Download presentation
Presentation is loading. Please wait.

1
Figure 3-1: Nutrients for growth of yeast (S. cerevisiae) cells.
 cells.")
4
Figure 3-2: Metabolism in yeast under aerobic and anaerobic conditions.

7
Figure 3. Schematic representation of the key enzymic reactions of pyruvate metabolism in Saccharomyces cerevisiae. Pyc, pyruvate carboxylase; Pdh, pyruvate-dehydrogenase complex; Pdc, pyruvate decarboxylase; Adh, alcohol dehydrogenase; Ald, acetaldehyde dehydrogenase; Acs, acetyl-coenzyme A synthetase.

8
Figure 4. Reoxidation of NADH via the alcoholic fermentation pathway in Saccharomyces cerevisiae. Pdc, pyruvate decarboxylase; Adh, alcohol dyhdrogenase.

9
Crabtree effect In general, the Crabtree effect is described as the occurrence of alcoholic fermentation under aerobic conditions (Crabtree, 1929; de Deken, 1966; Fiechter et al., 1981; Van Dijken & Scheffers, 1986). More specifically, the long-term Crabtree effect and the short-term Crabtree effect can be distinguished (Van Urk, 1989).
..")
10
Long term Crabtree effect The long-term Crabtree effect is defined as aerobic alcoholic fermentation under steady- state conditions at high specific growth rates (Fiechter et al., 1981; Postma et al., 1989). When S. cerevisiae is cultivated in a glucose-limited chemostat the long-term Crabtree effect appears when the dilution rate (and consequently the specific growth rate) exceeds the so-called 'critical specific growth rate' (Figure 1). The long-term Crabtree effect has been explained from a limited respiratory capacity of respiratory sugar metabolism (Fiechter et al., 1981; Petrik et al., 1983; Rieger et al., 1983; Käppeli, 1986). Postma (1989) studied the correlation between the occurrence of the long-term Crabtree effect and the levels of key enzymes in glucose metabolism in glucose-limited chemostat cultures grown at various dilution rates (Postma et al., 1989). An uncoupling effect of respiration (reflected by an enhanced qO2; Figure 1) coincided with the accumulation of some acetic acid in the cultures. Postma et al. proposed that the increased respiration rate might reflect uncoupling by acetate. The accumulation of acetate was explained by these authors from an insufficient amount of acetyl-CoA synthetase at high growth rates (Postma et al., 1989). Similarly, it was proposed that the onset at alcoholic fermentation at even higher dilution rates might be due to a limited capacity of acetaldehyde dehydrogenase, thus necessitating the redirection of the carbon flux via alcohol dehydrogenase (Postma, 1989).
 exceeds the so-called critical specific growth rate (Figure 1). The long-term Crabtree effect has been explained from a limited respiratory capacity of respiratory sugar metabolism (Fiechter et al., 1981; Petrik et al., 1983; Rieger et al., 1983; Käppeli, 1986). Postma (1989) studied the correlation between the occurrence of the long-term Crabtree effect and the levels of key enzymes in glucose metabolism in glucose-limited chemostat cultures grown at various dilution rates (Postma et al., 1989). An uncoupling effect of respiration (reflected by an enhanced qO2; Figure 1) coincided with the accumulation of some acetic acid in the cultures. Postma et al. proposed that the increased respiration rate might reflect uncoupling by acetate. The accumulation of acetate was explained by these authors from an insufficient amount of acetyl-CoA synthetase at high growth rates (Postma et al., 1989). Similarly, it was proposed that the onset at alcoholic fermentation at even higher dilution rates might be due to a limited capacity of acetaldehyde dehydrogenase, thus necessitating the redirection of the carbon flux via alcohol dehydrogenase (Postma, 1989)..")
11
Short-term Crabtree effect The definition of the short-term Crabtree effect is the instantaneous aerobic alcoholic fermentation upon addition of excess sugar to sugar-limited and/or non-fermenting cultures (Rieger et al., 1983; Verduyn et al., 1984; Van Urk, 1989; Figure 2). This effect was explained in terms of an 'overflow' in metabolism. When a culture of S. cerevisiae is exposed to excess sugar the glucose consumption rate increases until the respiratory capacity becomes limiting (Petrik et al.,1983; Käppeli and Sonnleitner, 1986). Van Urk (1989) studied the short-term Crabtree effect and compared some Crabtree-positive with Crabtree-negative yeasts. Several differences were revealed between Crabtree-positive and Crabtree-negative yeasts, including different kinetics of glucose uptake, rates of glycolysis and glycogen formation and the levels of a number of key enzymes involved in pyruvate metabolism. Most strikingly, ethanol production rates showed a clear positive correlation with the level of pyruvate decarboxylase in Crabtree-positive yeasts. All Crabtree-positive yeasts contained higher levels of this fermentative key enzyme than the Crabtree negative yeasts. Furthermore, high levels of acetaldehyde dehydrogenase and acetyl-CoA synthetase were observed in the Crabtree-negative yeasts. These may 'pull' the carbon flux towards respiration, despite the presence of some Pyruvatede carboxylase.
. Van Urk (1989) studied the short-term Crabtree effect and compared some Crabtree-positive with Crabtree-negative yeasts. Several differences were revealed between Crabtree-positive and Crabtree-negative yeasts, including different kinetics of glucose uptake, rates of glycolysis and glycogen formation and the levels of a number of key enzymes involved in pyruvate metabolism. Most strikingly, ethanol production rates showed a clear positive correlation with the level of pyruvate decarboxylase in Crabtree-positive yeasts. All Crabtree-positive yeasts contained higher levels of this fermentative key enzyme than the Crabtree negative yeasts. Furthermore, high levels of acetaldehyde dehydrogenase and acetyl-CoA synthetase were observed in the Crabtree-negative yeasts. These may pull the carbon flux towards respiration, despite the presence of some Pyruvatede carboxylase..")
12
Figure 2. Example of the short-term Crabtree effect in Saccharomyces cerevisiae. Production of ethanol and acetate after a pulse of glucose (50 mM) to an aerobic glucose-limited chemostat culture (Van Urk et al., 1988).
 to an aerobic glucose-limited chemostat culture (Van Urk et al., 1988)..")
14
Pasteur effect The definition of the Pasteur effect is the inhibition of sugar consumption by aerobiosis (Lagunas, 1986).This physiological behaviour or effect was named after Pasteur because of his observations in 1861 on experiments with cultivated brewers' yeast in the presence and absence of air. In her excellent review on the Pasteur effect, Lagunas (1986) identified e this observation as an artefact due to the absence in Pasteur's experiments of the extra nutrients (unsatured fatty acids and sterols) required for growth of S. cerevisiae under anaerobic conditions. In S. cerevisiae, the Pasteur effect is only observed under special experimental conditions, notably at very low dilution rates in the chemostat and in resting-cell suspensions. In sugar-limited chemostat cultures the growth rate and the glucose uptake rate is low due to a low sugar concentration; in resting cells the glucose uptake rate is low due to an inactivation of the sugar transport system. The mechanism which underlies this phenomenon is probably the higher affinity for one of the intermediates pyruvate, acetaldehyde and NADH of the respiratory system over the fermentative route (Lagunas et al., 1982; Lagunas, 1986).
 identified e this observation as an artefact due to the absence in Pasteur s experiments of the extra nutrients (unsatured fatty acids and sterols) required for growth of S. cerevisiae under anaerobic conditions. In S. cerevisiae, the Pasteur effect is only observed under special experimental conditions, notably at very low dilution rates in the chemostat and in resting-cell suspensions. In sugar-limited chemostat cultures the growth rate and the glucose uptake rate is low due to a low sugar concentration; in resting cells the glucose uptake rate is low due to an inactivation of the sugar transport system. The mechanism which underlies this phenomenon is probably the higher affinity for one of the intermediates pyruvate, acetaldehyde and NADH of the respiratory system over the fermentative route (Lagunas et al., 1982; Lagunas, 1986)..")
15
Kluyver effect The Kluyver effect has been described as follows: '.. certain yeasts can utilize particular disaccharides aerobically, but not anaerobically, although these yeast's can use one or more of the component hexoses anaerobically.' (Sims & Barnett, 1978). This phenomenon is widespread under facultatively fermentative yeasts. It has been reported in more than 100 yeast species (Sims and Barnett, 1978; Weusthuis, 1994). S. cerevisiae does not exhibit this effect. The yeast Candida utilis does show the Kluyver effect: it is able to ferment glucose under oxygen-limited conditions but it cannot ferment maltose under these conditions. C. utilis is, however, able to grow rapidly on maltose under aerobic conditions by respiring this disaccharide. Yeasts that exhibit the Kluyver effect for a particular sugar are able to ferment glucose, suggesting that the Kluyver effect must be caused in a difference between the metabolism of this sugar and that of glucose. The exact mechanism underlying this phenomenon has not yet been resolved. However there are some plausible explanations like a lowered rate of sugar transport (i.e. a transport limitation) or control of the synthesis of the sugar transporter (Barnett and Sims, 1982; Barnett, 1992, Weusthuis et al., 1994; Kaliterna et al., 1995). Furthermore some yeasts have a reduced activity of pyruvate decarboxylase during growth on non-glucose sugars, with may contribute to a lower flux through glycolysis (Sims & Barnett, 1991). Weusthuis et al. (1994) proposed feedback inhibition of disaccharide utilisation by ethanol as a possible cause of the Kluyver effect. They have shown that, indeed, added ethanol suppresses the utilisation of maltose by C. utilis (Weusthuis et al.,1994).
. This phenomenon is widespread under facultatively fermentative yeasts. It has been reported in more than 100 yeast species (Sims and Barnett, 1978; Weusthuis, 1994). S. cerevisiae does not exhibit this effect. The yeast Candida utilis does show the Kluyver effect: it is able to ferment glucose under oxygen-limited conditions but it cannot ferment maltose under these conditions. C. utilis is, however, able to grow rapidly on maltose under aerobic conditions by respiring this disaccharide. Yeasts that exhibit the Kluyver effect for a particular sugar are able to ferment glucose, suggesting that the Kluyver effect must be caused in a difference between the metabolism of this sugar and that of glucose. The exact mechanism underlying this phenomenon has not yet been resolved. However there are some plausible explanations like a lowered rate of sugar transport (i.e. a transport limitation) or control of the synthesis of the sugar transporter (Barnett and Sims, 1982; Barnett, 1992, Weusthuis et al., 1994; Kaliterna et al., 1995). Furthermore some yeasts have a reduced activity of pyruvate decarboxylase during growth on non-glucose sugars, with may contribute to a lower flux through glycolysis (Sims & Barnett, 1991). Weusthuis et al. (1994) proposed feedback inhibition of disaccharide utilisation by ethanol as a possible cause of the Kluyver effect. They have shown that, indeed, added ethanol suppresses the utilisation of maltose by C. utilis (Weusthuis et al.,1994)..")
16
Figure 3-3: Sugar metabolism in different yeasts.

17
Figure 3-5: Use of unusual nutrients in yeasts.

18
Figure 3-6: Fatty acid utilization.

19
Peroxisomes are organelles containing a single membrane and no DNA The organelles are involved in a variety of metabolic activities oxidation of fatty acids and lipids purines, amino acids biosynthesis of bile acids and cholesterol removal of peroxide The composition of peroxisomes is highly variable: Pulse-chase studies have shown that peroxisomal proteins are incorporated into the peroxisome after (20 min) synthesis in the cytosol
 synthesis in the cytosol")
21
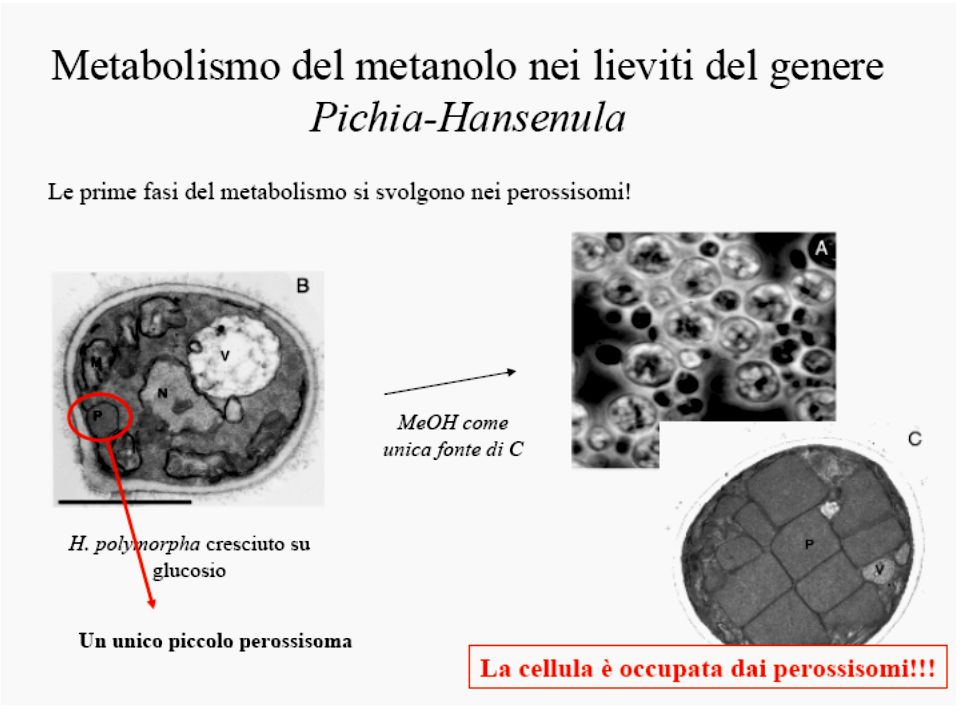
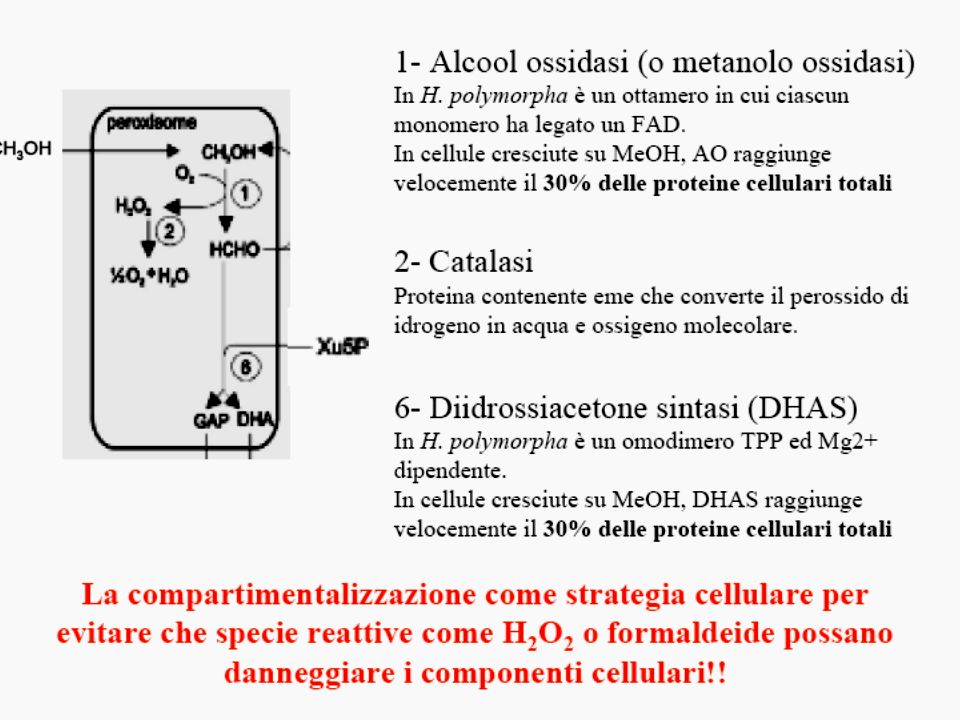
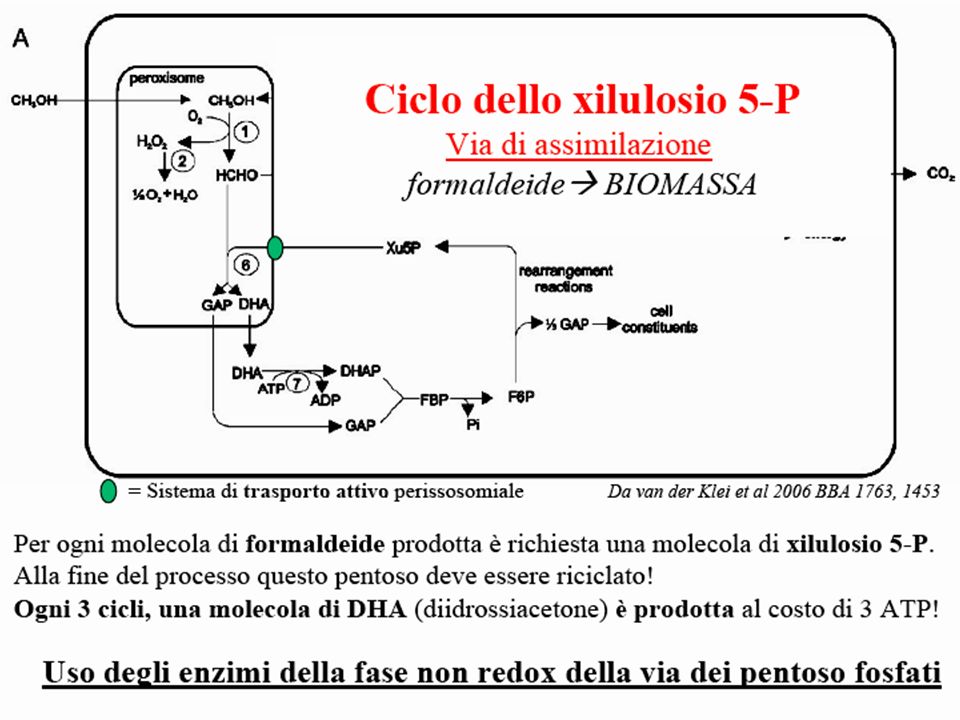
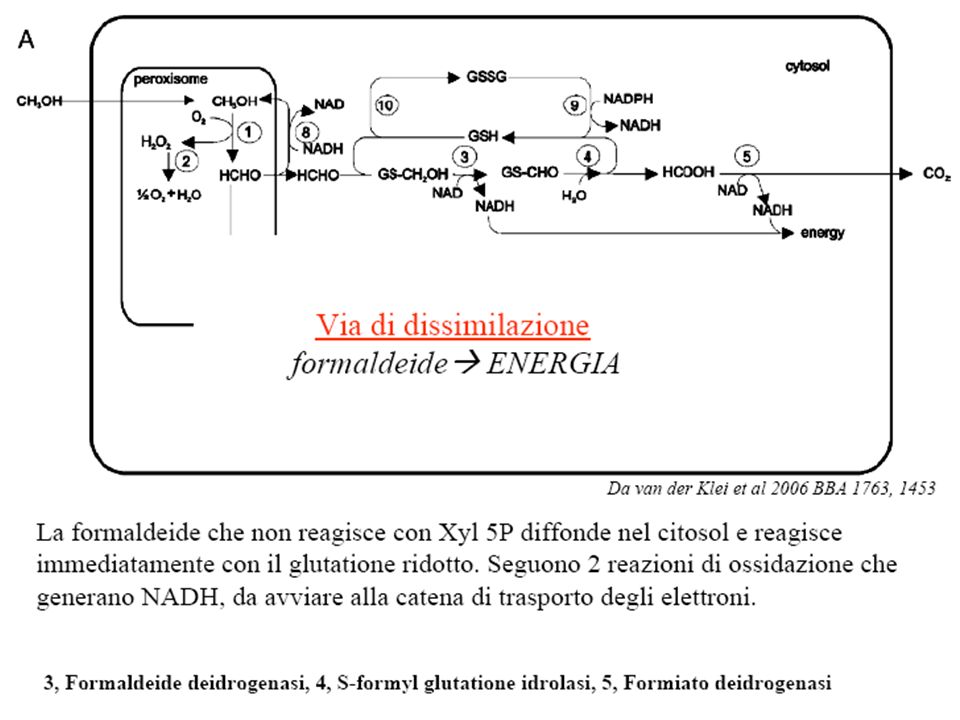
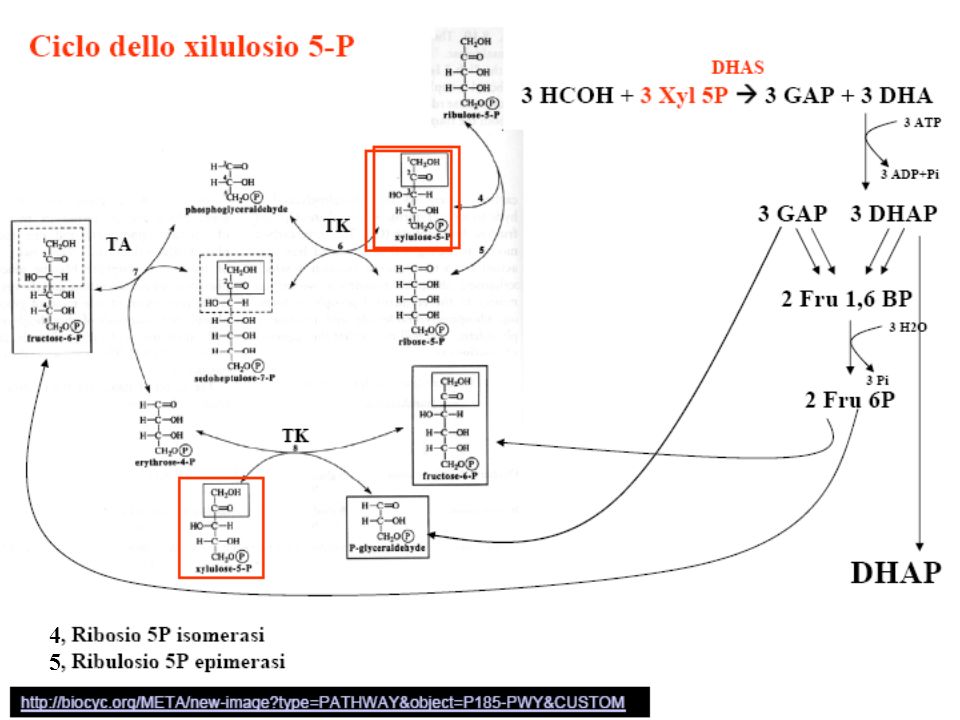
Schematic representation of methanol metabolism in H. polymorpha Schematic representation of the compartmentalization and the function of peroxisomes in methanol metabolism in H. polymorpha WT cells. 1—alcohol oxidase, 2—catalase, 3—formaldehyde dehydrogenase, 4—S-formyl glutathione hydrolase, 5—formate dehydrogenase, 6—dihydroxyacetone synthase, 7—dihydroxyacetone kinase, 8—formaldehyde reductase, 9—glutathione reductase, 10— oxidation of glutathione. Da van der Klei et al 2006 BBA 1763, 1453

27
4 5 4 5
Similar presentations