Download presentation
Presentation is loading. Please wait.

1
Reproduction Mating of odonates
Exam Monday April 11 5pm-7pm TFC Cafeteria Exam Jam Review April 5 Reproduction Mating of odonates Sperm competition: male rivalry of damselflies Rivalry leads to haemocoelic or traumatic insemination in bed bugs Vegetative, budding, polyembryony, parthenogenesis Egg and sperm morphology Hermaphroditism: annelids, limpets, wrasses: timing Pseudoscorpions

2
Terms: asexual vs sexual reproduction
Asexual reproduction: making offspring without syngamy (fusion of sperm and eggsexual) vegetative propagation/budding (e.g., plantlets, stolons, a smaller individual arises on another’s body etc.) binary fission (dividing evenly into two), polyembryony (armadillos) parthenogenesis (aphids) etc. The adaptiveness of sexual reproduction remains a difficult question with a complicated answer. “Current hypotheses suggest that asexual reproduction may have short term benefits when rapid population growth is important [see aphids] or in stable environments, while sexual reproduction offers a net advantage by allowing more rapid generation of genetic diversity, allowing adaptation to changing environments.” (Wikkipedia) dioecious: having male and female organs in separate individuals; monecious: having male and female organs in the same individual.
 vegetative propagation/budding (e.g., plantlets, stolons, a smaller individual arises on another’s body etc.) binary fission (dividing evenly into two), polyembryony (armadillos) parthenogenesis (aphids) etc. The adaptiveness of sexual reproduction remains a difficult question with a complicated answer. Current hypotheses suggest that asexual reproduction may have short term benefits when rapid population growth is important [see aphids] or in stable environments, while sexual reproduction offers a net advantage by allowing more rapid generation of genetic diversity, allowing adaptation to changing environments. (Wikkipedia) dioecious: having male and female organs in separate individuals; monecious: having male and female organs in the same individual.")
4
Order Odonata Suborder Anisoptera, dragonflies (right) Suborder Zygoptera, damselflies (left)
Anisoptera: Adult hind wings wider at base than front wings; wings held horizontal when at rest. Zygoptera: Adult front and hind wings similar in shape and both narrowed at the base; wings usually held together above body when at rest. Univ. of Florida Wikki

6
Larval odonates are aquatic & predatory: larval damselfly (left), larval dragonfly (right)
Gills of damselflies (left) are 3 leaf-like structures at the end of the abdomen; they swim by body undulations, i.e., retrograde body waves and the gills contribute like the caudal fin of a fish. harpercollege.edu harpercollege.edu Recall jetting; rectal gills
 are 3 leaf-like structures at the end of the abdomen; they swim by body undulations, i.e., retrograde body waves and the gills contribute like the caudal fin of a fish. harpercollege.edu. harpercollege.edu. Recall jetting; rectal gills.")
7
“The actual mating act of Odonata is one of the most bizarre performances to be seen anywhere… There is nothing special about the female, the long needle-like abdomen merely terminating in the sexual orifice and the egg-laying apparatus. In the male however, the abdomen terminates in a double set of powerful clasping organs – not unusual in itself until one realizes that these organs are not designed for grasping the genital organs of the female but for grasping her neck. In fact, many female dragonflies have grooves and pits on the back of the head into which these claspers fit. Here then is a dilemma: the claspers (and genital opening) of the male are located at his tail end, but this is applied to the neck of the female rather than to her genital opening. The dilemma is solved by the possession, in these and in no other insects, of a special genital pouch beneath the forward segments of the male abdomen – actually well in front of the middle of the body. Here one finds a jointed penis [aedeagus] and a variety of hooklike appendages associated with a sac [secondary spermatheca] that has no connection with the testes at all. Thus a male must fill these secondary genital organs, which he does by making a loop of his abdomen [think mobility of abdominal tagma], applying the tip briefly to the underside of the base, and permitting sperm to enter the sac at the base of the penis. In damselflies, the [spermatheca] is often filled after the male has grasped the female, but most dragonfly males are “loaded” before they pair with a female.” Howard Evans
 of the male are located at his tail end, but this is applied to the neck of the female rather than to her genital opening. The dilemma is solved by the possession, in these and in no other insects, of a special genital pouch beneath the forward segments of the male abdomen – actually well in front of the middle of the body. Here one finds a jointed penis [aedeagus] and a variety of hooklike appendages associated with a sac [secondary spermatheca] that has no connection with the testes at all. Thus a male must fill these secondary genital organs, which he does by making a loop of his abdomen [think mobility of abdominal tagma], applying the tip briefly to the underside of the base, and permitting sperm to enter the sac at the base of the penis. In damselflies, the [spermatheca] is often filled after the male has grasped the female, but most dragonfly males are loaded before they pair with a female. Howard Evans.")
8
“The act of copulation is initiated [on the wing] by the male following slightly above the female, then seizing her thorax with his legs. At this point, if the female is receptive, she flies slowly and allows herself to be carried. The male then makes a loop of his abdomen and grasps the female in the neck region with his claspers. He then lets go with his legs, straightens his abdomen, and flies ahead, carrying the female in tandem. The female then normally makes a loop downward and forward with her abdomen and applies her genital opening to the genital sac of the male. This is called the ‘wheel position’, since in fact the two form a loop, the female mostly upside-down beneath the male. The pair may remain together for anywhere from three seconds up to an hour or more, but all species that require more than a few seconds copulate while perched.” Howard Evans D. Klimas
![The act of copulation is initiated [on the wing] by the male following slightly above the female, then seizing her thorax with his legs. At this point, if the female is receptive, she flies slowly and allows herself to be carried. The male then makes a loop of his abdomen and grasps the female in the neck region with his claspers. He then lets go with his legs, straightens his abdomen, and flies ahead, carrying the female in tandem. The female then normally makes a loop downward and forward with her abdomen and applies her genital opening to the genital sac of the male. This is called the ‘wheel position’, since in fact the two form a loop, the female mostly upside-down beneath the male. The pair may remain together for anywhere from three seconds up to an hour or more, but all species that require more than a few seconds copulate while perched. Howard Evans](http://slideplayer.com/slide/10419624/35/images/8/The+act+of+copulation+is+initiated+%5Bon+the+wing%5D+by+the+male+following+slightly+above+the+female%2C+then+seizing+her+thorax+with+his+legs.+At+this+point%2C+if+the+female+is+receptive%2C+she+flies+slowly+and+allows+herself+to+be+carried.+The+male+then+makes+a+loop+of+his+abdomen+and+grasps+the+female+in+the+neck+region+with+his+claspers.+He+then+lets+go+with+his+legs%2C+straightens+his+abdomen%2C+and+flies+ahead%2C+carrying+the+female+in+tandem.+The+female+then+normally+makes+a+loop+downward+and+forward+with+her+abdomen+and+applies+her+genital+opening+to+the+genital+sac+of+the+male.+This+is+called+the+%E2%80%98wheel+position%E2%80%99%2C+since+in+fact+the+two+form+a+loop%2C+the+female+mostly+upside-down+beneath+the+male.+The+pair+may+remain+together+for+anywhere+from+three+seconds+up+to+an+hour+or+more%2C+but+all+species+that+require+more+than+a+few+seconds+copulate+while+perched.+Howard+Evans.jpg "D. Klimas.")
9
Wheel Position

10
Opportunity for sperm competition
Male insects transfer sperm to females that store the sperm in a spermatheca, an offshoot of the oviduct. As they eggs mature and advance down the oviduct various glands add protective material (shell). As the egg passes the spermatheca there is an opening in the eggshell through which they are introduced: so females fertilize eggs at the time of oviposition. “When two or more males mate with a female before she oviposits, their sperms do not necessarily have equal chances to fertilize her eggs.” Last in first out: the last male to mate may enjoy precedence over earlier males, and be more successful reproductively. But some insects have evolved structures to ensure their sperm take precedence: they actively remove rival sperm. Calopteryx maculata is a damselfly with beautiful jet-black wings and a green structural colour body. Males of this species (after the wheel position transfer) “defend ovipositing mates from disturbance and take-over attempts by other males”. Waage investigated what was happening in the spermatheca re ‘sperm competition’.
. As the egg passes the spermatheca there is an opening in the eggshell through which they are introduced: so females fertilize eggs at the time of oviposition. When two or more males mate with a female before she oviposits, their sperms do not necessarily have equal chances to fertilize her eggs. Last in first out: the last male to mate may enjoy precedence over earlier males, and be more successful reproductively. But some insects have evolved structures to ensure their sperm take precedence: they actively remove rival sperm. Calopteryx maculata is a damselfly with beautiful jet-black wings and a green structural colour body. Males of this species (after the wheel position transfer) defend ovipositing mates from disturbance and take-over attempts by other males . Waage investigated what was happening in the spermatheca re ‘sperm competition’.")
11
Sperm competition Waage, J.K Dual function of the damselfly penis: sperm removal and transfer. Science 203: Calopteryx maculata “In order to elucidate the mechanism of displacement, I observed copulating pairs directly…When tethered females were used, pairs in copula were induced to land on a net or my hand where the movement of the copulatory organs could be observed with a 10X hand lens. In the five pairs so observed the male made regular undulating movements of his first three abdominal segments [i.e., of secondary aedeagus] during all but the last few seconds of copulation. During this time the sperm vesicle was not in a position to transfer sperm to the female via the penis.”

12
“Scanning electron microscopy revealed several morphological features of the penis that could aid in sperm displacement…flexible head of the penis is probably extensible by internal fluid pressure [hydrostatic] and may aid in scooping …sperm from the bursa copulatrix” “the backward pointing hairs on the ventrolateral surface aid in removing the sperm mass” If insects didn’t store sperm this sort of sperm competition wouldn’t be possible.
![Scanning electron microscopy revealed several morphological features of the penis that could aid in sperm displacement…flexible head of the penis is probably extensible by internal fluid pressure [hydrostatic] and may aid in scooping …sperm from the bursa copulatrix the backward pointing hairs on the ventrolateral surface aid in removing the sperm mass](http://slideplayer.com/slide/10419624/35/images/12/Scanning+electron+microscopy+revealed+several+morphological+features+of+the+penis+that+could+aid+in+sperm+displacement%E2%80%A6flexible+head+of+the+penis+is+probably+extensible+by+internal+fluid+pressure+%5Bhydrostatic%5D+and+may+aid+in+scooping+%E2%80%A6sperm+from+the+bursa+copulatrix+the+backward+pointing+hairs+on+the+ventrolateral+surface+aid+in+removing+the+sperm+mass.jpg "If insects didn’t store sperm this sort of sperm competition wouldn’t be possible.")
13
The lateral horns and spines on the aedeagus of the male let him ‘scrub out’ the branches of the spermatheca (female sperm storage organ) before the male introduces his own sperm. The lower picture shows rival sperm caught in the horn’s spiny hairs.

14
Diagnostic animal genitalia
Animal genitalia are typically quite complex and especially useful in diagnosis of species (Eberhard W.G. 1985). Why should this be so? This is the approach of a book well worth reading: Sexual Selection and Animal Genitalia, Harvard Univ. Press, by William Eberhard. “Since any structure that is a consistently useful taxonomic character at the species level must have evolved both rapidly and divergently (that is, it acquires a new form in each new species), the question can be rephrased as: why do animal genitalia so often evolve both rapidly and divergently?
. Why should this be so This is the approach of a book well worth reading: Sexual Selection and Animal Genitalia, Harvard Univ. Press, by William Eberhard. Since any structure that is a consistently useful taxonomic character at the species level must have evolved both rapidly and divergently (that is, it acquires a new form in each new species), the question can be rephrased as: why do animal genitalia so often evolve both rapidly and divergently")
15
Eberhard W. G. 1985. Sexual selection and Animal Genitalia
Eberhard W.G Sexual selection and Animal Genitalia. Harvard Univ. Press, Cambridge Mass. Rapid and divergent genitalic evolution is common in animals that fertilize internally using an intromittent organ. [And internal fertilization is common terrestrially (though it also occurs in the sea of course) rather than among aquatic organisms.]. And there is a “wider trend toward rapid and divergent evolution of any male structure, genitalic or not, that is specialized to contact females in sexual contexts” (p.181). Why? Several hypotheses brought forward to explain evolution of genitalic diversity are rejected by Eberhard in his book. 1. Hypothesis: ‘lock and key’: elaborate diagnostic genitalia evolve to keep females from being fertilized by males of other species. [mutual structural adjustments don’t occur] 2. Hypothesis: pleiotropism: variations in genitalia are selectively neutral and are the result of selection for other genes. That is, they are effects not functions. [ Ho fails to explain why genitalia are affected like this and not other organs.] 3. Hypothesis: sexual selection by male conflict – intrasexual sexual selection. The hypothesis Eberhard favours: Hypothesis: sexual selection by female choice: “male genitalia function as ‘internal courtship’ devices to increase the likelihood that females will actually use a given male’s sperm to fertilize her eggs rather than those of another male.” and “…the diversity of male genitalic form is the result of runaway evolutionary processes generated by sexual selection by female choice on otherwise arbitrary aspects of male genitalia.” Males with certain abittrary morphologies (forms) of penis do better at leaving offspring than males lacking these features. So the penis evolves to become more and more elaborate just as with the peacock’s tail. (The same argument will explain the evolution of song-specific diversity among acoustic insects.)
 rather than among aquatic organisms.]. And there is a wider trend toward rapid and divergent evolution of any male structure, genitalic or not, that is specialized to contact females in sexual contexts (p.181). Why Several hypotheses brought forward to explain evolution of genitalic diversity are rejected by Eberhard in his book. 1. Hypothesis: ‘lock and key’: elaborate diagnostic genitalia evolve to keep females from being. fertilized by males of other species. [mutual structural adjustments don’t occur] 2. Hypothesis: pleiotropism: variations in genitalia are selectively neutral and are the result of selection for other genes. That is, they are effects not functions. [ Ho fails to explain why genitalia are affected like this and not other organs.] 3. Hypothesis: sexual selection by male conflict – intrasexual sexual selection. The hypothesis Eberhard favours: Hypothesis: sexual selection by female choice: male genitalia function as ‘internal courtship’ devices to increase the likelihood that females will actually use a given male’s sperm to fertilize her eggs rather than those of another male. and …the diversity of male genitalic form is the result of runaway evolutionary processes generated by sexual selection by female choice on otherwise arbitrary aspects of male genitalia. Males with certain abittrary morphologies (forms) of penis do better at leaving offspring than males lacking these features. So the penis evolves to become more and more elaborate just as with the peacock’s tail. (The same argument will explain the evolution of song-specific diversity among acoustic insects.)")
16
Cimicidae: bed bugs, flat, wingless <1 cm, nocturnal ectoparasites sucking blood from birds and bats. Cimex lectularius is the common bedbug that attacks humans; it doesn’t transmit human disease.

17
Male bed bugs practice obligate traumatic insemination: instead of depositing sperm in the genital tract of the female, which is available and functional, males pierce the female’s abdomen and release sperm directly into the haemocoel; sperm migrate to the ovary via the blood. The male intromittent organ is a strongly sclerotized paramere (parameres are terminal structures of insects). The female has evolved new secondary genitalia, an asymmetrically located structure called a ectospermalege (Organ of Berlese), a groove in the right hand margin of the 5th sclerite overlying a pleural membrane
, a groove in the right hand margin of the 5th sclerite overlying a pleural membrane.")
18
Sperm competition: males competing to fertilize the female’s eggs: those that historically put their sperm into the genital tract in the ‘proper’ way, were not as successful as those that put their sperm into the haemocoel. The traumatic inseminators got there first. Greater fitness of a male in competing to fertilize a female’s eggs (as many as possible and excluding so far as possible rival males) is not necessarily better for the female; her fitness may be reduced by increased risk of disease (immune system costs) as well as physical wounding. The evolution of the spermalege may be seen as females reducing these costs.
 is not necessarily better for the female; her fitness may be reduced by increased risk of disease (immune system costs) as well as physical wounding. The evolution of the spermalege may be seen as females reducing these costs..")
19
Pseudoscorpions: spermatophores
Males and females never meet. The males place spermatophores (sperm packages) at various locations in their habitat and the females are attracted and settle over the trigger. Pheromones attract the female to the spermatophore. On land sperm may be packaged in a special spermatophore: a package designed to protect and avoid dessication.
 at various locations in their habitat and the females are attracted and settle over the trigger. Pheromones attract the female to the spermatophore. On land sperm may be packaged in a special. spermatophore: a package designed to protect. and avoid dessication.")
20
Vegetative reproduction
Bryophyllum (Kalanchoe) No meiosis (reduction division). Plantlets on leaves are genetically identical clones.
 No meiosis (reduction division). Plantlets on leaves are genetically identical clones.")
21
Sexual reproduction: the form of a sperm and of an egg reflects their different functions: sperm with a flagellum are adapted for movement, eggs with yolk are specialized to provide the zygote with food. Why do eggs tend to be round. World Science Sexual reproduction involves meiosis to produce haploid gametes which unite to form a new individual/zygote.

22
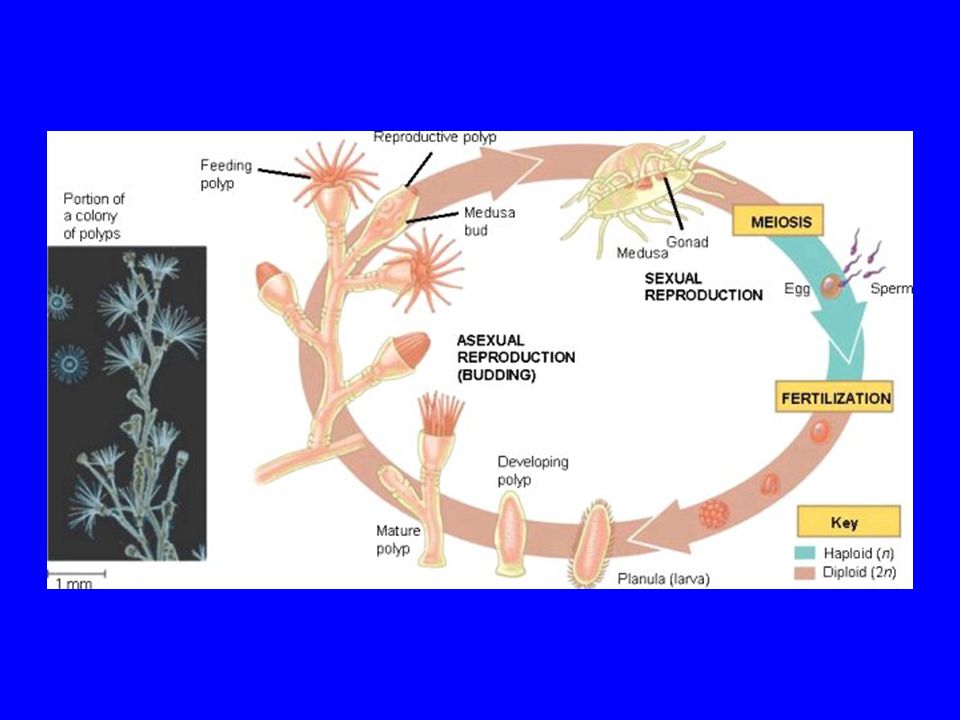
Portuguese Man-of-war: budding
Colonial hydrozoans: budding is the basis of creating the colony: zooids that stay together; but colonial hydrozoa also reproduce sexually with gonophores: zooids specialized for reproduction.

23
Budding and fission in Cnidaria
Coelenterates (Cnidaria) reproduce sexually in the medusa form (most are dioecious); but coelenterates also reproduce by budding and fission. Carolina biological supply
 reproduce sexually in the medusa form (most are dioecious); but coelenterates also reproduce by budding and fission. Carolina biological supply.")
25
parthenogenesis Live birth (viviparity) in aphids; winged female that overwintered as an egg, gives birth to 1st instar females parthenogenetically (without meiosis) in multiple generations, an adaptation to rapidly build up populations to exploit plentiful summertime resources
 in aphids; winged female that overwintered as an egg, gives birth to 1st instar females parthenogenetically (without meiosis) in multiple generations, an adaptation to rapidly build up populations to exploit plentiful summertime resources.")
26
Dasypus, 9-banded armadillo
Fertilized egg divides, creating genetically identical clones; each of these goes on to form an embryo: this is polyembryony.

27
Hermaphrodite: same individual producing both eggs and sperm; extremely common among animals.
Gonads: testes and ovaries, can occur in the same individual or in separate individuals. The latter situation is term dioecious (each gonad ‘in its own house’). If both gonads occur in the same individual that individual is monoecious or hermaphroditic. Incidence of hermaphroditism in organisms is widespread: in many invertebrate animals hermaphroditism is the normal condition: both partners can act simultaneously as "female" and "male: mating involves exchange of sperm. Most snails and slugs are hermaphrodites. Nereis and other polychaetes have separate sexes; but earthworms are hermaphrodites.
. If both gonads occur in the same individual that individual is monoecious or hermaphroditic. Incidence of hermaphroditism in organisms is widespread: in many invertebrate animals hermaphroditism is the normal condition: both partners can act simultaneously as female and male: mating involves exchange of sperm. Most snails and slugs are hermaphrodites. Nereis and other polychaetes have separate sexes; but earthworms are hermaphrodites.")
28
Monoecious and simultaneous hermaphrodites: earthworms are an example of an animal that has both male female gametes functional at the same time. They mate by exchanging sperm within a mucus cocoon (a terrestrial adaptation).
..")
29
Slipper limpets (Phylum Mollusca)
In some animals hermaphroditism is sequential. Slipper limpets (Phylum Mollusca) Crepidula fornicata: is a sequential hermaphrodite: protandry: first male then female. Male gonads mature first and the animal in the early part of its life is a functioning male with embryonic ovaries. It becomes a transitional hermaphrodite and then its male gonads become nonfunctional its ovaries complete development: it ends its life as a female producing eggs. Stacks of slipper limpets have 8 to 12 individuals, all oriented in the same direction; all cling by their muscular foot to the shell of the individual underneath. The basal individual in each stack is usually attached to the shell of dead limpet that is bonded firmly into the substratum. The most basal living individual is a female with mature ovaries that broods (aerates) eggs within the mantle cavity. The youngest and smallest individuals are at the top of the stack and are immatures gradulally becoming adult (functional) males; they arrived atop the stack as dispersing larvae.
 Crepidula fornicata: is a sequential hermaphrodite: protandry: first male then female. Male gonads mature first and the animal in the early part of its life is a functioning male with embryonic ovaries. It becomes a transitional hermaphrodite and then its male gonads become nonfunctional its ovaries complete development: it ends its life as a female producing eggs. Stacks of slipper limpets have 8 to 12 individuals, all oriented in the same direction; all cling by their muscular foot to the shell of the individual underneath. The basal individual in each stack is usually attached to the shell of dead limpet that is bonded firmly into the substratum. The most basal living individual is a female with mature ovaries that broods (aerates) eggs within the mantle cavity. The youngest and smallest individuals are at the top of the stack and are immatures gradulally becoming adult (functional) males; they arrived atop the stack as dispersing larvae.")
30
Slipper limpets contin.
Functional male has a long intromittent organ (penis, aedeagus) that can reach down the stack to fertilize the functional females below. Timing of these sex changes is influenced by the environment. When isolated a small male will develop into a transitional and then a female at a much smaller size than usual. This is apparently a device – an adaptation for most hermaphrodites – of self fertilization in the absence of any partners. A male kept in the presence of several functional females will grow to a much larger than normal size. This developmental effect is governed by release of pheromones into the water: the female releases this chemical which controls the development of the males in the stack above. In effect the mature female regulates the masculinity of the other individuals in the queue. Adaptive value of protandry: protandry separates the role of a female from that of being male, deferring the larger energetic demands of being female (eggs with nutrients) to later in life, when body size is greater, so allowing more accumulation of stored energy. Protandry also reduces brother-sister matings since animals that start life at the same time are not going to be of different sexes at the same time barnacle A long penis is important to a sedentary animal.
 that can reach down the stack to fertilize the functional females below. Timing of these sex changes is influenced by the environment. When isolated a small male will develop into a transitional and then a female at a much smaller size than usual. This is apparently a device – an adaptation for most hermaphrodites – of self fertilization in the absence of any partners. A male kept in the presence of several functional females will grow to a much larger than normal size. This developmental effect is governed by release of pheromones into the water: the female releases this chemical which controls the development of the males in the stack above. In effect the mature female regulates the masculinity of the other individuals in the queue. Adaptive value of protandry: protandry separates the role of a female from that of being male, deferring the larger energetic demands of being female (eggs with nutrients) to later in life, when body size is greater, so allowing more accumulation of stored energy. Protandry also reduces brother-sister matings since animals that start life at the same time are not going to be of different sexes at the same time. barnacle. A long penis is important to. a sedentary animal.")
31
Robertson, D.R. 1972. Social control of sex reversal in a coral-reef fish. Science 177: 1007-1009
Many fishes exhibit protogynous hermaphroditism where the individual develops ovaries first and is a functional female and then subsequently changes into a male with functional testes. Examples: Labridae are common fishes on coral reefs: their common name is wrasse. So too are Scaridae (parrot fishes) and Seranidae (groupers, sea basses). Along with this may go a female biased sex ratio: many more females than males. As an example Labroides dimidiatus blue-headed wrasse. [This fish cleans ectoparasites off other fishes at a ‘cleaner station’.] There are social units: one male with a harem of 3-6 mature females and a number of immatures. The females are yellow with a white underbelly and some black spots. The immatures are yellow (see photo on right).
 and Seranidae (groupers, sea basses). Along with this may go a female biased sex ratio: many more females than males. As an example Labroides dimidiatus blue-headed wrasse. [This fish cleans ectoparasites off other fishes at a ‘cleaner station’.] There are social units: one male with a harem of 3-6 mature females and a number of immatures. The females are yellow with a white underbelly and some black spots. The immatures are yellow (see photo on right).")
32
These fish are territorial on the reef
These fish are territorial on the reef. Removal of the harem-holding male through mischance or old age, permits the alpha female in the dominance hierarchy to initiate rapid developmental changes. She develops male colouration and her female gonads degenerate while her male gonads initiate development to maturity. This happens very quickly, in a matter of a few hours, and so the harem acquires a new male.

Similar presentations