Download presentation
Presentation is loading. Please wait.

1
Epigenetics: Modification of DNA and chromatin to control DNA structure and Gene Expression

2
Genetic Variation is a Change in the DNA Sequence
SNPs Indels Rearrangements Mendelian Inheritance

3
X

4
Epigenetics relies on DNA methylation
Epigenetics - heritable change in phenotype without a change in DNA sequence Organismal or cell generations A mechanism involved in epigenetic phenotype regulation DNA methylation is a transcription (OFF) switch Histone methylation is a transcription (OFF) switch Histone acetylation is a transcription (ON) switch Will also involve other molecular mechanism
 switch. Histone methylation is a transcription (OFF) switch. Histone acetylation is a transcription (ON) switch. Will also involve other molecular mechanism.")
5
DNA methylation in different species
S. cerevisiae Arabidopsis C. elegans Drosophila Vertebrates S. pombe N. crassa DNA methylation — — + — (+) + + DNA methylation binding proteins — + + + + + + Imprinting — — — — (+) + + A. Peaston TJL
 + + DNA. methylation. binding. proteins. — Imprinting. — — — — (+) + + A. Peaston TJL.")
6
DNA methylation -- 5'CpG3' cytosine guanine H N O C1' CH3 DNMT
A. Peaston TJL

7
Establishment of DNA methylation
7 5 10 15 20 Days post conception Relative methylation at CpG sites Establishment of DNA methylation ~ 70% Yoder, Walsh & Bestor, 1997 Trends Genet

8
Establishment of DNA methylation: imprinting
8 Concept – CpG islands are a special case: inactive X-chromosome is heavily CpG island methylated, whereas autosomes and active X are generally unmethylated. Concept – in the developing germ cells of male embryos, the female-derived unmethylated alleles of imprinted genes e.g. H19 become methylated, thus the mature male will pass on the male imprint to the next generation via his germ cells whilst his somatic cells will retain both the maternal and paternal allele-specific methylation. Similarly, in developing germ cells of female embryos, the male-derived methylated alleles of imprinted genes will become unmethylated, and the female will thus pass only female-pattern methylation of those alleles to her offspring although her somatic cells will retain both maternal and paternal allele-specific methylation. 8 Establishment of DNA methylation: imprinting 5 10 15 20 Days post conception Relative methylation at CpG sites H19 Imprinted gene Maternal allele in male germ cells Paternal allele in female germ cells Inactive X Autosomes & active X CpG islands

9
DNA methylation and cell memory
Self-complementary CpG dinucleotides DNA replication Maintenance methylation Holliday and Pugh 1975 Science Riggs 1975 Cytogenet Cell Genet

10
Law & Jacobsen Nat Rev. Genet 2010

11
DNA demethylation Passive Active e.g. zygotic paternal genome,
IL2 promoter A. Peaston TJL

12
DNA demethylation -- CpG islands
OFF ? CpG methylation default, islands exclude global activity ? A. Peaston TJL

13
Law & Jacobsen Nat Rev. Genet 2010

14
DNA Demethylation and 5hydroxymethyl cytosine
5-methylcytosine oxidized to 5 hydroxymethylcytosine Tet family (ten-eleven translocation)
")
15
Wu & Zhang 2011 Cell Cycle

16
How does DNA methylation inhibit transcription?
Titratable transcriptional inhibition DNA-protein complex specific for methylated DNA MeCP2, MBD1-4, Kaiso Methyl-binding proteins recruit different co-repressor complexes HDAC Sin3A HKMT MeCP2 Me me A. Peaston TJL

17
How does DNA methylation inhibit transcription?
Biochemical experiments using artificial CpG methylation & transfection re-expression of X-linked genes after 5-azacytidine treatment Interference with transcription factor or other protein binding e.g. CTCF binding Mat Pat Igf2 ICE CTCF E H19 me A. Peaston TJL

18
Assessing DNA methylation
Avy/a Morgan et al, Nature Genetics 1999

19
DNA methylation at the agouti locus
Avy allele Hypothesis IAP OFF LTR CpG methylated pseudoagouti LTR CpG unmethylated IAP ON yellow

20
Methylation-sensitive restriction enzymes
Avy allele B BamH1 M Msp1 H HpaII Isoschizomers, C/CGG A. Peaston TJL

21
Bisulfite sequencing Locus-specific detailed single-molecule analysis
5MeCpG CpG bisulfite DNA PCR UG CG TG Frommer et al, PNAS 1992

22
Bisulfite Sequencing Sperm 95% Oocytes 98% Zygotes 90% Blastocyst 62%
Lane et al, Genesis 2003

23
Higher throughput Pyrosequencing of clones (You still have to clone)
SAGE

24
Next Generation Sequencing
Higher throughput Next Generation Sequencing High throughput, non-biological cloning

25
And permutations..... Murrell, Rakyan & Beck, Hum Mol Genet 2005

26
Chromatin and DNA methylation
Neurospora and Arabidopsis – methylation depends on H3K9me Plants – RNAi targets chromatin modification, gene silencing & DNAme Mouse reduced CpG methylation with : deficiency of H3K9 methyltransferases deficiency of EZH2 (H3K27 methyltransferase) increased CpG methylation with : antisense transcription through promoters unsettled role of RNAi Many species – link to SWI/SNF-like remodeling proteins
 increased CpG methylation with : antisense transcription through promoters. unsettled role of RNAi. Many species – link to SWI/SNF-like remodeling proteins.")
27
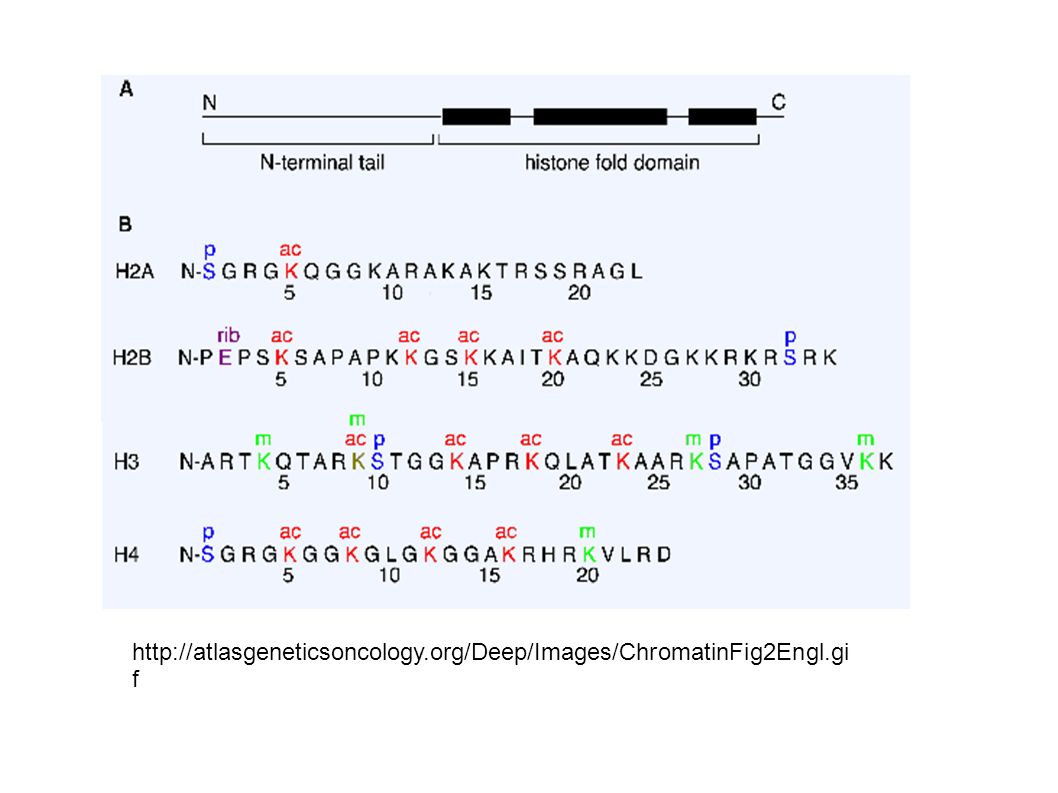
The nucleosome core. Site of action
The nucleosome core. Site of action. Tails that get modified are evident in this image.

29
Histone modifying enzymes
2929 Histone modifying enzymes often are part of large multi-subunit complexes. Many modifying enzymes have cell context-dependent substrate-specificity often with other substrates as well as histones. Histone modifications provide marks recognized by specific domains e.g. bromo and chromo domains . Bromo domain recognition sites are recognized by chromatin remodelling proteins of the Trithorax group, whilst chromo domains are recognized by at least 5 related families of proteins including the polycomb family of proteins. An additional level of complexity – several trithorax group proteins are able to perform more than one type of modificatin reaction e.g. some trithorax proteins methylate H3K4 a mark of active gene expression, as well as performing nucleosome remodelling activity. Also, the histone modfying enzymes provide a link with cellular response to external stimuli. E.g. Mitch Smith, Hbo1 HAT (histone acetyl transferase). Under physiologic stresses that stabilize p53 (in yeast), P53 binds Hbo1 protein and represses its activity. Moreover, Hbo1 also has documented roles in transcription and DNA replication. Histone modifying enzymes 29
. Under physiologic stresses that stabilize p53 (in yeast), P53 binds Hbo1 protein and represses its activity. Moreover, Hbo1 also has documented roles in transcription and DNA replication. Histone modifying enzymes. 29.")
30
Chromatin remodelling – Polycomb and Trithorax
3030 Example: recruiting ATP-dependent remodelling complexes such as Swi-snf group (Trithorax group) remodel chromatin by by nucleosome octamer sliding, altering nucleosome structure by DNA looping or replacement of core histones with histone variants Polycomb complexes recognize and promote repressive histone methylation and promote tight packing of nucleosomes. Note interactions btw two systems. Illustration in next slide. Chromatin remodelling – Polycomb and Trithorax 30
 remodel chromatin by. by nucleosome octamer sliding, altering nucleosome structure by DNA looping. or replacement of core histones with histone variants. Polycomb complexes recognize and promote repressive histone methylation and promote tight packing of nucleosomes. Note interactions btw two systems. Illustration in next slide. Chromatin remodelling – Polycomb and Trithorax. 30.")
31
3131 Concept: non-coding RNAs including for example Xist (large) and small RNAs directly transcribed fom genome or from double-stranded RNA degradation (RNA interference) also regulate epigenetic DNA modifications in ways that are only just beginning to be understood. e.g. slide illustrates that RNAi guides histone modification during S-phase. Fission yeast Swi6 = mammalian HP1. In G2, heterochromatic repeats have high levels of Swi6 and H3K9me2 and retain cohesin. In M, phosphorylation of H3S10 causes loss of Swi6 & transcription of the repeats in early S. Also H3K9me2 levels fall during replication, but are restored during S, guided by RNAi of the repeat transcripts and accompanied by recruitment of Swi6. 31 Kloc et al Current Biology 2008
 and small RNAs directly transcribed fom genome or from double-stranded RNA degradation (RNA interference) also regulate epigenetic DNA modifications in ways that are only just beginning to be understood. e.g. slide illustrates that RNAi guides histone modification during S-phase. Fission yeast Swi6 = mammalian HP1. In G2, heterochromatic repeats have high levels of Swi6 and H3K9me2 and retain cohesin. In M, phosphorylation of H3S10 causes loss of Swi6 & transcription of the repeats in early S. Also H3K9me2 levels fall during replication, but are restored during S, guided by RNAi of the repeat transcripts and accompanied by recruitment of Swi Kloc et al Current Biology")
32
Epigenetic changes and phenotype
It is not just DNA methylation that leads to various pathologic phenotypes, but abnormalities of DNA methylation have certainly been linked with these pathologic processes, in some cases as cause, in many cases it is not yet clear. 3232 Epigenetic changes and phenotype DNA methylation and mutation Genomic instability Cancer Complex disease Environmental effects on DNA methylation A. Peaston TJL

33
Epigenetics/Epigenomics
Modification of histones Mostly Acetylation (+) and Methylation (-) Histone acetyl transferases Histone deacetylases Modification of DNA In eukaryotes, mostly 5-methylcytosine (-) Dnmt (DNA methyl transferases) Removal? A sixth base; 5-hydroxymethylcytosine
 and Methylation (-) Histone acetyl transferases. Histone deacetylases. Modification of DNA. In eukaryotes, mostly 5-methylcytosine (-) Dnmt (DNA methyl transferases) Removal A sixth base; 5-hydroxymethylcytosine.")
34
Epigenetics/Epigenomics
Methylated DNA recruits HDACs Increased condensation of chromatin decreased expression Methylated chromatin may recruit Dnmts Transcription factors may recruit Dnmts, HDACs or HMTs or HATs depending on function Signals can propagate along the chromosome using bi-functional enzymes (chromodomain and HMT activity)
")
Similar presentations



 Neurochemistry Lab.>")

 DNA (gene) mRNA Protein Transcription RNA processing (splicing etc) Translation Folding Post translational modifications Peptides/amino.>")







 Wrap-up RNAi Overview and Discussion Preview: Regulating Gene Expression Exam Return.>")





 DNA (- charge) and histones (+>")
