Download presentation
Presentation is loading. Please wait.

1
Dr. Nasir Jalal ASAB/NUST
Enzymology Lecture 5 Dr. Nasir Jalal ASAB/NUST

2
Question: What do cells surf on?
Answer: Microwaves.

3
Linear plot of Michaelis-Menten
The plot of v versus [S] is not linear; although initially linear at low [S], it bends over to saturate at high [S]. Before the modern era of nonlinear curve-fitting on computers, this nonlinearity could make it difficult to estimate KM and Vmax accurately. Therefore, several researchers developed linearisations of the Michaelis–Menten equation, such as the Lineweaver–Burk plot, the Eadie–Hofstee diagram and the Hanes–Woolf plot. All of these linear representations can be useful for visualising data, but none should be used to determine kinetic parameters, as computer software is readily available that allows for more accurate determination by nonlinear regression methods

4
Why double reciprocal Plotting 1/v (reciprocal of rate) vs. 1/[s] (reciprocal of substrate concentration) yields a straight line. One can then use the equation of the linear regression to calculate Vmax and Km.
 vs. 1/[s] (reciprocal of substrate concentration) yields a straight line. One can then use the equation of the linear regression to calculate Vmax and Km.")
5
Lineweaver–Burk plot The Lineweaver–Burk plot or double reciprocal plot is a common way of illustrating kinetic data. This is produced by taking the reciprocal of both sides of the Michaelis–Menten equation. This is a linear form of the Michaelis–Menten equation and produces a straight line with the equation: y = mx + c with a y-intercept equivalent to 1/Vmax and an x-intercept of the graph representing -1/KM Any function: y = f(x) that graphs as a straight line has an equation of this form: y = mx + c 'x' and 'y' are the coordinates of the points that satisfy the function and so lie on the straight line graph. 'm' is the gradient of the straight line graph, and 'c' is the 'y intercept' of the straight line graph.
 that graphs as a straight line has an equation of this form: y = mx + c. x and y are the coordinates of the points that satisfy the function and so lie on the straight line graph. m is the gradient of the straight line graph, and. c is the y intercept of the straight line graph.")
6
Double reciprocal used to calculate Km
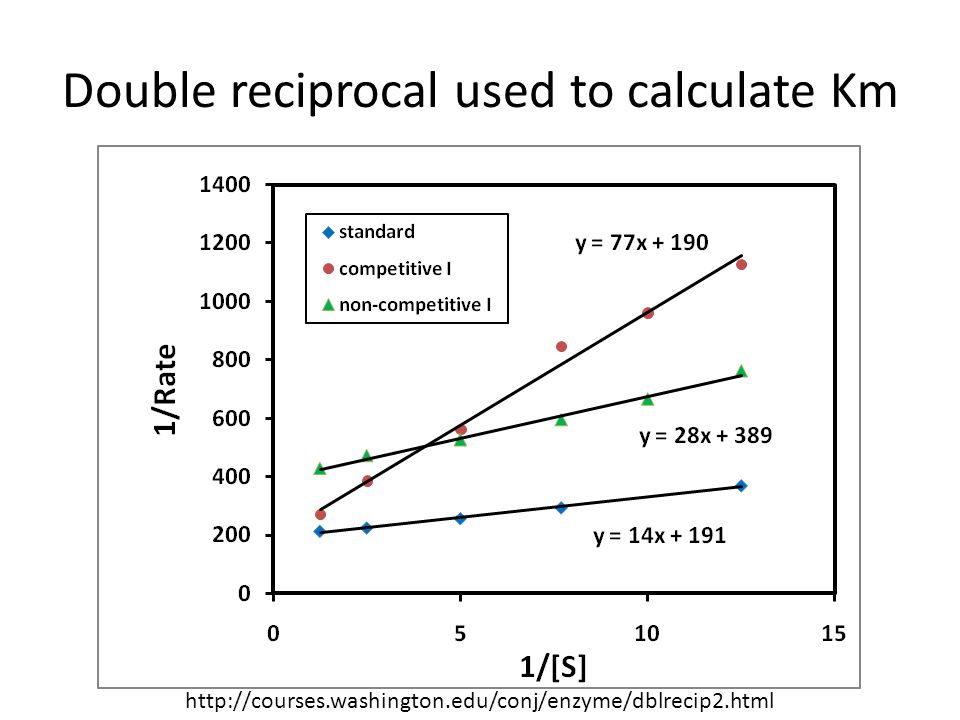
7
Quick Quiz When you plot 1/v vs. 1/[s] you get a straight line. What is the y-intercept? What is the slope for this line? Calculate Vmax for the standard data shown above. (You may give answer or just set problem up.) Calculate Km for the standard data shown above. (You may give answer or just set problem up.) 1/Vmax Km/Vmax Vmax=1/191=0.0052 Km=14*Vmax=0.073
![Quick Quiz When you plot 1/v vs. 1/[s] you get a straight line. What is the y-intercept What is the slope for this line](http://slideplayer.com/slide/1399057/3/images/7/Quick+Quiz+When+you+plot+1%2Fv+vs.+1%2F%5Bs%5D+you+get+a+straight+line.+What+is+the+y-intercept+What+is+the+slope+for+this+line.jpg "Calculate Vmax for the standard data shown above. (You may give answer or just set problem up.) Calculate Km for the standard data shown above. (You may give answer or just set problem up.) 1/Vmax. Km/Vmax. Vmax=1/191= Km=14*Vmax=")
8
Advantage and disadvantage of LWP
When used for determining the type of enzyme inhibition, the Lineweaver–Burk plot can distinguish competitive, non-competitive and uncompetitive inhibitors. Competitive inhibitors have the same y-intercept as uninhibited enzyme (since Vmax is unaffected by competitive inhibitors the inverse of Vmax also doesn't change) but there are different slopes and x-intercepts between the two data sets. Non-competitive inhibition produces plots with the same x-intercept as uninhibited enzyme (Km is unaffected) but different slopes and y-intercepts. Uncompetitive inhibition causes different intercepts on both the y- and x-axes but the same slope. Disadvantage: The Lineweaver–Burk plot was classically used but it is prone to error, as the y-axis takes the reciprocal of the rate of reaction thus increasing any small errors in measurement. Also, most points on the plot are found far to the right of the y-axis (due to limiting solubility not allowing for large values of [S] and hence no small values for 1/[S]), calling for a large extrapolation back to obtain x- and y-intercepts.
 but there are different slopes and x-intercepts between the two data sets. Non-competitive inhibition produces plots with the same x-intercept as uninhibited enzyme (Km is unaffected) but different slopes and y-intercepts. Uncompetitive inhibition causes different intercepts on both the y- and x-axes but the same slope. Disadvantage: The Lineweaver–Burk plot was classically used but it is prone to error, as the y-axis takes the reciprocal of the rate of reaction thus increasing any small errors in measurement. Also, most points on the plot are found far to the right of the y-axis (due to limiting solubility not allowing for large values of [S] and hence no small values for 1/[S]), calling for a large extrapolation back to obtain x- and y-intercepts.")
9
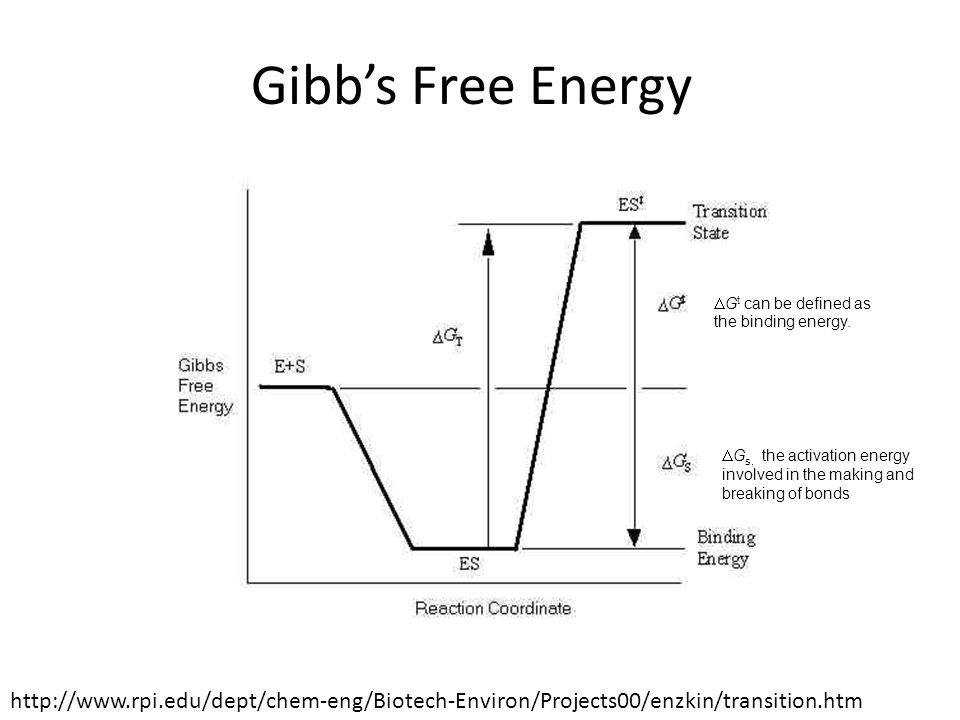
Transition State Model
For a reaction involving two molecules, a transition state is formed when the old bonds between two molecules are weakened and new bonds begin to form or the old bonds break first to form the transition state and then the new bonds form after. The theory suggests that as reactant molecules approach each other closely they are momentarily in a less stable state than either the reactants or the products. In the example below, the first scenario occurs to form the transition state: Old bonds between molecules (hydroxide and bromomethane) weaken New bonds begin to form. Less stable state. New molecule with new bonds forms, Bromide released
 weaken. New bonds begin to form. Less stable state. New molecule with new bonds forms, Bromide released.")
10
The equation for an enzymatic reaction is:
K‡ is the concentration equilibrium constant, defined as: This equation can be used to find out the rate constant K Where, kB is Boltzmann’s constant, h is Planck’s constant and T is the temperature Boltzmann constant = × m2 kg s-2 K-1 Planck's constant = × m2 kg / s

11
Gibb’s Free Energy K‡ concentration equilibrium constant resembles the equilibrium constant used to describe Gibbs free energy, defined as: where co is the standard state concentration. DGt can be defined as the Gibbs energy of activation. The Gibbs energy difference between the ground and transition state can be used to predict the rate of reaction. The binding energy associated with the specific substrate-enzyme interaction is a significant factor in lowering the Gibbs free energy change required for reaction. The large binding energies of substrates are due in part to the complementary shape of the active site of the enzyme. The Gibbs energy can be considered to be composed of two terms, DGt, the binding energy and DGs, the activation energy involved in the making and breaking of bonds leading to the transition state from enzyme-substrate intermediate (ES). They are related as follows: DGt = DGt + DGs
. They are related as follows: DGt = DGt + DGs.")
12
Gibb’s Free Energy DGt can be defined as the binding energy. DGs, the activation energy involved in the making and breaking of bonds
13
Using Gibbs free energy
In a problem X was trying to figure out if this reaction will proceed as written: (is the end result a positive/unfavorable or negative/favorable reaction). (DHAP) --> (GAP) Standard free energy change ΔG° = +1.8 kcal/mol. Concentration of DHAP = 8 Concentration of GAP = .5 The gas constant (R) is kcal/mol temp in cell = 298 K And my equation that I should be using is: ΔG = ΔG° + RTln([products]/[reactants]) So I end up with: ΔG = (1.987 * 298)ln(0.5/8) = Is this done? It seems like a very improbable answer . Or is there a NEED to factor in the temperature because the gas constant is calculated for the cell temp already? And then WE would end up with: (1.987)ln(0.5/8) = Your gas constant is wrong isn't it? Shouldn't it be cal/°K/mol, rather than kcal/mol? Delta G = (( x 298) x ln (0.5/8)) = (0.592 x ) = ( ) = 0.158
. (DHAP) --> (GAP) Standard free energy change ΔG° = +1.8 kcal/mol. Concentration of DHAP = 8 Concentration of GAP = .5 The gas constant (R) is kcal/mol temp in cell = 298 K And my equation that I should be using is: ΔG = ΔG° + RTln([products]/[reactants]) So I end up with: ΔG = (1.987 * 298)ln(0.5/8) = Is this done It seems like a very improbable answer . Or is there a NEED to factor in the temperature because the gas constant is calculated for the cell temp already And then WE would end up with: (1.987)ln(0.5/8) = Your gas constant is wrong isn t it Shouldn t it be cal/°K/mol, rather than kcal/mol Delta G = (( x 298) x ln (0.5/8)) = (0.592 x ) = ( ) =")
14
Because he worked for the special branch.
Question: Why did the policeman hide under a tree? Because he worked for the special branch.

15
Quantum tunneling model of Enzyme activity

16
Heisenberg’s Uncertainty Principle involving energy and time
The more precisely the position is determined, the less precisely the momentum is known in this instant, and vice versa. --Heisenberg, uncertainty paper, 1927 If our measurement lasts a certain time Dt, then we cannot know the energy better than an uncertainty DE

17
Quantum Tunneling Model through an example
To understand the phenomenon, particles attempting to travel between potential barriers can be compared to a ball trying to roll over a hill; quantum mechanics and classical mechanics differ in their treatment of this scenario. Classical mechanics predicts that particles that do not have enough energy to classically surmount a barrier will not be able to reach the other side. Thus, a ball without sufficient energy to surmount the hill would roll back down. Thus, a ball without sufficient energy to surmount the hill would roll back down. Or, lacking the energy to penetrate a wall, it would bounce back (reflection) or in the extreme case, bury itself inside the wall (absorption). In quantum mechanics, these particles can, with a very small probability, tunnel to the other side, thus crossing the barrier. The ball could, in a sense, borrow energy from its surroundings to tunnel through the wall or roll over the hill, paying it back by making the reflected electrons more energetic than they otherwise would have been
 or in the extreme case, bury itself inside the wall (absorption). In quantum mechanics, these particles can, with a very small probability, tunnel to the other side, thus crossing the barrier. The ball could, in a sense, borrow energy from its surroundings to tunnel through the wall or roll over the hill, paying it back by making the reflected electrons more energetic than they otherwise would have been.")
18
Quantum Tunneling Model through an example
Normally, the car can only get as far as C, before it falls back again But a fluctuation in energy could get it over the barrier to E!

19
Early concepts Warshel and Levitt's study of lysozyme was pioneering in this area. Warshel, A and Levitt, M J. Mol. Biol., 103: 227 [CrossRef], [PubMed], [Web of Science ®] The early ab initio QM/MM implementation by Singh and Kollman is notable. The semiempirical QM/MM method developed by Field et al.

20
Rules of Quantum Tunneling
A particle ‘borrows’ an energy DE to get over a barrier Does not violate the uncertainty principle, provided this energy is repaid within a certain time Dt The taller the barrier, the less likely tunneling would occur

21
Quantum tunneling phenomenon
Quantum tunneling through a barrier. At the origin (x=0), there is a very high, but narrow potential barrier. A significant tunneling effect can be seen. Quantum tunneling through a barrier. The energy of the tunneled particle is the same but the amplitude is decreased.
, there is a very high, but narrow potential barrier. A significant tunneling effect can be seen. Quantum tunneling through a barrier. The energy of the tunneled particle is the same but the amplitude is decreased.")
22
Electron wavepacket simulation
An electron wavepacket directed at a potential barrier. Note the dim spot on the right that represents tunnelling electrons.

23
A can toppling over due to quantum fluctuations of its position
Have to wait about years!!

24
QM/MM modeling Some enzymes operate with kinetics which are faster than what would be predicted by the classical ΔG‡. In "through the barrier" models, a proton or an electron can tunnel through activation barriers. Quantum tunneling for protons has been observed in tryptamine oxidation by aromatic amine dehydrogenase.

25
Chorismate mutase and QM/MM
An enzyme (chorismate mutase) partitioned into a QM region (shown as red spheres) and an MM region consisting of protein (yellow cartoons) and solvent (cyan sticks). Here the system is truncated to an approximate sphere (in this case with radius 25 Å), typical of the approach used in many QM/MM simulations of enzyme-catalysed reaction mechanisms (e.g. applying ‘stochastic boundary’)
 partitioned into a QM region (shown as red spheres) and an MM region consisting of protein (yellow cartoons) and solvent (cyan sticks). Here the system is truncated to an approximate sphere (in this case with radius 25 Å), typical of the approach used in many QM/MM simulations of enzyme-catalysed reaction mechanisms (e.g. applying ‘stochastic boundary’)")
26
Theoretical basis of QM/MM
The theoretical basis of QM/MM methods are outlined here. The system is divided into a (small) QM region and an (usually much larger) MM region. The total energy of a QM/MM system can be expressed as: E QM and E MM are the energies of the QM and MM regions, respectively, calculated in a standard way at those levels. For example, the MM energy is defined by the potential of the force field used.
 QM region and an (usually much larger) MM region. The total energy of a QM/MM system can be expressed as: E QM and E MM are the energies of the QM and MM regions, respectively, calculated in a standard way at those levels. For example, the MM energy is defined by the potential of the force field used.")
27
Theoretical basis of QM/MM
Current standard MM force fields for proteins treat atoms as point charges with van der Waals radii, which together determine the non-bonded interactions. Force fields also include terms to describe interactions between bonded atoms, e.g. bond, angle and dihedral terms. E QM/MM describes the interaction between the QM and MM regions, and can be treated in various ways. E Boundary is a term to account for the fact that the system may have to be truncated for practical reasons (e.g. QM/MM calculations are computationally demanding, and it may not be possible or desirable to include the whole of an enzyme molecule).
.")
28
Considerations for QM/MM
Broadly speaking, there are two particularly important considerations in the interaction between the QM and MM regions: (i) the treatment of non-covalent interactions between the QM and MM regions and (ii) where covalent bonds exist between atoms in the QM and MM regions, the treatment of bonds at the frontier between the two regions.
 the treatment of non-covalent interactions between the QM and MM regions and. (ii) where covalent bonds exist between atoms in the QM and MM regions, the treatment of bonds at the frontier between the two regions.")
29
Advantages of QM The QM treatment of the electronic structure of a small active site region allows chemical reactions to be modeled, including the effects of the environment through an empirical MM treatment. Enzyme mechanisms can be tested, transition states (TSs) and reactive species identified and characterised and the effects of mutations investigated. The dynamics of proteins can also be simulated, identifying conformational changes that may be associated with reaction.
 and reactive species identified and characterised and the effects of mutations investigated. The dynamics of proteins can also be simulated, identifying conformational changes that may be associated with reaction.")
30
oxidative deamination of tryptamine
The rate-limiting proton transfer step in the oxidative deamination of tryptamine by aromatic amine dehydrogenase (AADH). A primary H/D KIE of 55 ± 6 has been reported for this step from experimental studies, one of the highest KIEs reported for an enzyme-catalysed proton transfer . Calculations indicate that quantum tunnelling is important in this reaction, but find no role for large-scale protein dynamics in driving the reaction. John D. McGeagh, Kara E. Ranaghan, Adrian J. Mulholland, Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics. Volume 1814, Issue 8, August 2011, Pages 1077–1092
. A primary H/D KIE of 55 ± 6 has been reported for this step from experimental studies, one of the highest KIEs reported for an enzyme-catalysed proton transfer . Calculations indicate that quantum tunnelling is important in this reaction, but find no role for large-scale protein dynamics in driving the reaction. John D. McGeagh, Kara E. Ranaghan, Adrian J. Mulholland, Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics. Volume 1814, Issue 8, August 2011, Pages 1077–1092.")
31
Question: Which animal likes baseball the most?
A bat of course……………!

32
Combined QM/MM methods Goal: to do quantum chemical calculations of reactions and electronically excited states of large molecular systems. QM 24-40 Å MM Heff = Hqm + Hqm/mm + Hmm

33
Combined QM/MM Simulation
Generalized Hybrid Orbital (GHO) for the treatment of QM and MM boundary Balancing Accuracy and Applicability QM models can be systematically improved. Semiempirical QM models can be parameterized for specific reactions and properties. MM force fields can be applied to large systems. Bond-Making and Breaking Processes Electronic polarization by the dynamics of the environment is naturally included.
 for the treatment of QM and MM boundary. Balancing Accuracy and Applicability. QM models can be systematically improved. Semiempirical QM models can be parameterized for specific reactions and properties. MM force fields can be applied to large systems. Bond-Making and Breaking Processes. Electronic polarization by the dynamics of the environment is naturally included.")
34
Model Reaction Strategy
Alanine Phenoxide ion AlaR Ala-PLP(H+) Ala-PLP
 Ala-PLP.")
35
ENZYMES The activity of a cell is largely controlled by the activity of the network of enzymes within it. Monod (1972) describes networks of enzymes as 'microscopic cybernetics'. They are a clear-cut example of adaptive computation occuring on a subcellular level. Enzymes are, in most cases, incredibly specific. They are also orders of magnitude more powerful than non-organic catalysts. Networks of enzyme-mediated reactions are not simply a matter of enzymes catalysing the production of other enzymes. There are additional multiple feedback and feedforward effects. For example, enzymes can be activated by the degradation of their metabolites. This will tend to stabilise the level of the metabolites in the system. Enzymes may also be activated by the metabolites of enzymes in another sequence. Allosteric enzymes inhibit and/ or enhance the efficacy of other enzymes, as well as acting as catalysts themselves.
 describes networks of enzymes as microscopic cybernetics . They are a clear-cut example of adaptive computation occuring on a subcellular level. Enzymes are, in most cases, incredibly specific. They are also orders of magnitude more powerful than non-organic catalysts. Networks of enzyme-mediated reactions are not simply a matter of enzymes catalysing the production of other enzymes. There are additional multiple feedback and feedforward effects. For example, enzymes can be activated by the degradation of their metabolites. This will tend to stabilise the level of the metabolites in the system. Enzymes may also be activated by the metabolites of enzymes in another sequence. Allosteric enzymes inhibit and/ or enhance the efficacy of other enzymes, as well as acting as catalysts themselves.")
36
ENZYMES & QUANTUM TUNNELING
At least some enzymes catalyse using quantum tunneling. In actuality, quantum tunneling may be the norm in enzyme catalysis: Hydrogen-transfer processes are expected to show appreciable quantum mechanical behaviour. Intensive investigations of enzymes under their physiological conditions show this to be true of practically every example investigated. (Klinman 2003). Quantum tunneling reduces the amount of energy required to catalyze a reaction. There would therefore be strong evolutionary pressures this more energy-efficient catalysis to develop. There is a huge literature on this subject, most of it very technical. Redox chains are chains of electron transfer (by quantum tunneling) which occur as part of actual catalytic process. Redox chains can intersect at nodes called redox clusters. (See Moser, Page, Chen and Dutton 2000 for a discussion of redox chains). Maps of redox chains within the catalytic process display a similar structure (nodes; multiple linkages) to maps of chains of catalytic reactions.
. Quantum tunneling reduces the amount of energy required to catalyze a reaction. There would therefore be strong evolutionary pressures this more energy-efficient catalysis to develop. There is a huge literature on this subject, most of it very technical. Redox chains are chains of electron transfer (by quantum tunneling) which occur as part of actual catalytic process. Redox chains can intersect at nodes called redox clusters. (See Moser, Page, Chen and Dutton 2000 for a discussion of redox chains). Maps of redox chains within the catalytic process display a similar structure (nodes; multiple linkages) to maps of chains of catalytic reactions.")
37
Enzymes as proteins

38
Cofactors Cofactors (non-protein component of an enzyme):
Some enzymes do not need any additional components to show full activity. However, others require non-protein molecules called cofactors to be bound for activity. Cofactors can be either inorganic ( e.g. , metal ions e.g., Mn+2, Zn+2, Fe+2, Ca+2 and iron-sulfur clusters) or organic compounds, (e.g., flavin and heme). Organic compounds (Posthetic group/Co-enzymes): Prosthetic groups, which are tightly bound to an enzyme, or coenzymes, which are released from the enzyme's active site during the reaction. Coenzymes include NADH, NADPH and adenosine triphosphate . These molecules act to transfer chemical groups between enzymes. carbonic anhydrase, with a zinc cofactor bound as part of its active site. These loosely-bound molecules are usually found in the active site and are involved in catalysis but not released from the active site. For example, flavin and heme cofactors are often involved in redox reactions. Enzymes that require a cofactor but do not have one bound are called apoenzymes . An apoenzyme together with its cofactor(s) is called a holoenzyme (this is the active form). Most cofactors are not covalently attached to an enzyme, but are very tightly bound. However, organic prosthetic groups can be covalently bound.
 or organic compounds, (e.g., flavin and heme). Organic compounds (Posthetic group/Co-enzymes): Prosthetic groups, which are tightly bound to an enzyme, or coenzymes, which are released from the enzyme s active site during the reaction. Coenzymes include NADH, NADPH and adenosine triphosphate . These molecules act to transfer chemical groups between enzymes. carbonic anhydrase, with a zinc cofactor bound as part of its active site. These loosely-bound molecules are usually found in the active site and are involved in catalysis but not released from the active site. For example, flavin and heme cofactors are often involved in redox reactions. Enzymes that require a cofactor but do not have one bound are called apoenzymes . An apoenzyme together with its cofactor(s) is called a holoenzyme (this is the active form). Most cofactors are not covalently attached to an enzyme, but are very tightly bound. However, organic prosthetic groups can be covalently bound.")
39
Co-enzymes Coenzymes :
Coenzymes Coenzymes are small organic molecules that transport chemical groups from one enzyme to another. Some of these chemicals such as riboflavin, thiamine and folic acid are vitamins, (acquired). The chemical groups carried include the hydride ion (H - ) carried by NAD or NADP + , the acetyl group carried by coenzyme A , … etc. Since coenzymes are chemically changed as a consequence of enzyme action, it is useful to consider coenzymes to be a special class of substrates, or second substrates, which are common to many different enzymes. For example, about 700 enzymes are known to use the coenzyme NADH. Coenzymes are usually regenerated and their concentrations maintained at a steady level inside the cell: for example, NADPH is regenerated through the pentose phosphate pathway and S- adenosylmethionine by methionine adenosyltransferase .
. The chemical groups carried include the hydride ion (H - ) carried by NAD or NADP + , the acetyl group carried by coenzyme A , … etc. Since coenzymes are chemically changed as a consequence of enzyme action, it is useful to consider coenzymes to be a special class of substrates, or second substrates, which are common to many different enzymes. For example, about 700 enzymes are known to use the coenzyme NADH. Coenzymes are usually regenerated and their concentrations maintained at a steady level inside the cell: for example, NADPH is regenerated through the pentose phosphate pathway and S- adenosylmethionine by methionine adenosyltransferase .")
40
Recap prosthetic group – the non-amino acid part of a conjugated protein. cofactor - a nonprotein molecule or ion required by an enzyme for catalytic activity - can be an organic molecule or metal ion, such as Mg2+, Zn2+, Fe2+, Ca2+ coenzyme - an organic cofactor apoenzyme - a catalytically inactive protein (formed by removal of the cofactor from the active enzyme). - activated by a cofactor apoenzyme + cofactor(coenzyme or inorganic ion) --> active enzyme An important group of coenzymes are vitamins. For example, vitamin B12 acts as coenzyme B12 in the shift of H atoms between adjacent carbon atoms and in the transfer of methyl groups. Another example, niacin acts as nicotinamide adenine dinucleotide (NAD+) in hydrogen transfer. Another important group of cofactors are minerals in our diet. For example, the coenzyme Zn2+ is required for the enzyme carbonic anhydrase to function.
. - activated by a cofactor. apoenzyme + cofactor(coenzyme or inorganic ion) --> active enzyme. An important group of coenzymes are vitamins. For example, vitamin B12 acts as coenzyme B12 in the shift of H atoms between adjacent carbon atoms and in the transfer of methyl groups. Another example, niacin acts as nicotinamide adenine dinucleotide (NAD+) in hydrogen transfer. Another important group of cofactors are minerals in our diet. For example, the coenzyme Zn2+ is required for the enzyme carbonic anhydrase to function.")
41
Learning Check E2 The active site is (1) the enzyme
(2) a section of the enzyme (3) the substrate B. In the induced fit model, the shape of the enzyme when substrate binds (1) Stays the same (2) adapts to the shape of the substrate
 a section of the enzyme. (3) the substrate. B. In the induced fit model, the shape of the enzyme when substrate binds. (1) Stays the same. (2) adapts to the shape of the substrate.")
42
Learning Check E3 C. In the transition state model, the enzyme’s reactive stage can be defined as: (1) Staying the same. (2) Changing before binding with substrate. (3) Changing after binding with substrate. (4) Changing during binding with substrate. D. In the quantum tunneling model, the enzyme tends to overcome: (1) Substrate . (2) Energy deficit. (3) A gradient. (4) Climb a hill.
 Staying the same. (2) Changing before binding with substrate. (3) Changing after binding with substrate. (4) Changing during binding with substrate. D. In the quantum tunneling model, the enzyme tends to overcome: (1) Substrate . (2) Energy deficit. (3) A gradient. (4) Climb a hill.")
43
Solution E2, E3 The active site is (2) a section of the enzyme
B. In the induced fit model, the shape of the enzyme when substrate binds (2) adapts to the shape of the substrate C. In the transition state model, the enzyme’s reactive stage can be defined as: (4) Changing during binding with substrate. D. In the quantum tunneling model, the enzyme tends to overcome: (2) Energy deficit.
 adapts to the shape of the substrate. C. In the transition state model, the enzyme’s reactive stage can be defined as: (4) Changing during binding with substrate. D. In the quantum tunneling model, the enzyme tends to overcome: (2) Energy deficit.")
Similar presentations



















.>")