Download presentation
Presentation is loading. Please wait.

1
Chapter 18: Regulation of Gene Expression
The functions of the three parts of an operon. The role f repressor genes in operons. The impact of DNA methylation and histone acetylation on gene expression. The role of oncogenes, protooncogenes, and tumor supressor genes in cancer.

2
The Genetics of Viruses and Bacteria
Chapter 18 The Genetics of Viruses and Bacteria

3
Viruses They are extremely small--20nm in diameter.
Some are DNA viruses, others are RNA viruses. The DNA or RNA can be single stranded, double stranded, linear or circular. Small viruses have 4 genes, large ones several hundred.

4
Viruses Viruses come in many different shapes and sizes.
Regardless of the size, protein coat that encapsulates their genome is called a capsid. Capsides are built from protein subunits called capsomeres.

5
Viruses Viruses only reproduce inside of cells.
They lack all cellular machinery to carry out any function such as reproduction or metabolism. They only thing they can do is get their genes inside of cells and take them over. Viruses have a host range. Some are very specific, others are non-specific.

6
Viruses Most viruses have proteins on the outside that interact with specific receptor molecules on the surface of cells. This “lock-and-key” system is what limits the specificity of a virus to certain cell types. Once the virus attaches to the cell, it injects its DNA and takes over the control of the cell. It now uses the cell’s machinery to build new virus particles.

7
Viruses It sounds strange, but once the viral nucleic acids and capsomeres are formed, many viruses spontaneously assemble and hundreds of thousands of viral particles leave the cell, usually destroying it. This viral destruction of cells is often the cause of the viral related symptoms we feel when we are infected.

8
Virus Reproduction Phages are the best understood of all viruses.
Viruses reproduce using one of two mechanisms: 1. The lytic cycle. 2. The lysogenic cycle.

9
1. The Lytic Cycle When a phage reproduces and ultimately kills the host cell, it is said to use the lytic cycle. The lytic cycle is the last stage of infection, lysing the cell and releasing phages. Phages that reproduce using only the lytic cycle are said to be virulent phages.

10
1. The Lytic Cycle There are 5 main steps to the lytic cycle of a common phage--T4. 1. Attachment. 2. Injection of genetic material and control of the cell. 3. Synthesis of viral genomes and proteins. 4. Assembly of viruses. 5. Release of phages. The entire cycle lasts about min.

11
A Game of Cat and Mouse The main reason why viruses haven’t wiped out all bacteria and other cells is because surface receptors on cells often mutate preventing attachment of viruses. On the other hand, viruses often mutate enabling them to adapt and attach to the surfaces of cells displaying these new receptors.

12
A Game of Cat and Mouse Another defense many cells have is the ability to detect foreign genetic material and cut it up with degradative enzymes (restriction endonucleases). Some phage mutants produce nucleic acid that is not recognizable by restriction enzymes and can thus take over a cell’s function.
. Some phage mutants produce nucleic acid that is not recognizable by restriction enzymes and can thus take over a cell’s function.")
13
2. The Lysogenic Cycle The lysogenic cycle is yet another reason why viruses haven’t taken over all cells. In this cycle, the phage genome is replicated without destroying the cell. Some phages are called temperate phages because they use both the lysogenic and lytic modes of reproduction. phage is an example.

14
Phage Lifecycle The phage lifecycle is as follows:
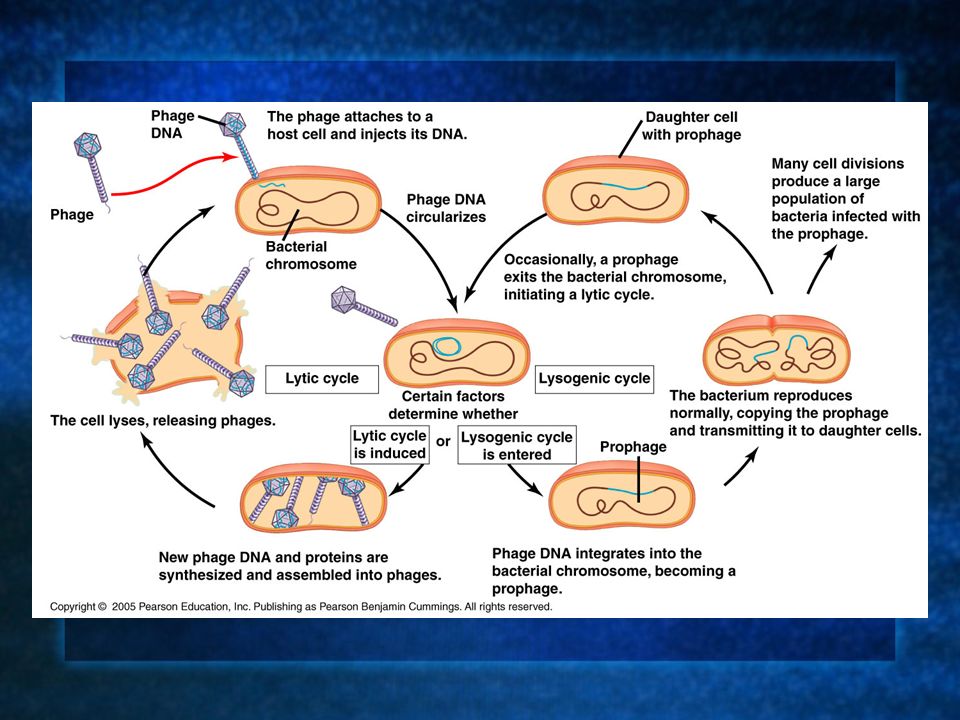
1. Attachment. 2. Injection of DNA into cells. 3. Phage DNA “circulizes.” Environmental factors determine the lytic or lysogenic cycles. Lytic cycle: 4. Phage DNA and proteins are made. 5. Cell lyses releasing phages.

15
Phage Lifecycle The phage lifecycle is as follows:
1. Attachment. 2. Injection of DNA into cells. 3. Phage DNA “circulizes.” Environmental factors determine the lytic or lysogenic cycles. Lysogenic cycle: 4. Phage DNA integrates with bacterial DNA. 5. Bacteria reproduce normally transmitting all DNA to both cells, crossing over occurs. 6. Many cell divisions produce many cells with phage DNA and altered phenotypes.

17
Lytic or Lysogenic The cycle by which a temperate phage reproduces is largely due to environmental signals. A temperate phage can use the lysogenic cycle to incorporate its genome into a bacterium’s and many copies of the phage DNA can be made as the bacterium divides. When something in the environment triggers a switchover to the lytic cycle, the phage then destroys the cell(s).
.")
18
The Ways in Which Viruses Infect Cells
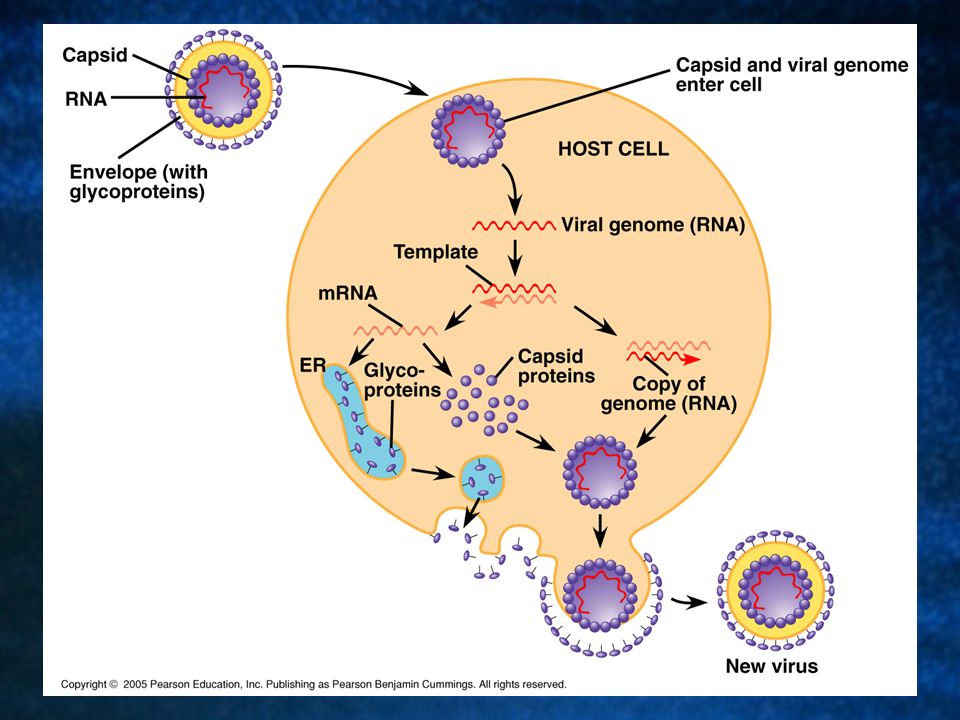
One of the key variables is whether the genetic material is DNA or RNA, and whether or not it is single or double stranded. Many bacteriophages are not RNA viruses and lack membranous envelopes. Most animal viruses with RNA genomes have envelopes.

19
Animal Viruses with RNA Genomes and Viral Envelopes
Animal viruses with viral envelopes use them to enter host cells. The envelopes contain viral glycoproteins that attach to the surface of host cells. When the virus enters the cell, takes it over and begins synthesizing new virus particles. It uses the cellular enzymes in the ER to synthesize glycoproteins to form a new envelope.

20
Animal Viruses with RNA Genomes and Viral Envelopes
In a process similar to exocytosis, the new viruses wrap themselves in the membrane derived from the host’s plasma membrane and bud off from the cell. This reproductive cycle doesn’t necessarily kill the host cell like the lytic cycle.

22
Other Forms of Viral Envelopes
Some viruses reproduce within the nucleus of the cell and don’t use the plasma membrane as an envelope--instead they are derived from the nuclear membrane of the cell. Many of these viruses have double stranded DNA and use a combination of host and viral enzymes to replicate and transcribe their genomes.

23
HIV-A Retrovirus Retroviruses are RNA viruses which reproduce via a DNA intermediate. The actual flow of information is reversed; it goes RNA to DNA back to RNA.

24
Bacteria Most bacteria have a single, circular, double stranded DNA molecule as their chromosome. In contrast to eukaryotic chromosomes, bacterial chromosomes are associated with a relatively small amount of protein. These chromosomes exist in the nucleoid region.

25
Bacteria Bacteria are also often associated with small pieces of DNA called plasmids. Plasmids usually contain DNA fragments that are of benefit to the cell. Bacteria divide via binary fission after replicating their DNA. Recall that the DNA replication begins at the origin and moves around until finished.

27
Bacterial Reproduction
Mutations arise in bacteria at a rate of about 1:10,000,000. This doesn’t seem like much, but considering how prolific they are, this is a high rate. Example: In the human gut, 20 billion new E. coli cells arise each day. With a 1:10 million mutation rate, this produces about 2000 new mutants that increase the genetic diversity of the population.

28
Mutations Mutations are a major source of genetic variation in quickly reproducing organisms. In slowly reproducing populations, genetic recombination (crossing over) is a source of new variation. Bacteria also have a means by which recombination occurs and produces new variation.
 is a source of new variation. Bacteria also have a means by which recombination occurs and produces new variation.")
29
Evidence for Recombination in E. coli
Researchers used two mutant strains of E. coli. One could synthesize tryptophan and not arginine. The other could synthesize arginine and not tryptophan. Samples from the two strains were mixed together to see if they could grow on minimal medium.

31
Evidence for Recombination in E. coli
If recombination occurs, some cells from the mixed culture should grow on minimal medium, whereas they should not from the original mutant strain. Cell colonies grew on minimal medium after being incubated together. Somehow the cells, when incubated together, exchanged genetic material and gave rise to individuals that could survive on minimal medium.

33
3 Mechanisms of Genetic Recombination in Bacteria
1. Transformation. 2. Transduction. 3. Conjugation.

34
1. Transformation Transformation is the alteration of the genotype and phenotype of a cell by the uptake of naked DNA from the surroundings. Recall the Griffith experiment with mice and pneumonia. This was transformation. The mixture of heat killed smooth and the living rough produced recombinant cells that became virulent. They were recombinant because they contained genes from two different cells.

36
2. Transduction Transduction is another means by which bacterial cells can undergo recombination. This process employs phages that carry genes from one cell to another. It often results from irregularities in the phage reproductive cycle.

37
2. Transduction The most effective way in which transduction occurs is when, during the lytic cycle, a piece of host cell DNA gets packaged into a capsid of a newly formed virus accidentally.

38
2. Transduction When this occurs, as it sometimes does, the virus will inject only the DNA taken up from the host cell. If this DNA then crosses over and is taken up by the bacterium, recombination is said to occur.

39
Generalized Transduction
Generalized transduction occurs when bacterial genes are randomly transferred from one bacterial cell to another through a phage intermediate. It occurs when DNA from a bacterium gets accidentally inserted into a newly formed phage.

40
Specialized Transduction
In this process, a temperate phage (a phage that can use the lytic or lysogenic mode of reproduction), inserts its genome into a bacterial chromosome becoming a prophage. Specialized transduction occurs when the prophage picks up a few specific (adjacent) bacterial genes as it exits the chromosome transferring them to a new host cell.
, inserts its genome into a bacterial chromosome becoming a prophage. Specialized transduction occurs when the prophage picks up a few specific (adjacent) bacterial genes as it exits the chromosome transferring them to a new host cell.")
42
3. Conjugation Conjugation is basically bacterial sex. It is the direct transfer of genetic information from one cell to the next. The cells join briefly through the formation of a sex pilus which acts to pull the cells close together.

43
3. Conjugation Once the cells are close enough together, a cytoplasmic bridge forms between the two cells and an exchange of genetic information takes place. The ability to form sex pili depends on F factor which is a small segment of DNA in the bacterial chromosome or on a plasmid.

44
Plasmids Plasmids are small pieces of DNA that integrate into bacterial chromosomes and confer some sort of phenotypic change to it. R plasmids, for instance, often confer antibiotic resistance to various bacteria. It can also stand alone and replicate independently of the bacterium. When this happens, it is called an episome.

45
F Plasmids and Conjugation
F plasmids consist of about 25 genes and are responsible for the formation of sex pili. Cells with F plasmid are F+ which are DNA donors. Cells without F plasmid are F- and are the DNA recipients. F plasmids replicate with chromosomal DNA

46
F Plasmids and Conjugation
An F+ plasmid can turn an F- cell into an F+ cell when the two cells mix genes. One of the two strands of plasmid DNA is transferred to the F- plasmid. Each of the parental strands now serve as templates in each of the respective cells where DNA replication completes.

47
F Plasmids and Conjugation
When a donor cell’s F factor gets integrated into its chromosome, chromosomal genes can also get transferred during conjugation. Cells with F factor built into the chromosome are called Hfr cells (high frequency of recombination), and they are the donors.
, and they are the donors.")
48
F Plasmids and Conjugation
Many times when the two cells are joined together during conjugation, chromosomal DNA is brought into the F- cell. Random movements of the bacteria often prevent complete transfer of an F+ genome. The recipient cell now contains small amounts of DNA which often mixes with its DNA. This is the mechanism of recombination.

49
Bacterial Metabolism Bacteria can fine tune their metabolism to suit their varying metabolic demands. There are two ways in which the bacterium can control its metabolism: 1. They can adjust the enzymatic activity within the cell. 2. They can adjust the amount of product being made by certain enzymes--regulation of gene expression coding for enzymes.

50
The Operon An operon is a unit of genetic function common to bacteria and phages that consist of coordinately controlled clusters of genes with similar functions. An operon consists of an operator, a promoter, and the genes that code for the enzymes they control in a particular pathway.

51
The Basics of the Operon
The operon is the operator, the promoter and the genes they control. The operator is the segment of the DNA that acts as a switch. A repressor is a protein that can bind to the operon and switch it off. The repressor is usually a product of a regulatory gene often acting allosterically. Corepressors are small molecules that cooperate with repressor proteins to switch operons off.

52
The Basics of the Operon
Inducers are small molecules that inactivate repressors. Activators are proteins that bind to DNA and stimulate transcription of specific genes.

53
The Amino Acid Tryptophan: An Example
When the cell lacks tryptophan in its medium, a series of steps can be activated so that the cell can synthesize it. When tryptophan is synthesized from a precursor molecule, a series of steps are activated and each is catalyzed by a specific enzyme. The 5 genes that code for the enzymes are clustered together in a single stretch on the bacterial chromosome.

54
The Amino Acid Tryptophan: An Example
A single promoter serves all 5 genes. This constitutes a transcriptional unit, and when the mRNA is synthesized, it codes for all five enzymes in the pathway. The mRNA is punctuated with start and stop codons that signal where the coding sequences begin and end. Thus, the 5 separate polypeptides can be translated as needed.

55
The Amino Acid Tryptophan: An Example
The advantage to this is that there is one on-off switch for the series of genes related to tryptophan synthesis. When the cell needs tryptophan, all enzymes are switched on. The “switch” is the segment of DNA called the operator. The operator is positioned within or between the promoter and the enzyme-coding genes. The operator controls the access of RNA polymerase to the genes.

56
The Operon The operator, the promoter and the genes they control make up the operon.

57
The Operon Operons move back and forth between 2 states:
One with a repressor bound, and one without. Also, most regulatory proteins are allosterically controlled. That is, there is an active state and an inactive state. Controlled by the amount of gene product available.

58
The trp Operon, a Repressible Operon
The operon is switched off and on by a protein called a repressor. The operon, in this case, is always on and binding of the repressor blocks the attachment of RNA polymerase to the promoter and thus transcription of the genes. This blocking action is very specific and has no effect on other genes within the cell.

59
The trp Operon, a Repressible Operon
In our example using tryptophan, the trp repressor is the product of a regulatory gene (trpR) which is located some distance from the trp operon it is controlling. It has its own promoter. Regulatory genes are always on and always expressed within a cell. However, they bind reversibly.
 which is located some distance from the trp operon it is controlling. It has its own promoter. Regulatory genes are always on and always expressed within a cell. However, they bind reversibly.")
60
The trp Operon, a Repressible Operon
If the active genes ultimately produce tryptophan, and its available, it will bind to and alter the function of the trp repressor protein. That is, it binds to the protein, activating it and enabling it to bind to the operon shutting it off. Tryptophan is a corepressor.

61
The lac Operon The lac operon is an example of an inducible operon.
-galactosidase is an enzyme that breaks down lactose into glucose and galactose. Most E. coli cells only have a few molecules of -galactosidase available at any one time. When you add lactose to the medium in which they are growing, the amount of -galactosidase increases sharply.

62
The lac Operon The gene encoding for -galactosidase is part of the lac operon. It includes 2 other genes that code for enzymes that function in lactose metabolism. The whole thing is considered one transcriptional unit and is under the control of 1 operator and 1 promoter.

63
The lac Operon The lacI gene is a regulatory gene located outside of the operon. It encodes for an allosteric repressor protein that can switch off the lac operon by binding to the operator. The reason it is called an inducible operon is because the lac repressor is active by itself, and binds to the operator and shuts the lac operon off.

64
The lac Operon To inactivate the repressor, an inducer is needed. An inducer is a small molecule which binds to and inactivates a repressor. In the case of the lac operon, allolactose is the inducer. Allolactose is an isomer of lactose found in small amounts when lactose enters a cell.

65
The lac Operon When lactose is present, allolactose binds to the repressor altering its conformation, preventing the ability of the repressor from binding to the operator. When the repressor is unable to bind to the operator, the genes in the lac operon can be transcribed into mRNA which can be translated into lactose-metabolizing enzymes.

66
Summary The regulation of the trp and lac operons involves negative control of genes--their operons are switched off by the active form of the repressor gene. Gene regulation is said to be under positive control only when a regulatory protein interacts directly with the genome to switch transcription on.

67
Positive Control of the lac Operon
Not only are the lactose utilizing enzymes working and on when lactose is present, but they are also working when glucose is in short supply. E. coli cells can sense low concentrations of glucose and relay the information to the genome. It does so with the interaction of an allosteric regulatory protein and a small organic molecule.

68
Positive Control of the lac Operon
When glucose is scarce, cAMP accumulates in the cell. When accumulation occurs, it interacts with the regulatory protein called catabolite activator protein (CAP). CAP acts as an activator of transcription.
. CAP acts as an activator of transcription.")
69
Positive Control of the lac Operon
When cAMP binds to CAP, CAP assumes an active shape, binds to a specific site on the lac promoter stimulating gene expression. This is called positive regulation.

70
Positive Control of the lac Operon
When glucose levels within the cell increase, cAMP falls and CAP detaches from the operon. When CAP is inactive, transcription of the lac operon proceeds at a low level.

71
Summary The state of the lac repressor determines whether or not transcription of the lac operon’s genes occurs at all. The state of CAP controls the rate of transcription if the operon is repressor free. In both cases, when glucose is available, it will be used preferentially by the cell. Lactose enzymes will only be made when lactose is available and glucose is not.

72
Summary In our example involving tryptophan, the trp operon, is a repressible operon because its transcription is usually on but can be turned off. In contrast, the example involving lactose, the lac operon, is an inducible operon. Transcription is normall off, but can be turned on.

73
Summary When looking at the figure at the right, keep in mind the following 2 things: Negative control, binding has a negative effect, it shuts down transcription. Positive control, binding has a positive effect, it stimulates transcription.

74
Chapter 19: Viruses The components of a virus.
The differences between the lytic and lysogenic cycles.

75
Eukaryotic Genomes: Organization, Regulation and Evolution.
Chapter 19 Eukaryotic Genomes: Organization, Regulation and Evolution.

76
Chromatin The DNA-protein complex found in eukaryotes.
It is much more complex in eukaryotes than in prokaryotes.

77
The DNA Within Cells It undergoes a variety of changes as it proceeds through the cell cycle. Recall, in prophase it’s highly diffuse, but as the cell prepares to divide, it becomes highly condensed. Proteins called histones are responsible for the first level of DNA packing in chromatin. The mass of histone is nearly equal to the mass of DNA.

78
DNA-Histone Binding DNA is negatively charged, and histones contain a high proportion of positively charged aa’s and enable easy binding of the histones to the DNA.

79
DNA-Histone Binding Histones play a very important role in organizing DNA and they are very good at it. Thus, this is a likely reason why histone genes have been conserved throughout the generations in the course of evolution. The structure of histones are very similar among eukaryotes and between eukaryotes and prokaryotes.

80
DNA-Histone Binding and DNA Packing
Electron micrographs show unfolded chromatin and they look like beads on a string. These “beads” are referred to as nucleosomes (the basic unit of DNA packing), and the string is DNA.
, and the string is DNA.")
81
The Nucleosome and DNA Packing
A nucleosome is a piece of DNA wound around a protein core. This DNA-histone association remains in tact throughout the cell cycle. Histones only leave the DNA very briefly during DNA replication. With very few exceptions, histones stay with the DNA during transcription.

82
Nucleosome Interaction and DNA Packing
The next level of DNA packing takes place between the histone tails of one nucleosome/linker DNA and the nucleosomes to either side. The interactions between these cause the DNA to coil even tighter. As they continue to coil and fold, eventually the DNA resembles that of the metaphase chromosome.

83
Heterochromatin Vs. Euchromatin
During interphase, some of the DNA remains condensed as you would normally see it in metaphase. (centrosomes, telomeres, and some other regions of the chromosome). This is called heterochromatin to distinguish it from euchromatin which condenses and relaxes with the cell cycle. Heterochromatin is rarely transcribed.
. This is called heterochromatin to distinguish it from euchromatin which condenses and relaxes with the cell cycle. Heterochromatin is rarely transcribed.")
84
Acetylation There is a lot of evidence supporting the notion that the regulation of gene expression is, in part, dependent upon chemical modifications to histones. When an acetyl group is added to the histone tail, the histones become neutralized and the chromatin loosens up. As a result, transcription can occur.

85
Methylation Addition of a methyl group to a histone tail leads to condensation of the chromatin.

86
DNA Methylation DNA methylation is completely separate from histone methylation, but may be a way in which genes become inactivated. Evidence: Inactivated X chromosomes are heavily methylated. In many cells that have inactivated genes, the genes are more heavily methylated than in cells where the genes are active.

87
Control of Eukaryotic Gene Expression
Recall the idea of the operon and how it regulated bacterial gene expression. The mechanism of gene expression in eukaryotes is different. It involves chromatin modifications, but they do not involve a change in DNA sequence.

88
Chromatin Modifying Enzymes
These provide initial control of gene expression. They make the region of DNA more or less able to bind DNA machinery. Once optimized for expression, the initiation of transcription is the most universally used stage at which gene expression is regulated.

89
Recall, Eukaryotic genes have promoters, a DNA sequence where RNA polymerase II binds and starts transcription. There are numerous control elements involved in regulating the initiation of transcription. 5’ caps. Poly-A tails.

90
Also, RNA modifications help prevent enzymatic degradation of mRNA, allowing more protein to be made.

91
Recall, RNA processing involves 3 steps: 1. Addition of the 5’ cap.
2. Addition of the poly-A tail. 3. Gene splicing. Removal of introns and splicing together of exons.

92
Recall, The transcription initiation complex assembles on the promoter sequence. RNA polymerase II proceeds to transcribe the gene making pre-mRNA. Transcription factors are proteins that assist RNA polymerase II to initiate transcription.

93
Eukaryotic Gene Expression
Most eukaryotic genes are associated with multiple control elements which are segments of non-coding DNA that help regulate transcription by binding certain proteins. These control elements are crucial to the regulation of certain genes within different cells.

94
Eukaryotic Gene Expression
Only after the complete initiation complex has assembled can the polymerase begin to move along the DNA template strand, producing a complementary strand of DNA.

95
Eukaryotic Gene Expression
In eukaryotes, high levels of transcription of a particular gene at the appropriate time depends on the interaction of control elements with other proteins called transcription factors. Enhancers and activators play important roles in gene expression. Enhancers are nucleotide sequences that bind activators and stimulate gene expression.

96
Enhancer-Activator Interaction and Eukaryotic Gene Expression
When the activators bind to the enhancers, this causes the DNA to bend allowing interaction of the proteins and the promoter. This helps to position the initiation complex on the promoter so RNA synthesis can occur.

97
Eukaryotic Gene Expression
Some specific transcription factors function as repressors to inhibit expression of a particular gene. Certain repressors can block the binding of activators either to their control elements or to parts of their transcriptional machinery. Other repressors bind directly to their own control elements in an enhancer and act to turn off transcription.

99
Eukaryotic Gene Expression
There are only a dozen or so short nucleotide sequences that exist in control elements for different genes. The combinations of these control elements are more important than the presence of single unique control elements in regulating the transcription of a gene.

100
Recall, Prokaryotes typically have coordinately controlled genes clustered in an operon. The operons are regulated by single promoters and get transcribed into a single mRNA molecule. Thus genes are expressed together, and proteins are made concurrently.

101
Control of Eukaryotic Gene Expression
Recent studies indicate that within genomes of many eukaryotic species, co-expressed genes are clustered near one another on the same chromosome. However, unlike the genes in the operons of prokaryotes, each of the eukaryotic genes have their own promoter and is individually transcribed. It is thought that the coordinate regulation of genes clustered in eukaryotic cells involves changes in chromatin structure that makes the entire group of genes available or unavailable.

102
Control of Eukaryotic Gene Expression
More commonly, co-expressed eukaryotic genes are found scattered over different chromosomes. In these cases, coordinate gene expression is seemingly dependent on the association of specific control elements or combinations of every gene of a dispersed group. Copies of activators that recognize these control elements bind to them, promoting simultaneous transcription of the genes no matter where they are in the genome.

103
Control of Eukaryotic Gene Expression
The coordinate control of dispersed genes in a eukaryotic cell often occurs in response to external signals such as hormones. When the steroid enters the cell, it binds to a specific intracellular receptor protein forming a hormone-receptor complex that serves as a transcription activator.

104
Control of Eukaryotic Gene Expression
In an alternative mechanism, a signal molecule such as a non-steroid hormone or a growth factor bind to a receptor on a cell’s surface and never enter a cell. Instead, they control gene expression by inducing a signal transduction pathway.

105
Post-transcriptional Regulation and Control of Gene Expression
The mechanisms we’ve just discussed involve regulating the expression of the gene. Post-transcriptional regulation involves regulating the transcript after the mRNA has been made. These modes are unique to eukaryotes.

106
Alternative RNA Splicing and Control of Gene Expression
Alternative RNA splicing is a way in which different mRNA transcripts are produced from the same primary transcript. This is determined by which RNA segments are treated as introns and which are treated as exons.

107
Alternative RNA Splicing and Control of Gene Expression
Different cells have different regulatory proteins that control intron-exon choices by binding to regulatory sequences within the primary transcript.

108
Alternative Mechanisms to Control Gene Expression
Protein processing is the final spot for controlling gene expression. Often, eukaryotic polypeptides undergo further processing to yield a functional protein. Regulation can occur at any of the sites of protein modification.

109
Chapter 20: Biotechnology
The terminology of biotechnology. The steps in gene cloning with special attention to the biotechnology tools that make cloning possible. The key ideas that make PCR possible. How gel electrophoresis can be used to separate DNA fragments or protein molecules.

110
DNA Technology and Genomics
Chapter 20 DNA Technology and Genomics

111
The Challenges of Studying DNA
The size of the DNA poses big problems. Naturally occurring DNA is very long and particular genes may only comprise a small portion of the DNA, maybe 1/100,000 of the chromosome. There may only be a small difference in the surrounding nucleotides.

112
Help in Overcoming These Challenges
To make this easier, scientists have developed methods for gene cloning. Genes are cloned to make multiple copies of the same gene and to produce a protein product. Cloning can be used to endow an organism with a new metabolic activity. Pest resistance, drought resistance, etc. We can also insert genes into organisms for medical purposes. Insulin production by E. coli.

113
Most Methods for Cloning DNA Share General Features:
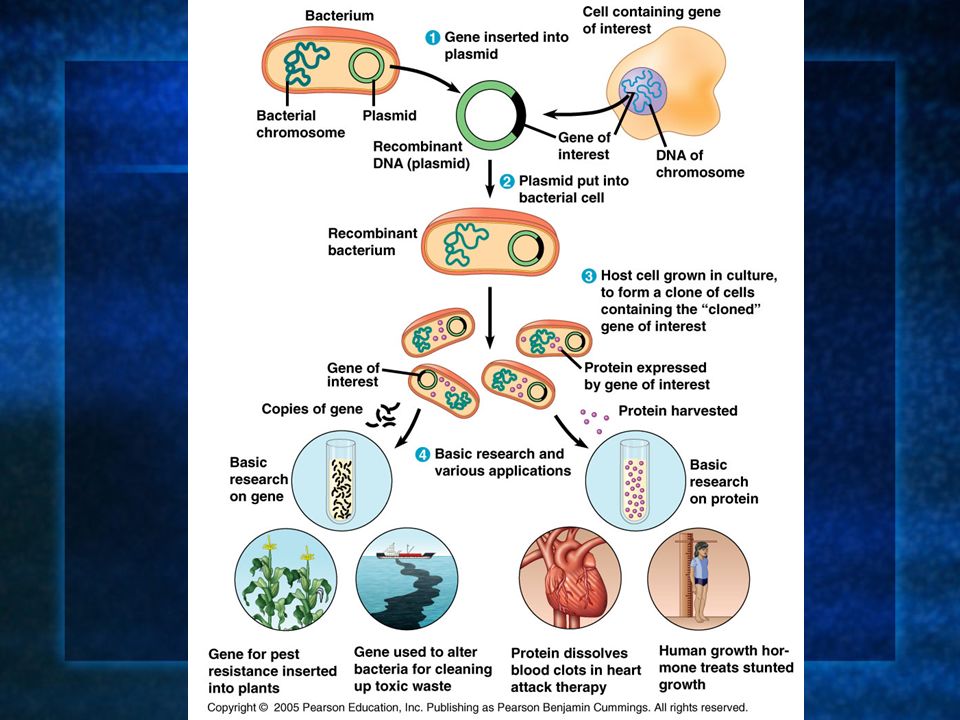
A commonly used method uses bacteria (E. coli) and their plasmids: 1. The plasmid is isolated from bacteria. 2. DNA is inserted into it. 3. The plasmid is now recombinant (DNA from 2 sources). 4. The bacteria reproduces forming a clone of the original cell. 5. The foreign (inserted) gene is cloned at the same time.
 and their plasmids: 1. The plasmid is isolated from bacteria. 2. DNA is inserted into it. 3. The plasmid is now recombinant (DNA from 2 sources). 4. The bacteria reproduces forming a clone of the original cell. 5. The foreign (inserted) gene is cloned at the same time.")
115
Restriction Enzymes These are enzymes that cut DNA molecules at a limited number of specific locations. In nature, these enzymes help prevent a bacterial cell from foreign DNA (from phages and other organisms). Many different restriction enzymes have been identified and isolated.
. Many different restriction enzymes have been identified and isolated.")
116
Restriction Enzymes Each restriction enzyme is very specific and recognizes a short DNA sequence known as a restriction site. The DNA itself is cut at specific sites within the DNA strand. A bacterial cell will protect its own DNA from its own restriction enzymes by addition of methyl (-CH3) groups to A’s and C’s within the sequences recognized by these enzymes.
 groups to A’s and C’s within the sequences recognized by these enzymes.")
117
Restriction Enzymes They recognize sequences 4-8 nucleotides in length. Many such sequences occur by chance throughout the genome, thus a restriction enzyme will produce a numerous amount of fragments (called restriction fragments) when they are introduced to DNA.
 when they are introduced to DNA.")
118
Restriction Enzymes All copies of a particular DNA molecule always produce the same DNA fragments when introduced to the same restriction enzymes. Thus, a restriction enzyme cuts DNA in a reproducible way.

119
Restriction Enzymes The most useful RE’s cleave DNA in a certain way and produce sticky ends. We call them sticky ends because they combine with other DNA fragments that have been cut by the same enzyme. These fragments usually hydrogen bond together and then are joined permanently by DNA ligase which catalyzes the formation of covalent bonds in the sugar-phosphate backbones. This produces a stable, recombinant DNA molecule.

120
Gene Cloning in Plasmids
Genes are cloned in plasmids using a cloning vector which is the original plasmid that carries the foreign DNA into the cell. Bacterial plasmids are commonly used because they are easy to manipulate and most of the experimentation with them can be done in vitro.

121
A Common Method for Cloning:
1. Isolation of plasmids from E. coli cells and DNA from human cells grown in culture. 2. Treat the 2 with the same RE producing the same sticky ends. Plasmids cut at one spot, human DNA cut at many. 3. Mix the human and plasmid fragments. 4. Add DNA ligase to permanently fuse the sticky ends of the plasmid and human DNA. 5. Mix the recombinant plasmids with bacteria. 6. Plate the bacteria out on selective media to isolate the recombinants.

123
Recognizing the Clone Originally, the bacteria lacked a gene conferring resistance to something, say an antibiotic. When the bacterium was plated on growth medium containing the antibiotic, they would die. Those bacteria containing the antibiotic resistance gene are now able to survive on the medium containing antibiotic.

125
Problems with Cloned DNA and Expression in the Bacteria
Two different types of cells (prokaryotic vs. eukaryotic) pose problems for gene expression. The presence of introns in eukaryotic transcripts and no way for the prokaryotes to splice them out.
 pose problems for gene expression. The presence of introns in eukaryotic transcripts and no way for the prokaryotes to splice them out.")
126
2 Different Cell Types To overcome this, scientists use expression vectors that have highly active prokaryotic promoters just upstream from the gene to be expressed. The bacterial cell will now recognize the promoter and express the foreign DNA linked to the promoter. In this way, many eukaryotic genes are expressed in prokaryotic cells.

127
The Presence of Introns
Prokaryotes lack splicing machinery, and the long eukaryotic gene is often prevented from being expressed in the bacteria. Scientists use cDNA to circumvent the problem. The cDNA contains only the exons.

128
cDNA Synthesis cDNA is synthesized from mRNA extracted from the cells.
Retroviruses make reverse transcriptase and this is used to make single stranded DNA molecules from the mRNA. mRNA gets enzymatically degraded and DNA polymerase then synthesizes a second strand of the DNA. The cDNA is modified with REs to ease the transition into plasmids and then bacterial cells.

129
cDNA Synthesis Scientists can overcome the differences between prokaryotes and eukaryotes using expression vectors. cDNA is used and will be expressed by the bacterial cell as long as the expression vector contains the bacterial promoter and other control elements necessary for transcription and translation.

130
Other Ways to Get Around Compatibility Problems:
Scientists often use eukaryotic yeasts and single-celled fungi which are as easy to grow as bacteria and also contain plasmids. Scientists have also made recombinant plasmids that combine yeast and bacterial DNA that can replicate in either type of cell.

131
PCR Sometimes scientists want to prepare a large quantity of DNA when only a small amount is present. To get around this, PCR is used and can quickly generate a large amount of DNA from a small amount.

132
PCR A 3-step cycle brings about a chain reaction that produces exponential growth of identical DNA molecules. A double stranded piece of of DNA is obtained. A. The solution containing the piece of DNA is heated so as to denature the DNA and separate it into single strands. B. DNA primers (short, single stranded DNA molecules) are added to the mixture and it is allowed to cool so the primers anneal to the cDNA strands. C. The heat-stable DNA polymerase adds nucleotides to the primers in the standard 5’-->3’ direction synthesizing the target sequence.
 are added to the mixture and it is allowed to cool so the primers anneal to the cDNA strands. C. The heat-stable DNA polymerase adds nucleotides to the primers in the standard 5’-->3’ direction synthesizing the target sequence.")
134
PCR What makes this process so useful is its specificity. If a target segment is identified and a primer made to it, then only a small amount is really necessary from the start. It is easy to see how quickly a large amount of DNA can be made: 1, 2, 4, 8, 16, …..

135
PCR The primer will only replicate the target segment because this is all they are able to bind to. After just a few cycles, a very large amount of the target segment will be identified.

136
PCR DNA cloning in cells remains the best way to prepare a large quantity of a gene or DNA segment. PCR can’t be used to obtain a large quantity of gene because occasional errors in PCR replication impose limits on the number of good copies that can be made. Often times though, enough of a specific DNA fragment can be made to insert it into a vector and clone it.

137
Gel Electrophoresis To study DNA, scientists often use gel electrophoresis. Agarose gel is often used to separate DNA fragments based on size, charge, etc., that have been treated with a restriction enzyme. The fibers in the gel separate out the fragments; smaller fragments migrate further than the larger fragments.

138
Gel Electrophoresis The negatively charged DNA fragments migrate toward the positive pole of the electrophoresis box.

139
Restriction Fragment Analysis
This is a powerful tool scientists use to analyze differences in the nucleotide sequences of DNA molecules. When the researchers began analyzing the restriction fragments of non-coding DNA from individuals, they began noticing small nucleotide differences on homologous chromosomes.

140
Restriction Fragment Analysis
Treating the DNA with restriction enzymes and then running the samples through a gel enable researchers to produce banding patterns characteristic of the starting molecule and the restriction enzyme(s) used to treat the DNA. You will use this analysis to examine your bacterial chromosome for certain genes.
 used to treat the DNA. You will use this analysis to examine your bacterial chromosome for certain genes.")
144
Restriction Fragment Length Polymorphisms
When non-coding regions of DNA were treated with restriction enzymes and banded, scientists discovered differences in non-coding regions on homologous chromosomes. These were given the name restriction fragment length polymorphisms.

145
Restriction Fragment Length Polymorphisms
These serve as genetic markers of non-coding DNA that appears near a particular locus in a genome. There are many RFLP variants within a population. RFLP data is often used in crime investigations because the likelihood that two individuals will have the same banding patterns are miniscule at best.

146
Scientists use a Southern Blot to Analyze RFLPs or Regular DNA.

147
Why is This Important? It is used in crime scene investigations and paternity suits all the time.

148
Why is This Important? Scientists use the information to determine if a person has a particular disease.

149
Why is This Important? Scientist can also use RFLP information to determine the likelihood of inheriting a certain genetic disease.

150
Chapter 21: Genomes and Their Evolution

151
The Genetic Basis of Development
Chapter 21 The Genetic Basis of Development

152
Zygote and Cell Division
When the zygote divides, it undergoes 3 major changes: 1. Cell division 2. Cell differentiation 3. Morphogenesis

153
Cell Signaling Cell signaling is largely responsible for the developmental processes.

154
1. Cell Division Cell division gives rise to numerous cells.

155
2. Cell Differentiation Cell differentiation is the process by which cells become specialized in form and function. These cells undergo changes that organize them into tissues and organs.

156
3. Morphogenesis As the dividing cells begin to take form, they are undergoing morphogenesis which means the “creation of form.” Morphogenetic events lay out the development very early on in development as cell division, cell differentiation and morphogenesis overlap.

157
3. Morphogenesis These morphogenetic events “tell” the organism where the head and tail are, which is the front and back, and what is left and right. As time progresses, later morphogenetic events will give instructions as to where certain appendages will be located.

158
Morphogenetic Events Morphogenetic events, as well as cell division and differentiation, take place in all multicellular organisms. Morphogenesis differs in 2 major ways in plants and animals: 1. In animals, movements of cells and tissues are required for the transformation of the early embryo into the characteristic 3D form of the organism. 2. In plants, morphogenesis and growth in overall size are not limited to embryonic and juvenile periods, they occur throughout the life of the plant.

160
Apical Meristems For example, apical meristems of plants are responsible for a plant’s continued growth and development and the formation of new organs throughout the plant’s life. These are perpetually embryonic regions in the tips of shoots and roots.

161
Totipotent Cells Totipotency is the ability of a single cell to divide and produce all the differentiated cells in an organism, including extraembryonic tissues. Totipotent cells are formed as a result of sexual reproduction.

162
Pluripotent In cell biology, the definition of pluripotency has come to refer to a stem cell that has the potential to differentiate into any of the three germ layers: endoderm, mesoderm, or ectoderm. Pluripotent stem cells can give rise to any fetal or adult cell type. However, alone they cannot develop into a fetal or adult animal because they lack the potential to contribute to extraembryonic tissue, such as the placent.

164
Multicellular Organisms
The cells of multicellular organisms come almost entirely from differences in gene expression. Regulatory mechanisms turn certain genes on and off during development. These regulatory mechanisms are what makes cells different because nearly all cells have the same genetic complement.

165
Cloning Using the somatic cells of a multicellular organism to generate a new organism is called cloning. Each clone is genetically identical to the parent plant. Differentiated cells don’t usually divide in culture, so researchers had to take a different approach to decide if animal cells were totipotent.

166
What Researchers Did… They removed the nucleus of an unfertilized egg and replaced it with one from a differentiated cell. The process is called nuclear transplantation. If the transplanted cell retains all of its genetic information, the recipient cell should develop with all of the necessary tissues and organs.

167
Nuclear Transplantation
As these experiments were conducted on frogs, it was determined that something in the DNA does change. In tadpoles, normal development proceeded, but as the age of the donor nucleus increased, the percentage of organisms that developed correctly decreased.

168
Nuclear Transplantation
Continued research showed that the DNA remains the same for the most part, but the chromatin changes in a way that problems arise.

169
Nuclear Transplantation
Often times, the histones get modified or DNA is methylated and these changes in the chromatin prevent dedifferentiation. Sometimes the process is reversible, but usually it isn’t. One thing is certain, most scientists agree that all cells contain the necessary genetic information to make an entire organism. However, the different cell types exist because of the variations in gene expression.

170
Nuclear Transplanting and Cloning
In 1997, Scottish researchers cloned a sheep named Dolly. They used cells from mammary tissue in an adult sheep, implanted the nucleus from the cell into egg cells from which the nucleus had been removed and implanted into the uterus of a lamb.

172
Nuclear Transplanting and Cloning
Analysis of the DNA from Dolly showed it was identical to that of the original sheep, and its mitochondria matched that of the mother lamb. However, Dolly’s cells appeared older than her age would indicate.

173
Stem Cells The use of stem cells, especially embryonic stem cells, has many obvious medical applications. There are obvious ethical dilemmas that arise from the research. There are moral issues on both sides: One is that it is immoral to tamper with human embryos for medical purposes. The other is that it is immoral not to because the benefits outweigh the cost of doing nothing.

175
Cell Differentiation There are 2 major things telling a cell when and how to differentiate: 1. The “stuff” found within the egg at the time of conception. 2. The environment in which the embryo develops.

176
1. The “Stuff” in the Egg The egg cell’s cytoplasm contains RNA and protein molecules encoded by the mother’s DNA. mRNA, proteins, organelles, and other substances are scattered unevenly throughout the cytoplasm of an unfertilized egg. These maternal substances influence the course of early development called cytoplasmic determinants.

177
Cytoplasmic Determinants
Following fertilization, mitotic divisions distribute the zygote’s cytoplasm into separate cells. The nuclei of these cells are subjected to many different cytoplasmic determinants. What has been received will determine the developmental fate of each of the cells.

178
Cytoplasmic Determinants
Cytoplasmic determinants help to create an animal’s 3D arrangement before morphogenesis can shape the animal.

179
2. The Environment The environment in which the embryo develops plays an important factor in outcome of the developing organism. The surface contact of cell-to-cell interaction helps to signal development. By the process of induction, an embryo’s genes signal the expression of proteins that cause changes in nearby target cells. These signals send a cell down a specific developmental pathway--inducing further differentiation of the many specialized cells within the new organism.

181
Pattern Formation Pattern formation in animals begins in the embryo when the major axes are determined. Before tissues and organs within an animal can be formed, the 3D arrangement must be established. Recall that this occurs as a result of cytoplasmic determinants. This process has been extensively studied in many animals such as the fruit fly, sea urchin, frog, nematode, and chicken.

182
Apoptosis Apoptosis is the programmed cell death that occurs through the normal course of development. It is usually triggered by signals that activate a cascade of signal proteins in cells that are to die. During the process, the cell shrinks, the nucleus breaks down and the nearby cells quickly engulf and break down the contents of the cell.

184
Apoptosis Apoptosis is essential to the development of all cells. The process helps in the growth and development of the major structures and systems of an organism. It controls cell division helping to slow or stop division in certain cells.

Similar presentations





















 cells retain a complete set of genes –In general,>")










 Cannot live outside of a cellular host Do not.>")




