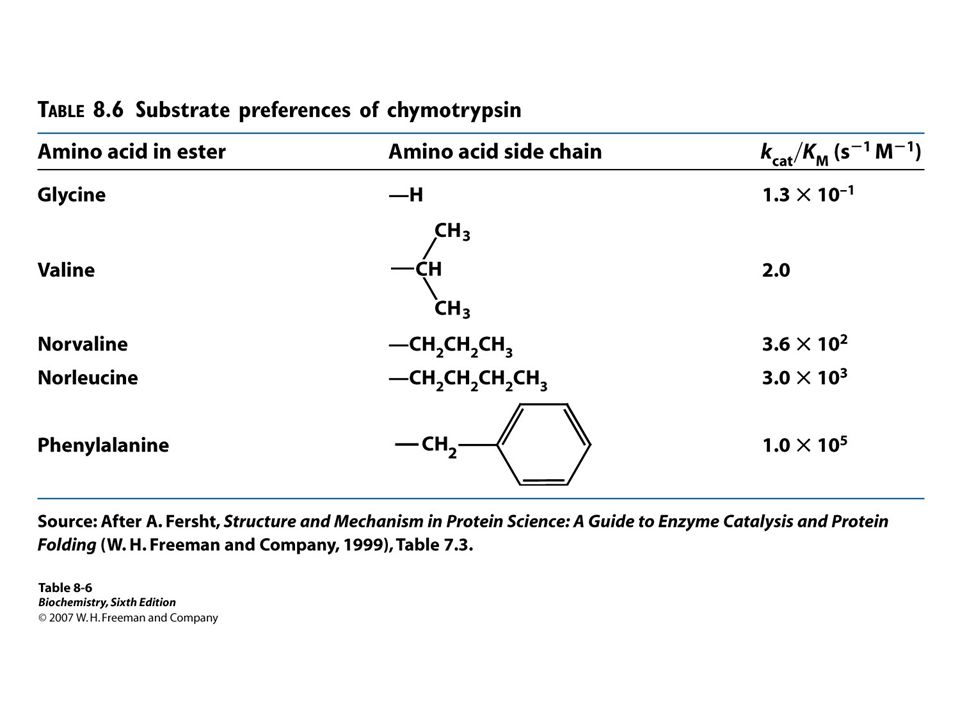
kcat/KM kcat/ KM can be used as a measure of catalytic efficiency.
When the [S]>>>>KM, the rate of catalysis is equal to kcat. But, under physiological conditions, the [S]/KM is equal to (meaning [S]<<<<KM) The enzymatic rate is less than kcat because enzymes active sites are unoccupied. kcat/ KM can be used as a measure of catalytic efficiency. By using kcat/ KM, we can compare an enzyme’s preference for different substrates.
 The enzymatic rate is less than kcat because enzymes active sites are unoccupied. kcat/ KM can be used as a measure of catalytic efficiency. By using kcat/ KM, we can compare an enzyme’s preference for different substrates.")
44
Most biochemical reactions include multiple substrates.
Multiple substrate reactions can be divided into 2 classes: Sequential displacement Double displacement Lactate dehydrogenase enzyme: ordered sequential (NADH must bind first and lactate released first)
")
45
The enzyme (LDH) exists as a ternary complex
Sequential displacement (ordered) The enzyme (LDH) exists as a ternary complex
 The enzyme (LDH) exists as a ternary complex.")
46
Sequential displacement (random)
The order of addition of substrates and release of products is random! Creatine kinase is the enzyme.

47
Cleland notation

48
Double -displacement (ping-pong) reactions
One or more products are released before all substrates bind the enzyme. Enzyme: aspartate amino transferase

49
Ping-pong mechanism

50
Allosteric enzymes Do not obey M-M model!
Multiple subunits, multiple active sites. Display sigmoidal plots. Binding of S to one active site affects properties of other active sites in same enzyme molecule.

51
Allosteric Models:

52
Allosteric inhibitor stabilizes the “T” state whereas an allosteric activator stabilizes the “R” state.

54
Allosteric activators and inhibitors shift the curve in different directions (fES: fraction of sites filled)
")
55
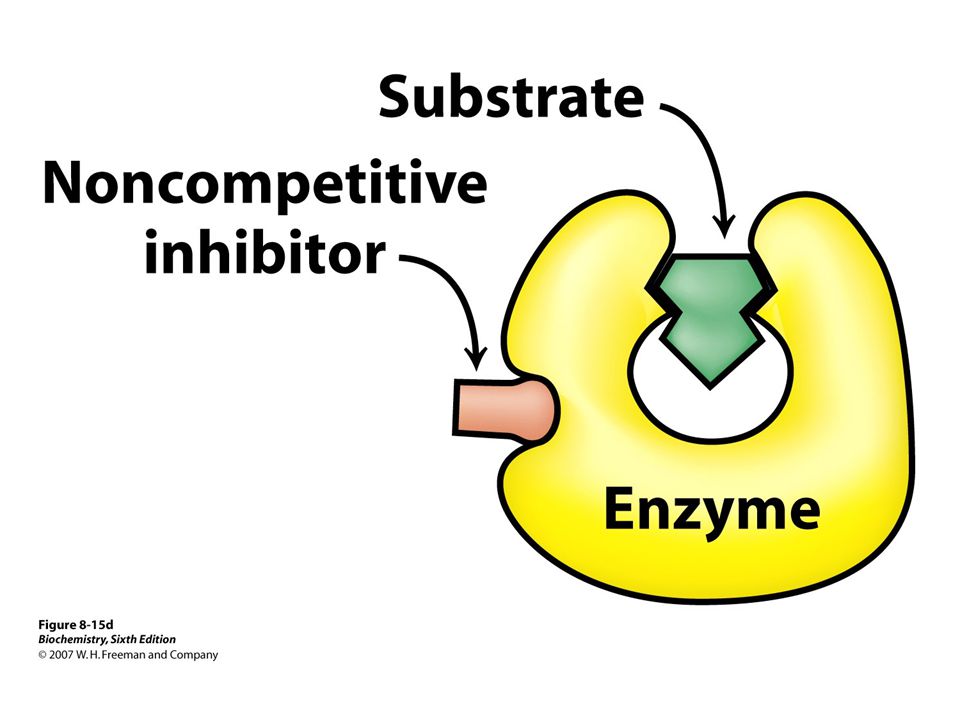
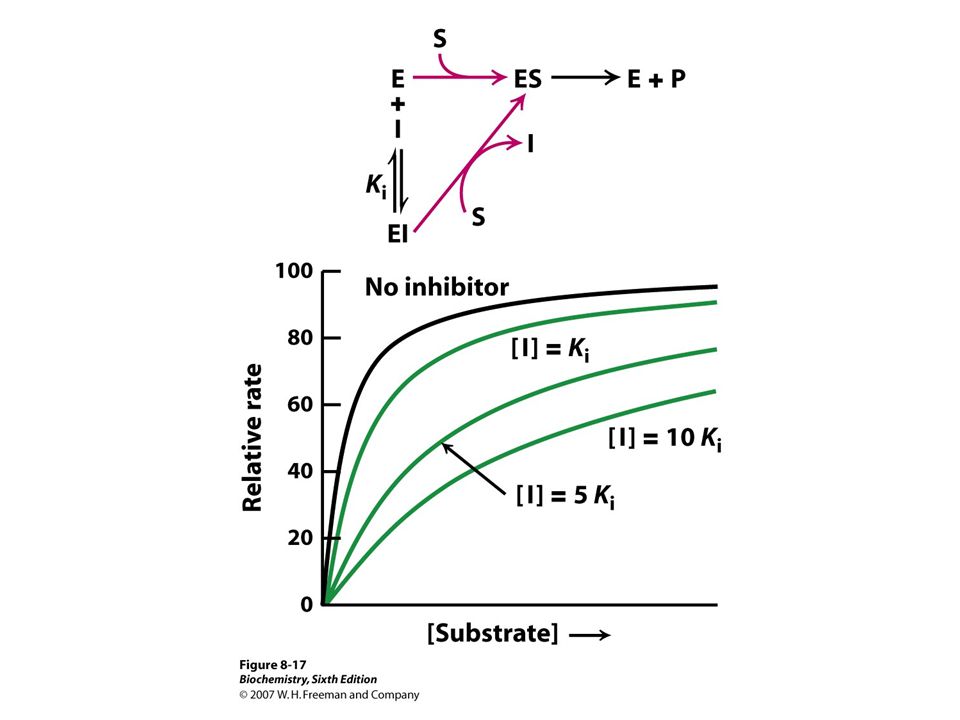
Enzymes inhibited by specific molecules
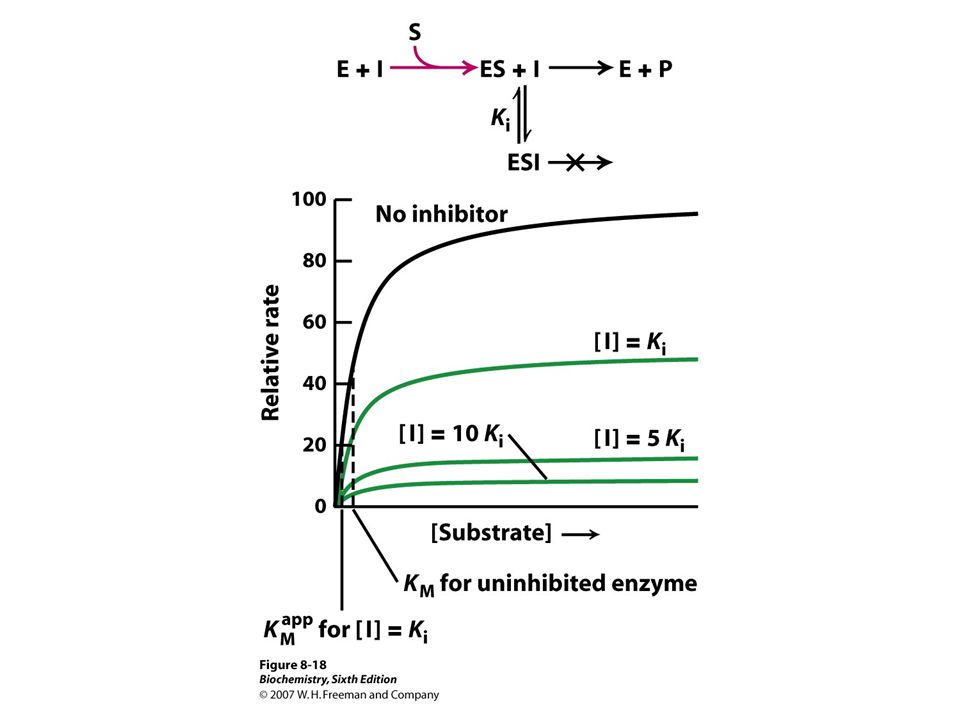
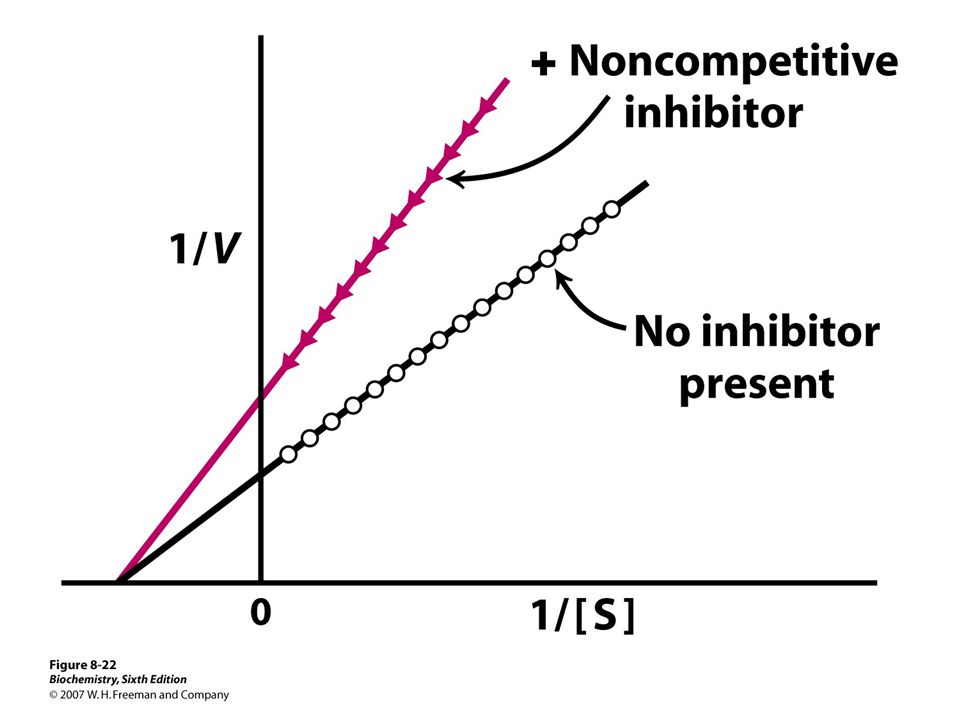
Important because: Major control mechanism Many drugs and toxic agents act this way. 2 kinds of enzyme activity: Reversible competitive noncompetitive Irreversible I dissociates very slowly from the enzyme because I binds to the enzyme covalently. (Ex.: nerve gases)
")
60
Enzyme inhibitors Reversible Irreversible Competitive Noncompetitive
Group specific reagents Substrate analogs Suicide inhibitors

67
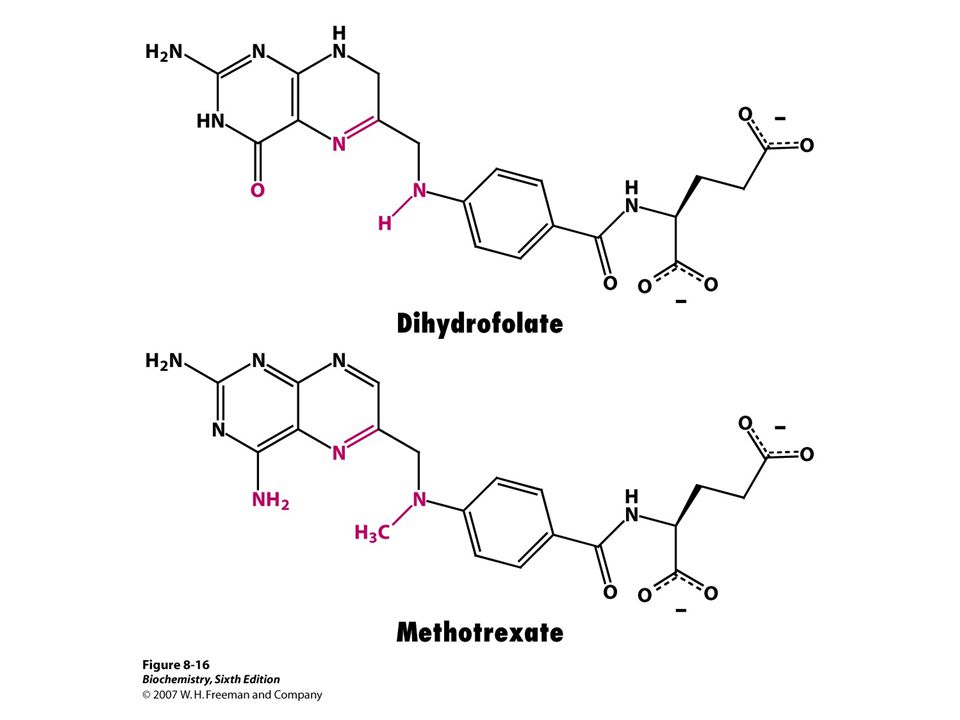
Competitive inhibitors
Resemble substrate, bind to active site of enzyme Methotrexate is structural analog of tetrahydrofolate(coenzyme of dihydrofolate reductase that is involved in purine and prymidine synthesis) Diminish rate of catalysis by reducing proportion of enzyme molecules bound to substrate Competitive inhibitors are used suchmmmmmmmm
 Diminish rate of catalysis by reducing proportion of enzyme molecules bound to substrate. Competitive inhibitors are used suchmmmmmmmm.")
69
Irreversible inhibitors used to map active site
Functional groups important for enzymes activity. How can we be certain about functional groups? X-ray crystallographic of enzyme bound to its substrate Irreversible inhibitors provide alternative approach

70
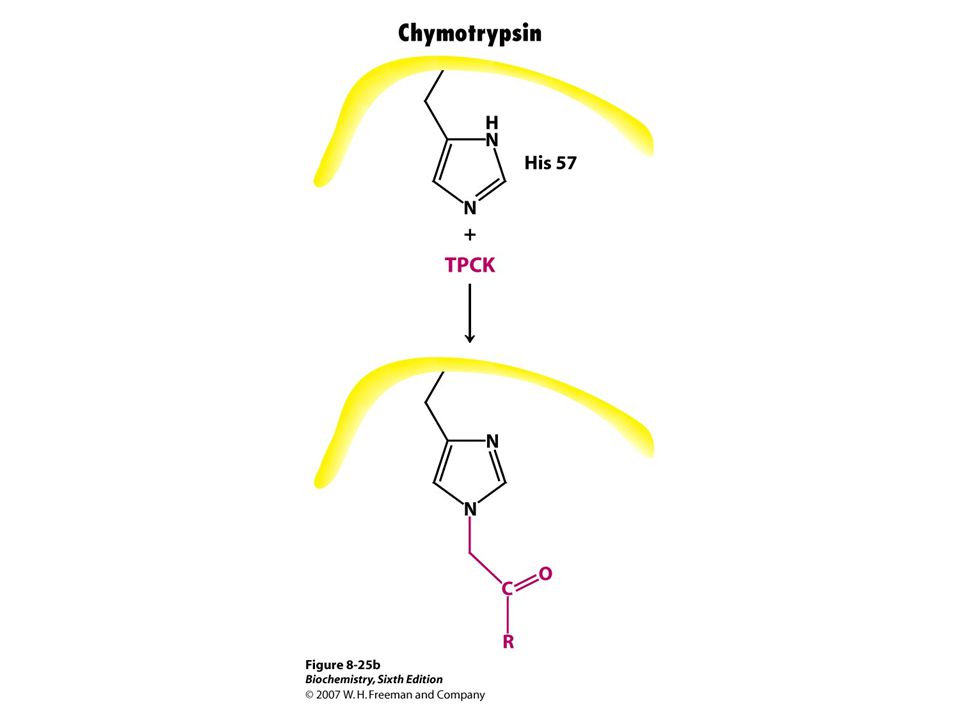
Three groups of irreversible inhibitors
Group specific Reacts with R groups of amino acids DIPF(diisopropylphosphofluoridate) Iodoacetamide Substrate analog Molecules structurally similar to S for enzyme that covalently modifies active site TPCK (tosyl-L-Phe chloromethyl ketone) 3 bromoacetol Suicide inhibitors Enzyme participates in its own irreversible inhibition
 Iodoacetamide. Substrate analog. Molecules structurally similar to S for enzyme that covalently modifies active site. TPCK (tosyl-L-Phe chloromethyl ketone) 3 bromoacetol. Suicide inhibitors. Enzyme participates in its own irreversible inhibition.")
72
Iodoacetamide can inactivate an enzyme by reacting
with a critical cysteine residue.

76
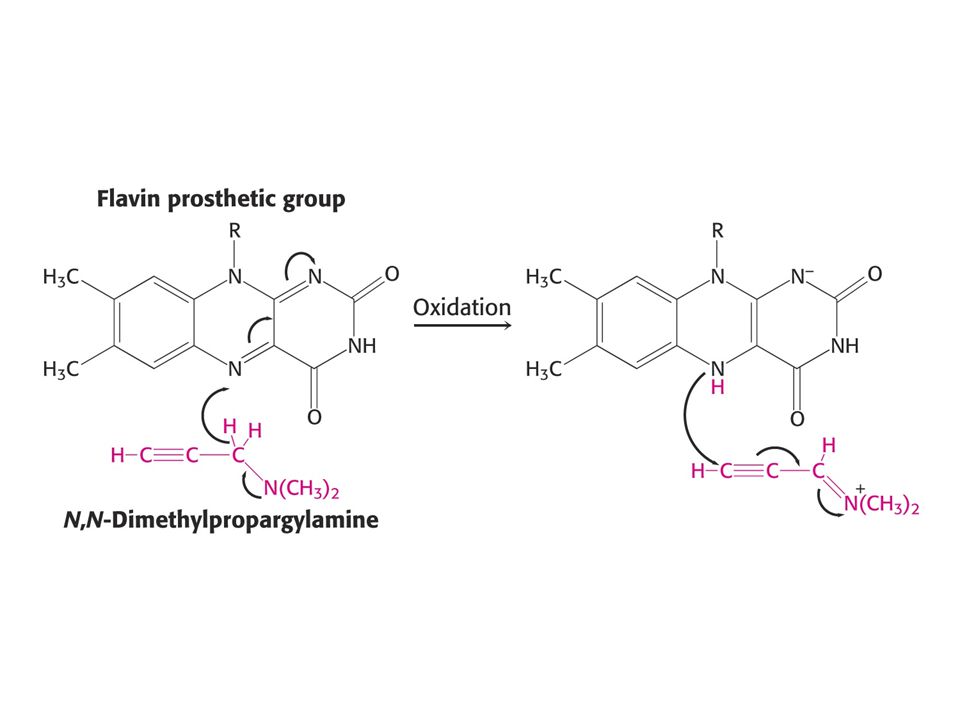
Mechanism-based (suicide) inhibition (MAO inhibitor)
 inhibition (MAO inhibitor)")
80
Suicide inhibitor of MAO

81
Transition state analogs are potent inhibitors of enzymes

84
Penicillin irreversibly inactivates key enzyme in bacterial cell wall synthesis
Cell wall made of macromolecule called “peptidoglycan”, linear polysaccaride chains crosslinked by short peptides. Penicillin blocks last step of cell wall synthesis, i.e., crosslinking of different peptidoglycan strands. Penicillin inhibits crosslinking action of “glycopeptide transpeptidase” enzyme. Penicillin fits in active site of enzyme because looks like D-Ala-D-Ala!

85
Structure of penicilline

87
Peptidoglycan in staphylococcus:
sugar yellow peptide red pentagly blue

88
Formation of crosslinks in peptidoglycan
The terminal amino group of the pentaglycine bridge attacks the peptide bond.

89
Transpeptidation reaction
An acyl-enzyme intermediate is formed in the transpeptidation reaction leading to cross-link formation.

92
Conformations of penicillin and a normal substrate
Penicillin resembles the postulated conformation of the transition state of D-Ala-D-Ala

93
Formation of a penicilloyl-enzyme complex

Similar presentations






























 Involvement.>")




>")







