Download presentation
Presentation is loading. Please wait.

1
Lipid metabolism You give me fever Peggy lee

2
FATTY ACID OXIDATION β-oxidation is the pathway by which the fatty acids from both dietary fat and adipose tissue TAG are oxidized to acetyl-CoA. The reducing equivalents released during fatty acid oxidation are captured in the form of FADH2 and NADH, which are used to support oxidative phosphorylation In most circumstances, the acetyl-CoA units generated by β -oxidation will subsequently be oxidized through the TCA cycle, generating additional FADH2 and NADH and ultimately, additional ATP All cells and tissues except red blood cells and the brain oxidize fatty acids to generate ATP The β-oxidation pathway is absent in red blood cells because they lack mitochondria Although neuronal cells in the brain do contain mitochondria, there is only limited transport of fatty acids across the blood-brain barrier

3
The skeletal and heart muscles in particular have a large capacity for oxidizing fatty acids
Normally, 60 to 90% of the energy required for contraction of the heart is derived from the oxidation of fatty acids In the fasted state, many cells and tissues depend on β -oxidation of fatty acids to provide the ATP needed to maintain ion gradients and to support biosynthetic processes such as gluconeogenesis During prolonged fasting or starvation the brain meets its energy needs by oxidizing ketone bodies as well as glucose The ketone bodies of physiological significance are four-carbon anions produced from acetyl-CoA generated by the β-oxidation of long-chain fatty acids in the liver Oxidation of ketone bodies by the brain reduces the brain’s dependence on glucose and thus decreases the body’s need to catabolize muscle proteins for gluconeogenesis

4
The major fatty acids of both dietary TAG and adipose stores in the body contain 16 to 18 carbons and are oxidized through mitochondrial β-oxidation Essentially the same pathway is utilized for short- (C4-C6) and medium-chain (C8-C12) fatty acids Related peroxisomal pathways oxidize the less common branched-chain and very long-chain (>C22) fatty acids (VLCFA) Transport and Activation of Fatty Acids Since long-chain fatty acids are poorly soluble in aqueous media, they must be transported in the plasma complexed with albumin When a fatty acid dissociates from albumin it is transferred from the capillary lumen through the capillary endothelium and interstitial space to the cells below Long-chain fatty acids enter cells both by simple diffusion and by carrier-mediated transport
 and medium-chain (C8-C12) fatty acids. Related peroxisomal pathways oxidize the less common branched-chain and very long-chain (>C22) fatty acids (VLCFA) Transport and Activation of Fatty Acids. Since long-chain fatty acids are poorly soluble in aqueous media, they must be transported in the plasma complexed with albumin. When a fatty acid dissociates from albumin it is transferred from the capillary lumen through the capillary endothelium and interstitial space to the cells below. Long-chain fatty acids enter cells both by simple diffusion and by carrier-mediated transport.")
5
Intracellularly, fatty acids are bound to cytosolic fatty acid binding proteins (FABP), which deliver the fatty acids to the sites where they are metabolized Once inside cells, fatty acids must be activated before they can be metabolized In contrast to glucose, which is activated and trapped within cells as glucose 6-phosphate,fatty acids are converted not to acyl phosphates but to thioesters of coenzyme A Long-chain fatty acids destined for β-oxidation are activated to their CoA forms primarily on the surface of the outer mitochondrial membrane The inner mitochondrial membrane is, however, impermeable to long-chain fatty acyl-CoA molecules Transport of LCFA across the inner mitochondrial membrane is facilitated by a fatty acid transport mechanism called the carnitine shuttle

6
Carnitine is a quaternary amine that has a hydroxyl group to which a fatty acid can be attached
Since carnitine can be synthesized in the liver and kidney, it is not usually considered an essential dietary nutrient The activity of the carnitine translocase system is dependent on two enzymes, carnitine palmitoyltransferase I (CPT-I) and carnitine palmitoyltransferase II (CPT-II), both of which catalyze the reversible transfer of long-chain fatty acids between CoA and carnitine CPT-I is localized to the mitochondrial outer membrane and acts to generate acylcarnitine Carnitine translocase, which is embedded in the inner mitochondrial membrane transports acylcarnitine into the mitochondria in exchange for free carnitine, which is concurrently exported from the mitochondrial matrix into the cytosol
 and carnitine palmitoyltransferase II (CPT-II), both of which catalyze the reversible transfer of long-chain fatty acids between CoA and carnitine. CPT-I is localized to the mitochondrial outer membrane and acts to generate acylcarnitine. Carnitine translocase, which is embedded in the inner mitochondrial membrane transports acylcarnitine into the mitochondria in exchange for free carnitine, which is concurrently exported from the mitochondrial matrix into the cytosol.")
7
CPT-II, which is localized to the matrix face of the inner mitochondrial membrane, forms mitochondrial acyl-CoA from CoASH and acylcarnitine The net effect is the transfer of a long-chain fatty acid from an cytosolic CoASH to a mitochondrial CoASH Intramitochondrial β-Oxidation The term β-oxidation is derived from the fact that the critical chemistry of the four core reactions that comprise the pathway takes place on the third carbon from the carboxyl end: that is, the β-carbon atom The first of the four reactions of the β-oxidation pathway is irreversible and is catalyzed by acyl-CoA dehydrogenase Two hydrogen atoms are removed-one each from the α and β and -generating a carbon-carbon double bond between the α and β carbons These hydrogen atoms are transferred to FAD to form FADH2

8
The Mechanism of The Carnitine
Shuttle

9
Mitochondria contain a family of FAD-linked acyl-CoA dehydrogenases: a long-chain acyl-CoA dehydrogenase, a medium-chain acyl-CoA dehydrogenase and a short-chain acyl-CoA dehydrogenase The second step in β-oxidation involves hydration of the carbon-carbon double bond between by enoyl-CoA hydratase. The hydroxyl group is introduced onto the β-carbon In the third step, a second dehydrogenase, NAD +-linked β-hydroxyacyl-CoA dehydrogenase, oxidizes the hydroxyacyl-CoA molecule to generate a β-ketoacyl-CoA and a molecule of NADH The fourth and final step in β-oxidation involves cleavage of the fatty acid chain with attachment of a second molecule of CoASH to the β-carbon and generation of one molecule of acetyl-CoA The enzyme that catalyzes this reaction is called β-ketoacyl-CoA thiolase

10
The Reactions of β-Oxidation

11
The net effect of the four steps in β-oxidation is the production of one molecule of acetyl-CoA and one fatty acyl-CoA molecule whose carbon chain is two carbons shorter than the original substrate The four steps are then repeated, with successive chain shortening by two carbon atoms The final thiolytic cleavage reaction converts the 4-carbon β -ketobutyryl-CoA (acetoacetyl-CoA) into two molecules of acetyl-CoA The Energy Yield from β-oxidation A cycle of β-oxidation releases one molecule each of FADH2, NADH and acetyl-CoA The four steps are repeated n/ times ( n = number of carbons) until the final acetyl-CoA is released n/2 acetyl-CoA and n/ each of FADH2 and NADH are produced by the complete oxidation of a fatty acid with an even number of carbons
 into two molecules of acetyl-CoA. The Energy Yield from β-oxidation. A cycle of β-oxidation releases one molecule each of FADH2, NADH and acetyl-CoA. The four steps are repeated n/2 - 1 times ( n = number of carbons) until the final acetyl-CoA is released. n/2 acetyl-CoA and n/2 - 1 each of FADH2 and NADH are produced by the complete oxidation of a fatty acid with an even number of carbons.")
12
Taking the oxidation of palmitate :
Palmitoyl-CoA+ 7 CoA+ 7 FAD+ 7 NAD++7 H2O→ 8 acetyl-CoA+ 7 FADH2+7 NADH+ 7H+ 8 acetyl-CoA ≈ 8o ATP, 7 FADH2 ≈ 10.5 ATP and 7 NADH ≈ 17.5 ATP → total = 108 ATP The equivalent of two ATP molecules is spent on the activation of fatty acids; this means a net production of 106 ATP Ancillary Reactions to the Pathway of β-Oxidation β –oxidation of saturated fatty acids generates an unsaturated intermediate with a Δ2,3 -trans double bond which is then hydrated by enoyl-CoA hydratase The metabolism of unsaturated and polyunsaturated fatty acids requires additional enzymes to act on preexisting cis double bonds For example, the oxidation of linoleic acid proceeds in essentially the same manner as the β-oxidation of saturated fatty acids

13
However, after three cycles of β-oxidation, the chain- shortening process produces an acyl-CoA with a cis-3,4 double bond At this point, an additional enzyme, Δ 3, Δ 2 -enoyl-CoA isomerase, converts the Δ 3,4-cis double bond to a Δ 2,3-trans double bond, thus providing a suitable substrate for enoyl-CoA hydratase A slightly different situation arises when the chain-shortening process produces an acyl-CoA molecule that has a cis-4,5 double bond Under these conditions, the acyl-CoA dehydrogenase step generates a Δ 2,3-trans /Δ 4,5-cis conjugated di-unsaturated fatty acyl-CoA intermediate At this point another enzyme, NADPH-dependent 2,4-dienoyl-CoA reductase, transfers hydrogen atoms from NADPH to carbons 4 and 5, generating a Δ 3,4-enoyl-CoA. Δ 3, Δ 2 -enoyl-CoA isomerase then converts Δ3,4-trans-enoyl-CoA to Δ2, 3 -trans-enoyl-CoA

14
The Role of Auxiliary Enzymes in the β-Oxidation of Linoleic Acid

15
Oxidation of Medium-Chain Fatty Acids
Cow’s milk contains relatively large amounts of medium-chain fatty acids while long-chain fatty acids (particularly palmitic, oleic, and linoleic acids) predominate in breast milk These shorter fatty acids are more soluble than are their more common C16-C20 counterparts and can enter the mitochondrion directly from the cytosol without need for the carnitine transporter system The C8-C 12 fatty acids are activated to their corresponding acyl-CoA derivatives within the mitochondrion and then undergo β-oxidation The initial oxidation reaction is catalyzed by medium-chain acyl-CoA dehydrogenase (MCAD) Oxidation of Fatty Acids With an Odd-Number Of Carbons Dietary lipids often contain a small amount of odd-carbon fatty acids such as 17:0
 predominate in breast milk. These shorter fatty acids are more soluble than are their more common C16-C20 counterparts and can enter the mitochondrion directly from the cytosol without need for the carnitine transporter system. The C8-C 12 fatty acids are activated to their corresponding acyl-CoA derivatives within the mitochondrion and then undergo β-oxidation. The initial oxidation reaction is catalyzed by medium-chain acyl-CoA dehydrogenase (MCAD) Oxidation of Fatty Acids With an Odd-Number Of Carbons. Dietary lipids often contain a small amount of odd-carbon fatty acids such as 17:0.")
16
Odd-chain fatty acids also undergo β-oxidation
However, the last thiolytic cleavage step produces one molecule of acetyl-CoA and one molecule of propionyl-CoA Carboxylation of propionyl CoA yields methylmalonyl CoA, which is ultimately converted to succinyl CoA in a vitamin B12–dependent reaction Propionyl CoA also arises from the oxidation of branched chain amino acids The propionyl CoA to succinyl CoA pathway is a major anaplerotic route for the TCA cycle Thus, this small proportion of the odd-carbon number fatty acid chain can be converted to glucose In contrast, the acetyl-CoA formed from β-oxidation of even-chain-number (and odd-chain) fatty acids in the liver either enters the TCA cycle or is converted to ketone bodies
 fatty acids in the liver either enters the TCA cycle or is converted to ketone bodies.")
17
The Metabolism of Propionyl-CoA
Ketone Bodies The association of ketone bodies with the ketoacidosis of diabetes mellitus has given these substances a bad reputation However, ketone bodies are normal metabolites that serve as circulating fuels, especially during periods of moderate (12 to 24 hours) or severe (>5 days) fasting The two physiologically significant ketones are acetoacetate (β –ketobutyrate) and β –hydroxybutyrate Acetone is the product of the non-enzymatic decarboxylation of acetoacetate The Metabolism of Propionyl-CoA
 or severe (>5 days) fasting. The two physiologically significant ketones are acetoacetate (β –ketobutyrate) and β –hydroxybutyrate. Acetone is the product of the non-enzymatic decarboxylation of acetoacetate. The Metabolism of Propionyl-CoA.")
18
Unlike hydrophobic long-chain fatty acids that require albumin for their transport in the plasma, ketone bodies are water-soluble and do not require a carrier protein for transport Ketone bodies can hence be thought of as easily transportable forms of fatty acids Ketone bodies are synthesized mainly in the liver with a smaller contribution from the renal cortex In both tissues, the substrate for ketogenesis is mitochondrial acetyl-CoA, which is derived from β-oxidation and to a lesser extent, from the oxidation of ketogenic amino acids (e.g., leucine) In the fasted state, much of the acetyl-CoA generated by β-oxidation cannot enter the TCA cycle because of a relative shortage of oxaloacetate which has been diverted to gluconeogenesis
 In the fasted state, much of the acetyl-CoA generated by β-oxidation cannot enter the TCA cycle because of a relative shortage of oxaloacetate which has been diverted to gluconeogenesis.")
19
2Acetyl-CoA⇌acetoacetyl-CoA + CoA
The pathway of ketone body synthesis converts two acetyl-CoA molecules into one four-carbon acetoacetate molecule while releasing two free CoASH molecules, which are required for continued β-oxidation Continued β-oxidation, in turn, provides FADH2 and NADH for oxidative phosphorylation The first step in acetoacetate synthesis is catalyzed by β -ketothiolase, which also catalyzes the last step in β-oxidation 2Acetyl-CoA⇌acetoacetyl-CoA + CoA This reversible reaction is driven to the right by a high concentration of acetyl-CoA arising from β-oxidation The acetoacetyl-CoA is then combined with a third molecule of acetyl-CoA to form β-hydroxy- β -methylglutaryl-CoA (HMG-CoA) in a reaction catalyzed by HMG-CoA synthase Most cells contain a second HMG-CoA synthase that is localized to the cytosol, where it is involved in cholesterol synthesis
 in a reaction catalyzed by HMG-CoA synthase. Most cells contain a second HMG-CoA synthase that is localized to the cytosol, where it is involved in cholesterol synthesis.")
20
Acetoacetate + NADH + H+ → β-hydroxybutyrate + NAD+
Mitochondrial HMG-CoA is then hydrolyzed by HMG-CoA lyase to produce acetoacetate plus acetyl-CoA While about one-third of the acetoacetate produced by HMG-CoA lyase is secreted by the liver into the circulation, the other two-thirds is first reduced by mitochondrial β-hydroxybutyrate dehydrogenase and then secreted: Acetoacetate + NADH + H+ → β-hydroxybutyrate + NAD+ This reaction is driven in the direction of β-hydroxybutyrate synthesis by the relatively high mitochondrial ratio of NADH/NAD+ generated by active β-oxidation of fatty acids β -Hydroxybutyrate is more reduced and more energy-rich than acetoacetate Utilization of Ketone Bodies Although the liver does not oxidize ketone bodies, the heart and skeletal muscle are capable of efficiently oxidizing ketones bodies

21
Ketogenesis

22
Ketone utilization is initiated by mitochondrial β –hydroxy-butyrate dehydrogenase, which converts β-hydroxybutyrate back into acetoacetate Acetoacetate is then activated (and trapped within the cell) by one of two mitochondrial enzymatic reactions The first trapping reaction is reversible and catalyzed by succinyl-CoA: β -ketoacid CoA-transferase: The other trapping reaction is catalyzed by acetoacetyl-CoA synthetase β-Ketoacid CoA-transferase and acetoacetyl-CoA synthetase are both absent from hepatocytes, which accounts for the inability of liver to oxidize ketone bodies Acetoacetyl-CoA is then cleaved by β-ketothiolase into two molecules of acetyl- CoA. Since tissues such as muscle that oxidize ketone bodies do not perform gluconeogenesis and thus do not deplete their supply of oxaloacetate in the fasted state, the acetyl-CoA molecules join the TCA cycle
 by one of two mitochondrial enzymatic reactions. The first trapping reaction is reversible and catalyzed by succinyl-CoA: β -ketoacid CoA-transferase: The other trapping reaction is catalyzed by acetoacetyl-CoA synthetase. β-Ketoacid CoA-transferase and acetoacetyl-CoA synthetase are both absent from hepatocytes, which accounts for the inability of liver to oxidize ketone bodies. Acetoacetyl-CoA is then cleaved by β-ketothiolase into two molecules of acetyl- CoA. Since tissues such as muscle that oxidize ketone bodies do not perform gluconeogenesis and thus do not deplete their supply of oxaloacetate in the fasted state, the acetyl-CoA molecules join the TCA cycle.")
23
Alternate Routes of Fatty Acid Oxidation
Oxidation of Very-Long-Chain Fatty Acids (VLCFA) The initial oxidation of VLCFA comprised of 22 carbon atoms or more is accomplished by a modified β-oxidation pathway that operates in peroxisomes One major difference between the mitochondrial and peroxisomal pathways is that, since peroxisomes lack an electron transport system, the reduced cofactors generated during peroxisomal β-oxidation are not channeled directly into oxidative phosphorylation The Utilization of Ketone Bodies
 The initial oxidation of VLCFA comprised of 22 carbon atoms or more is accomplished by a modified β-oxidation pathway that operates in peroxisomes. One major difference between the mitochondrial and peroxisomal pathways is that, since peroxisomes lack an electron transport system, the reduced cofactors generated during peroxisomal β-oxidation are not channeled directly into oxidative phosphorylation. The Utilization of Ketone Bodies.")
24
The VLCFA are first activated to acyl-CoAs by a distinct acyl-CoA synthase
The first FAD-linked dehydrogenase step in the peroxisomal β-oxidation pathway is different from the corresponding step in standard mitochondrial β-oxidation The peroxisomal FAD-linked dehydrogenase that removes two hydrogen atoms from the fatty acid chain transfers those hydrogens to molecular oxygen, thus producing H2O2 Catalase then breaks down the hydrogen peroxide The subsequent steps of the β-oxidation pathway in peroxisomes are similar to those that operate in mitochondrial β-oxidation The reducing equivalents from the NADH generated by hydroxyacyl-CoA dehydrogenase are utilized for reactions within peroxisomes or shuttled out of the peroxisomes and eventually into mitochondria

25
α-Oxidation of Fatty Acids Containing a Branched Methyl
Once the peroxisomal β-oxidation pathway has reduced the VLCFA chain to the level of an 8- or 10-carbon acyl-CoA molecule, the shortened fatty acid chain is transferred to mitochondria and further catabolized via the mitochondrial β-oxidation pathway The peroxisomal acetyl-CoA units are probably hydrolyzed to acetate, which is subsequently oxidized in mitochondria α-Oxidation of Fatty Acids Containing a Branched Methyl Group The chemistry of the β-oxidation pathway entails removal of both hydrogen atoms from the β-carbon atom Therefore, fatty acids that have a methyl group on C3 (the β -carbon) cannot be oxidized by regular β-oxidation and require a specialized pathway, which is called α-oxidation One such branched-chain fatty acid is phytanic acid, which is derived from the phytanol side chain of chlorophyll
 cannot be oxidized by regular β-oxidation and require a specialized pathway, which is called α-oxidation. One such branched-chain fatty acid is phytanic acid, which is derived from the phytanol side chain of chlorophyll.")
26
Phytanic acid has methyl groups on carbon atoms 3,7, 11, and 15
Humans get most of their phytanic acid from dietary dairy products, beef, and fatty fish Phytanic acid hydroxylase in the peroxisomes introduces a hydroxyl group on the α-carbon, which is then oxidized to a carboxyl group with release of the original carboxyl group as CO2 By shortening the fatty acid by one carbon, the methyl groups will appear on the α-carbon rather than the β-carbon Therefore, the product, pristanoic acid, is a suitable substrate for β-oxidation The remaining CH3 of the fatty acyl-CoA chain are now positioned on even-numbered carbon atoms and therefore do not present a problem for the enzymes of the standard β-oxidation pathway Wherever a methyl group is attached to the α-carbon, cleavage of the carbon chain by β-ketothiolase will generate propionyl-CoA rather than acetyl-CoA

27
Peroxisomal Oxidation of Phytanic Acid

28
ω-Oxidation of Fatty Acids
Fatty acids also may be oxidized at the ω-carbon of the chain (the terminal methyl group) by enzymes in the endoplasmic reticulum; the preferred substrates are C10-C12 fatty acids The ω-methyl group is first oxidized to an alcohol by an enzyme that uses cytochrome P450, molecular oxygen, and NADPH Dehydrogenases convert the alcohol group to a carboxylic acid. The dicarboxylic acids produced by ω-oxidation can undergo β–oxidation, forming compounds with 6 to 10 carbons that are water-soluble Such compounds may be oxidized further, or be excreted in urine as dicarboxylic acids Normally, ω -oxidation is a minor process However, in some conditions that interfere with β-oxidation ω-oxidation produces dicarboxylic acids in increased amounts
 by enzymes in the endoplasmic reticulum; the preferred substrates are C10-C12 fatty acids. The ω-methyl group is first oxidized to an alcohol by an enzyme that uses cytochrome P450, molecular oxygen, and NADPH. Dehydrogenases convert the alcohol group to a carboxylic acid. The dicarboxylic acids produced by ω-oxidation can undergo β–oxidation, forming compounds with 6 to 10 carbons that are water-soluble. Such compounds may be oxidized further, or be excreted in urine as dicarboxylic acids. Normally, ω -oxidation is a minor process. However, in some conditions that interfere with β-oxidation ω-oxidation produces dicarboxylic acids in increased amounts.")
29
ω-Oxidation and Products
These dicarboxylic acids are excreted in the urine The pathways of peroxisomal β and α-oxidation, and microsomal ω-oxidation are not feedback regulated These pathways function to decrease levels of water-insoluble fatty acids or of xenobiotic compounds with a fatty acid–like structure that would become toxic to cells at high concentrations Thus, their rate is regulated by the availability of substrate ω-Oxidation and Products

30
Regulation of Mitochondrial Fatty Acid Oxidation
Regulation by Energy Charge The major site of regulation of the mitochondrial β-oxidation CPT-I, which controls the entry of long-chain fatty acids into the mitochondrion The activity of CPT-I is inhibited by malonyl-CoA, the product of the key regulatory enzyme of fatty acid synthesis: acetyl-CoA carboxylase (ACC) In the fed state, inhibition of CPT-I by malonyl-CoA prevents fatty acid oxidation when glucose is plentiful and when acetyl-CoA is being directed toward fatty acid synthesis When a cell is actively synthesizing fatty acids de novo, the malonyl-CoA concentration in the cytosol increases Subsequent inhibition of CPT-I by malonyl-CoA decreases import of long-chain fatty acids into mitochondria, thereby preventing a futile cycle of simultaneous fatty acid synthesis and β-oxidation
 In the fed state, inhibition of CPT-I by malonyl-CoA prevents fatty acid oxidation when glucose is plentiful and when acetyl-CoA is being directed toward fatty acid synthesis. When a cell is actively synthesizing fatty acids de novo, the malonyl-CoA concentration in the cytosol increases. Subsequent inhibition of CPT-I by malonyl-CoA decreases import of long-chain fatty acids into mitochondria, thereby preventing a futile cycle of simultaneous fatty acid synthesis and β-oxidation.")
31
Conversely, when the energy charge of the cell is low, the increased concentration of AMP activates AMP-activated protein kinase (AMPK), which phosphorylates ACC thereby inhibiting the enzyme –it no longer produces malonyl-CoA Thus, the effect of AMP activation of AMPK is to permit transport of fatty acids into the mitochondrion and ultimately increase the rate of β-oxidation. β-Oxidation of fatty acids within the mitochondrion is also regulated by the energy charge of the cell A high ATP/ADP ratio inhibits entry of reducing equivalents from NADH and FADH2 into the electron-transport chain The resulting increased concentrations of these reduced cofactors in turn prevent the two dehydrogenases of β-oxidation from acting when further generation of ATP is not required In liver, in addition to the regulation by the AMPK, acetyl CoA carboxylase is activated by insulin-dependent mechanisms

32
Regulation by Energy Charge Regulation by Gene Transcription Peroxisome proliferation-activator receptor-α (PPAR- α) is a ligand-activated transcription factor that stimulates fatty acid oxidation (among other things) in liver and muscle Certain metabolites bind ligand-activated PPAR-α which is located in the nucleus This binding induces the synthesis of many different genes, including members of the family of enzymes and proteins involved in β-oxidation
 is a ligand-activated transcription factor that stimulates fatty acid oxidation (among other things) in liver and muscle. Certain metabolites bind ligand-activated PPAR-α which is located in the nucleus. This binding induces the synthesis of many different genes, including members of the family of enzymes and proteins involved in β-oxidation.")
33
Diseases Related to Fatty Acid Oxidation
Medium-Chain Acyl-CoA Dehydrogenase Deficiency (MCADD) The most common genetic defect in fatty acid oxidation is the one that affects the medium-chain acyl-CoA dehydrogenase A deficiency in MCAD activity is associated with high concentrations of both medium chain fatty acid and middle chain acylcarnitines in the plasma and urine of affected persons. Partial oxidation of these intermediate-chain-length fatty acids also generates dicarboxylic fatty acids whose presence in body fluids is diagnostic of MCADD MCADD causes fasting hypoglycemia and muscle weakness Limited utilization of fatty acids as fuels results in an increased dependence on glucose for muscle work At the same time, gluconeogenesis is impaired because of the limited production of both ATP and NADH substrates needed to drive hepatic gluconeogenesis
 The most common genetic defect in fatty acid oxidation is the one that affects the medium-chain acyl-CoA dehydrogenase. A deficiency in MCAD activity is associated with high concentrations of both medium chain fatty acid and middle chain acylcarnitines in the plasma and urine of affected persons. Partial oxidation of these intermediate-chain-length fatty acids also generates dicarboxylic fatty acids whose presence in body fluids is diagnostic of MCADD. MCADD causes fasting hypoglycemia and muscle weakness. Limited utilization of fatty acids as fuels results in an increased dependence on glucose for muscle work. At the same time, gluconeogenesis is impaired because of the limited production of both ATP and NADH substrates needed to drive hepatic gluconeogenesis.")
34
Treatment of persons with MCADD involves avoiding periods of fasting that would tend to produce hypoglycemia Patients with MCADD are advised to take frequent small meals that are relatively high in carbohydrates Genetic defects in many of the other proteins required for mitochondrial β-oxidation have also been documented They include deficiencies in the genes encoding CPT-I, CPT-II, carnitine translocase, acyl-CoA dehydrogenase, and β-hydroxyacyl- CoA dehydrogenase In all of these cases, the clinical manifestations include muscle weakness and fasting hypoglycemia, similar to those observed in patients with MCADD Unlike the situation with MCADD, people with deficiencies in enzymes that metabolize long-chain fatty acids do benefit from diets that contain TAG composed primarily of medium-chain fatty acids. The utilization of these medium-chain fatty acids is not dependent on the carnitine shuttle and LCAD

35
Impaired Peroxisomal Oxidation
X-linked adrenoleukodystrophy (ALD) is a relatively common genetic disease characterized by elevated levels of VLCFA in plasma The accumulation of cholesteryl esters of VLCFA, particularly in the central nervous system, the adrenal glands, and the testes, with adverse effects on membrane structure and steroidogenesis The genetic defect lies in defective transport of VLCFA into the peroxisomes Impaired β-oxidation of VLCFA is also observed in patients with peroxisomal biogenesis disorders such as Zellweger syndrome These persons have a defect in one or more of the proteins that are required to import enzymes into the peroxisome Cells of people with peroxisomal biogenesis disorders are essentially devoid of peroxisomes and exhibit defects in multiple peroxisomal metabolic pathways
 is a relatively common genetic disease characterized by elevated levels of VLCFA in plasma. The accumulation of cholesteryl esters of VLCFA, particularly in the central nervous system, the adrenal glands, and the testes, with adverse effects on membrane structure and steroidogenesis. The genetic defect lies in defective transport of VLCFA into the peroxisomes. Impaired β-oxidation of VLCFA is also observed in patients with peroxisomal biogenesis disorders such as Zellweger syndrome. These persons have a defect in one or more of the proteins that are required to import enzymes into the peroxisome. Cells of people with peroxisomal biogenesis disorders are essentially devoid of peroxisomes and exhibit defects in multiple peroxisomal metabolic pathways.")
36
These include synthesis of ether lipids, α-oxidation of phytanic acid, bile acid synthesis as well as β-oxidation of VLCFA Refsum Disease is caused by a lack of the α-hydroxylase required for α-oxidation of fatty acids, such as phytanic acid Accumulation of large quantities of phytanic acid in the nervous tissue and liver results in chronic polyneuropathy and cerebellar dysfunction The reason peroxisomal defects affect the brain is because VLCFA synthesis occurs in cells of the nervous system, which incorporate them into the lipids of myelin When membranes are remodeled, the VLCFA should be degraded and resynthesized FATTY ACID SYNTHESIS Fatty acid synthesis serves two main functions One is to convert dietary carbohydrates and the carbon skeletons of excess amino acids into triacylglycerols (TAG) that can be stored until needed during periods of fasting
 that can be stored until needed during periods of fasting.")
37
The other function is to produce a variety of fatty acids, which are components of the complex lipids of biological membranes and the precursors of the eicosanoid lipid hormones The major pathway of fatty acid synthesis converts acetyl-CoA molecules derived from dietary carbohydrates and amino acids into the long-chain fatty acid palmitic acid (16:0) Additional enzymes elongate and desaturate both endogenous palmitate and dietary fatty acids to produce a number of other fatty acids, of which the most common are stearic and oleic acid Two fatty acids, linoleic acid and α-linolenic acid are essential fatty acids in the sense that they cannot be synthesized by humans, and as such must be obtained from the diet Although neither linoleic acid nor α-linolenic acid can be synthesized by humans, these dietary fatty acids can be elongated and further desaturated to produce 20- and 22-carbon polyunsaturated fatty acids
 Additional enzymes elongate and desaturate both endogenous palmitate and dietary fatty acids to produce a number of other fatty acids, of which the most common are stearic and oleic acid. Two fatty acids, linoleic acid and α-linolenic acid are essential fatty acids in the sense that they cannot be synthesized by humans, and as such must be obtained from the diet. Although neither linoleic acid nor α-linolenic acid can be synthesized by humans, these dietary fatty acids can be elongated and further desaturated to produce 20- and 22-carbon polyunsaturated fatty acids.")
38
Localization of Fatty Acid Synthesis
Fatty acid synthesis takes place in the cytosol of most cells and tissues; however, hepatocytes and adipocytes are endowed with an especially high capacity for de novo fatty acid synthesis. In the case of fat cells, the fatty acids are esterified to glycerol and stored in the form of TAG In the fasted state, the TAG in adipocytes are hydrolyzed and the free fatty acids are released from adipocytes and transported through the blood bound to albumin Although the liver is the primary site of fatty acid synthesis in humans, hepatocytes do not normally accumulate TAG Instead, the TAG are packaged into very low density lipoproteins (VLDL) and secreted into the circulation In fact, the accumulation of extensive amounts of TAG in the liver is pathologic and can ultimately result in cirrhosis
 and secreted into the circulation. In fact, the accumulation of extensive amounts of TAG in the liver is pathologic and can ultimately result in cirrhosis.")
39
Fatty acid synthesis is most active following a meal
In the first few hours after foods containing carbohydrates such as starch and sucrose have been digested and absorbed, the body experiences a period of transient hyperglycemia This hyperglycemia triggers insulin secretion The resulting high insulin/glucagon ratio signals hepatocytes and adipocytes to take up glucose from the circulation and convert it into fatty acids, and ultimately into TAG Fatty acid synthesis is thus greater when a person is consuming a high-carbohydrate diet than a diet that is relatively low in carbohydrates The Reactions of Fatty Acid Synthesis Even though the main source of of acetyl-CoA for fatty acid synthesis is glucose, acetyl-CoA is also generated from oxidation of the carbon skeletons of excess dietary amino acids and from ethanol

40
In all cases, acetyl-CoA that is not needed for the immediate generation of ATP is routed to the synthesis of fatty acids Once mitochondrial acetyl-CoA has been transported to the cytosol , it serves as the immediate donor of the two carbons at the methyl end of a newly synthesized fatty acid Malonyl-CoA serves as the high-energy, highly reactive donor of the additional acetyl units used during the process of fatty acid synthesis Acetyl-CoA carboxylase (ACC), the cytosolic, biotin-containing enzyme that catalyzes the synthesis of malonyl-CoA, is the rate-limiting step of fatty acid synthesis Like in the case of pyruvate carboxylase , biotin is covalently attached to a lysine residue of ACC
, the cytosolic, biotin-containing enzyme that catalyzes the synthesis of malonyl-CoA, is the rate-limiting step of fatty acid synthesis. Like in the case of pyruvate carboxylase , biotin is covalently attached to a lysine residue of ACC.")
41
During the process of fatty acid synthesis, release of the ionized carboxyl group of malonyl-CoA as CO2 drives the formation of carbon-carbon bonds Fatty Acid Synthase Complex (FAS) Fatty acid synthase sequentially adds 2-carbon units from malonyl-CoA to the growing fatty acyl chain to form palmitate After the addition of each 2-carbon unit, the growing chain undergoes two reduction reactions that require NADPH FAS is a large enzyme composed of two identical dimers, which each have seven catalytic activities and an acyl carrier protein (ACP) segment in a continuous polypeptide chain The ACP segment contains a phosphopantetheine residue that is derived from the cleavage of coenzyme A The two dimers associate in a head-to-tail arrangement, so that the phosphopantetheinyl sulfhydryl group on one subunit and a cysteinyl SH group on another subunit are closely aligned
 Fatty acid synthase sequentially adds 2-carbon units from malonyl-CoA to the growing fatty acyl chain to form palmitate. After the addition of each 2-carbon unit, the growing chain undergoes two reduction reactions that require NADPH. FAS is a large enzyme composed of two identical dimers, which each have seven catalytic activities and an acyl carrier protein (ACP) segment in a continuous polypeptide chain. The ACP segment contains a phosphopantetheine residue that is derived from the cleavage of coenzyme A. The two dimers associate in a head-to-tail arrangement, so that the phosphopantetheinyl sulfhydryl group on one subunit and a cysteinyl SH group on another subunit are closely aligned.")
42
A Comparison of ACP and CoA In the initial step of fatty acid synthesis, an acetyl moiety is transferred from acetyl CoA to the ACP phosphopantetheinyl sulfhydryl group of one subunit and then to the cysteinyl sulfhydryl group of the other subunit The malonyl moiety from malonyl CoA then attaches to the ACP phosphopantetheinyl sulfhydryl group of the first subunit

43
The acetyl and malonyl moieties condense, with the release of the malonyl carboxyl group as CO2
Decarboxylation allows the reaction to go to completion, pulling the whole sequence of reactions in the forward direction A 4-carbon -keto acyl chain is now attached to the ACP phosphopantetheinyl sulfhydryl group A series of three reactions reduces the 4-carbon keto group to an alcohol, removes water to form a double bond, and reduces the double bond NADPH provides the reducing equivalents for these reactions The net result is that the original acetyl group is elongated by two carbons The 4-carbon fatty acyl chain is then transferred to the cysteinyl sulfhydryl group and subsequently condenses with a malonyl group This sequence of reactions is repeated until the chain is 16 carbons in length (palmitate)
")
44
Palmitate is liberated from the enzyme complex by the activity of a seventh enzyme in the complex, thioesterase The free palmitate must be activated to acyl-CoA before it can proceed via any other metabolic pathway Its usual fate is esterification into acylglycerols, chain elongation or desaturation, or esterification to cholesteryl ester The oxidative reactions of the pentose phosphate pathway are the chief source of the hydrogen required for the reductive synthesis of fatty acids Tissues specializing in active lipogenesis possess an active pentose phosphate pathway. Moreover, both metabolic pathways are found in the cytosol of the cell; so, there are no membranes or permeability barriers against the transfer of NADPH The other main source of NADPH is the reaction that converts malate to pyruvate catalyzed by the malic enzyme

45
The FAS Complex

46
Loading and Condensation
Reduction

47
Modification Reactions
Most cells have the ability to increase the chain length and degree of unsaturation of long-chain fatty acids Modification of both dietary-derived fatty acids and the palmitate synthesized de novo in the body accounts for the great diversity of structural fatty acids in membrane lipids and those involved in signaling Fatty Acid Chain Elongation Elongation of fatty acids occurs primarily in the smooth endoplasmic reticulum and utilizes malonyl-CoA to add two-carbon units to long-chain fatty acyl-CoAs. There is a minor, secondary chain elongation system (elongase) in mitochondria that utilizes acetyl-CoA as the two-carbon donor and it appears to be involved primarily in the synthesis of lipoic acid, a cofactor for PDC and α-ketoglutarate dehydrogenase. The elongation system is comprised of a condensing enzyme that adds two carbons to a molecule of fatty acyl-CoA
 in mitochondria that utilizes acetyl-CoA as the two-carbon donor and it appears to be involved primarily in the synthesis of lipoic acid, a cofactor for PDC and α-ketoglutarate dehydrogenase. The elongation system is comprised of a condensing enzyme that adds two carbons to a molecule of fatty acyl-CoA.")
48
It also has three additional enzyme activities:β-ketoacyl-CoA reductase, β-hydroxyacyl-CoA dehydratase, and enoyl-CoA reductase, whose activities are similar to the enzymes of FAS that catalyze the reduction sequence The major elongation reaction that occurs in the body involves the conversion of palmitoyl CoA (C16) to stearyl CoA (C18) Very-long-chain fatty acids (C22 to C24) are also produced, particularly in the brain Desaturation of Fatty Acids Desaturation of fatty acids involves a process that requires O2, NADH, and cytochrome b5 The reaction, which occurs in the endoplasmic reticulum, results in the oxidation of both the fatty acid and NADH The desaturation complex includes the actual desaturase enzyme, cytochrome b5 which serves as an electron acceptor, and NADH-cytochrome b5 reductase, which contains FAD as a prosthetic group
 to stearyl CoA (C18) Very-long-chain fatty acids (C22 to C24) are also produced, particularly in the brain. Desaturation of Fatty Acids. Desaturation of fatty acids involves a process that requires O2, NADH, and cytochrome b5. The reaction, which occurs in the endoplasmic reticulum, results in the oxidation of both the fatty acid and NADH. The desaturation complex includes the actual desaturase enzyme, cytochrome b5 which serves as an electron acceptor, and NADH-cytochrome b5 reductase, which contains FAD as a prosthetic group.")
49
The most common desaturation reactions involve the placement of a double bond between carbons 9 and 10 in the conversion of palmitic acid to palmitoleic acid (16:1,Δ9) and the conversion of stearic acid to oleic acid (18:1, Δ9) Other positions that can be desaturated in humans include carbons 4, 5, and 6 Desaturation

50
Polyunsaturated fatty acids with double bonds three carbons from the methyl end (ω-3 fatty acids) and six carbons from the methyl end (ω-6 fatty acids) are required for the synthesis of eicosanoids Since human cells cannot introduce double bonds beyond carbon 9 from the carboxyl end of long-chain fatty acids, ω-3 and ω-6 fatty acids must be present in the diet or the diet must contain other fatty acids that can be converted to these fatty acids We obtain ω-3 and ω-6 polyunsaturated fatty acids mainly from dietary plant oils that contain the ω-6 fatty acid linoleic acid (18:2, Δ9,12) and the ω-3 fatty acid α-linolenic acid (18:3, Δ9,12,15) In the body, linoleic acid can be converted by elongation and desaturation reactions to arachidonic acid (20:4, Δ5,8,11,14), which is used for the synthesis of the major class of human eicosanoids
 and the ω-3 fatty acid α-linolenic acid (18:3, Δ9,12,15) In the body, linoleic acid can be converted by elongation and desaturation reactions to arachidonic acid (20:4, Δ5,8,11,14), which is used for the synthesis of the major class of human eicosanoids.")
51
Elongation and desaturation of α-linolenic acid produces eicosapentaenoic acid (EPA; 20:5, Δ5,8,11,14,17), which is the precursor of a different class of eicosanoids Another important product of the elongation and desaturation of fatty acids in the body is docosahexaenoic acid (DHA,22:6 Δ4,7,10,13,16,19 ) Regulation of Fatty Acid Synthesis Regulation of the Activity of Acetyl-CoA Carboxylase ACC is regulated both by allosteric modulators and by phosphorylation/dephosphorylation The main allosteric activator is citrate By contrast, palmitoyl-CoA and other long-chain fatty acyl-CoAs inhibit ACC; this phenomenon is an example of feedback inhibition Citrate activates ACC by inducing formation of an active filamentous polymer from relatively inactive enzyme dimers Activation of ACC by citrate reflects the energy status of the cell
 Regulation of Fatty Acid Synthesis. Regulation of the Activity of Acetyl-CoA Carboxylase. ACC is regulated both by allosteric modulators and by phosphorylation/dephosphorylation. The main allosteric activator is citrate. By contrast, palmitoyl-CoA and other long-chain fatty acyl-CoAs inhibit ACC; this phenomenon is an example of feedback inhibition. Citrate activates ACC by inducing formation of an active filamentous polymer from relatively inactive enzyme dimers. Activation of ACC by citrate reflects the energy status of the cell.")
52
Polyunsaturated Fatty Acids
Elongation and Desaturation of Polyunsaturated Fatty Acids

53
In the fed state, when tissues are energy-replete, the high mitochondrial concentration of ATP inhibits isocitrate dehydrogenase and citrate is transported to the cytosol where it can serve as a signal for the cell to synthesize fatty acids from excess acetyl-CoA In the fasted state when the insulin/glucagon ratio is low, the concentration of cAMP increases, and cAMP-activated protein kinase A (PKA) phosphorylates and inhibits ACC ACC is also inhibited by the action of a second protein kinase, AMP-activated protein kinase (AMPK), whose activity reflects depletion of intracellular ATP In the fed state, insulin activates the protein phosphatase, which dephosphorylates ACC, thus increasing its activity Regulation of Enzyme Synthesis Insulin is a powerful anabolic signal, particularly in hepatocytes and adipocytes, where it induces synthesis of the lipogenic family of enzymes
 phosphorylates and inhibits ACC. ACC is also inhibited by the action of a second protein kinase, AMP-activated protein kinase (AMPK), whose activity reflects depletion of intracellular ATP. In the fed state, insulin activates the protein phosphatase, which dephosphorylates ACC, thus increasing its activity. Regulation of Enzyme Synthesis. Insulin is a powerful anabolic signal, particularly in hepatocytes and adipocytes, where it induces synthesis of the lipogenic family of enzymes.")
54
These enzymes include ACC, citrate lyase, the malic enzyme, G6PD, pyruvate kinase, and the FAS complex The mechanism underlying this action by insulin involves activation of the sterol regulatory element-binding protein- I (SREBP- I), a membrane-bound transcription factor that enhances transcription of the genes encoding proteins required for fatty acid synthesis Glucagon, on the other hand, represses de novo synthesis of these enzymes in adipocytes and liver, and stimulates degradation of the lipogenic family of enzyme proteins The Regulation of ACC
, a membrane-bound transcription factor that enhances transcription of the genes encoding proteins required for fatty acid synthesis. Glucagon, on the other hand, represses de novo synthesis of these enzymes in adipocytes and liver, and stimulates degradation of the lipogenic family of enzyme proteins. The Regulation of ACC.")
55
THE EICOSANOIDS The eicosanoids are a complex family of bioactive lipid messengers generated by oxygenation of 20-carbon polyunsaturated fatty acids, primarily arachidonic acid Eicosanoids are local-acting autocrine and paracrine hormones that stimulate cells adjacent to their site of synthesis In general, eicosanoids have a short half-life, usually on the order of minutes They are not stored in cells but instead are released as soon as they are synthesized Eicosanoids fall into two main classes: (1) prostanoids that have a ring structure, including prostaglandins, thromboxanes, and prostacyclins, and (2) linear eicosanoids consisting of leukotrienes, lipoxins, and hydroxyeicosatetraenoic acids (HETEs)
 prostanoids that have a ring structure, including prostaglandins, thromboxanes, and prostacyclins, and. (2) linear eicosanoids consisting of leukotrienes, lipoxins, and hydroxyeicosatetraenoic acids (HETEs)")
56
The Prostanoids The term prostaglandin reflects the original isolation of these molecules from seminal fluid, into which they are secreted by the seminal glands (rather than the prostate). Prostaglandins act to modulate many physiological functions, including blood pressure, uterine contraction, and the induction of pain and fever Prostaglandins are designated PGA, PGD, PGE, or PGF, based on the functional groups on the cyclopentane ring that is comprised of carbons 8 through 12 For example, PGE2 contains a 9-keto and an 1 1-hydroxy group, while PGF2α, contains two hydroxy groups; α designates the stereochemistry of the C9 hydroxyl group All of the prostanoids that are derived from arachidonic acid have numeral 2 as a subscript, referring to the number of carbon-carbon double bonds in the two side chains
. Prostaglandins act to modulate many physiological functions, including blood pressure, uterine contraction, and the induction of pain and fever. Prostaglandins are designated PGA, PGD, PGE, or PGF, based on the functional groups on the cyclopentane ring that is comprised of carbons 8 through 12. For example, PGE2 contains a 9-keto and an 1 1-hydroxy group, while PGF2α, contains two hydroxy groups; α designates the stereochemistry of the C9 hydroxyl group. All of the prostanoids that are derived from arachidonic acid have numeral 2 as a subscript, referring to the number of carbon-carbon double bonds in the two side chains.")
57
Prostacyclin (PGI2) and thromboxane (TX2) are two prostanoids that have somewhat unusual ring structures The term thromboxane refers to the platelet-aggregating activity, which has thrombus-forming potential Linear Eicosanoids Unlike the prostanoids, which contain a ring element in their structure, the leukotrienes are linear molecules The term leukotriene derives from their cell of origin (leukocytes) and the fact that their structures contain three carbon-carbon double bonds in conjugation The most important leukotrienes in humans are LTA4 and LTB4 and their cysteinyl derivatives, LTC4, LTD4, and LTE4 All are derived from arachidonic acid and contain four double bonds The cysteinyl leukotrienes constitute the slow-reacting substance of anaphylaxis (SRS-A)
 and the fact that their structures contain three carbon-carbon double bonds in conjugation. The most important leukotrienes in humans are LTA4 and LTB4 and their cysteinyl derivatives, LTC4, LTD4, and LTE4. All are derived from arachidonic acid and contain four double bonds. The cysteinyl leukotrienes constitute the slow-reacting substance of anaphylaxis (SRS-A)")
58
SRS-A promotes smooth muscle contraction, constriction of pulmonary airways, trachea and intestine, and increases in capillary permeability (edema) HETEs are closely related in structure to cysteinyl leukotrienes; they are involved in chemotaxis, degranulation of polymorphonuclear cells, etc Lipoxins have got structures that are distinct from leukotrienes or HETEs; have got anti angiogenic properties, … The Synthesis of Eicosanoids Synthesis of eicosanoids is initiated by release of arachidonic acid The arachidonic acid present in membrane phospholipids is released from the lipid bilayer as a consequence of the activation of membrane-bound phospholipase A2 or C This activation occurs when a variety of stimuli (agonists), such as histamine and the cytokines, interact with a specific plasma membrane receptor on the target cell surface
, such as histamine and the cytokines, interact with a specific plasma membrane receptor on the target cell surface.")
59
The Release of Arachidonate
Phospholipase A2 is specific for the sn-2 position of phosphoacylglycerols, the site of attachment of arachidonic acid to the glycerol moiety Phospholipase C hydrolyzes phosphorylated inositol from the inositol glycerophospholipids, generating a diacylglycerol containing arachidonic acid This arachidonic acid is subsequently released by the action of other lipases The Release of Arachidonate

60
The Synthesis of Prostanoids
The key enzyme of prostanoid biosynthesis is the bifunctional enzyme prostaglandin G/H synthase (PGS) PGS is a single-polypeptide-chain enzyme that has two catalytic sites One catalytic site has cyclooxygenase activity which catalyzes the addition of two molecules of oxygen to arachidonate to form the initial prostaglandin, which is PGG2 The second catalytic site is a glutathione-dependent, heme-containing peroxidase that converts the hydroperoxide group (-OOH) on carbon 15 of PGG2 to a hydroxyl group; gives PGH2 There are two major isozymes of prostaglandin G/H synthase or cyclooxygenase (COX), commonly called COX-1 and COX-2 COX-1 is primarily a constitutive enzyme of gastric mucosa, platelets, vascular endothelium, and kidney COX-2 is inducible and expressed in activated macrophages and monocytes as well as smooth muscle and epithelial cells
 PGS is a single-polypeptide-chain enzyme that has two catalytic sites. One catalytic site has cyclooxygenase activity which catalyzes the addition of two molecules of oxygen to arachidonate to form the initial prostaglandin, which is PGG2. The second catalytic site is a glutathione-dependent, heme-containing peroxidase that converts the hydroperoxide group (-OOH) on carbon 15 of PGG2 to a hydroxyl group; gives PGH2. There are two major isozymes of prostaglandin G/H synthase or cyclooxygenase (COX), commonly called COX-1 and COX-2. COX-1 is primarily a constitutive enzyme of gastric mucosa, platelets, vascular endothelium, and kidney. COX-2 is inducible and expressed in activated macrophages and monocytes as well as smooth muscle and epithelial cells.")
61
Studies have identified a third PGS isozyme, COX-3, which is made from the COX-1 gene but retains intron 1 in its mRNA COX-3 is expressed in the cerebral cortex and is inhibited by analgesic and antipyretic drugs, such as acetaminophen (Tylenol), that do not inhibit COX-1 and COX-2 A family of prostaglandin synthases (i.e., PGD synthase, PGE synthase, PGI synthase) convert PGH2 to the various prostaglandins PGH2 is also the precursor of thromboxane A2 The one prostaglandin that is not synthesized directly from PGH2 is PGF2α, which is synthesized from PGE2 The Synthesis of Linear Eicosanoids Lipoxygenases are dioxygenases that attach both atoms of molecular oxygen to a particular carbon atom of arachidonic acid (e.g., position 5 , 12, or 15)
, that do not inhibit COX-1 and COX-2. A family of prostaglandin synthases (i.e., PGD synthase, PGE synthase, PGI synthase) convert PGH2 to the various prostaglandins. PGH2 is also the precursor of thromboxane A2. The one prostaglandin that is not synthesized directly from PGH2 is PGF2α, which is synthesized from PGE2. The Synthesis of Linear Eicosanoids. Lipoxygenases are dioxygenases that attach both atoms of molecular oxygen to a particular carbon atom of arachidonic acid (e.g., position 5 , 12, or 15)")
62
The Synthesis of Prostanoids
from Arachidonate

63
In humans, the most important leukotrienes are the 5- lipoxygenase (5-LOX) products, which mediate inflammation Their synthesis is initiated by the formation of hydroperoxyeicosatetraenoic acid (5-HPETE) The reduction of HPETEs to HETEs occurs spontaneously or is catalyzed by peroxidases 5-LOX contains two enzymatic activities: the dioxygenase activity that converts arachidonic acid to 5-HPETE and a dehydrase activity that transforms 5-HPETE to LTA4 LTA4 is an important branch point in the pathway of leukotriene synthesis. It can be converted to LTB4 or to LTC4 Successive degradation of LTC4 gives LTD4 and LTE4, respectively Eicosanoids from Other Precursors The major precursor for the synthesis of human eicosanoids is arachidonic acid, which gives rise to the 2-series of prostaglandins and the 4-series of leukotrienes
 The reduction of HPETEs to HETEs occurs spontaneously or is catalyzed by peroxidases. 5-LOX contains two enzymatic activities: the dioxygenase activity that converts arachidonic acid to 5-HPETE and a dehydrase activity that transforms 5-HPETE to LTA4. LTA4 is an important branch point in the pathway of leukotriene synthesis. It can be converted to LTB4 or to LTC4. Successive degradation of LTC4 gives LTD4 and LTE4, respectively. Eicosanoids from Other Precursors. The major precursor for the synthesis of human eicosanoids is arachidonic acid, which gives rise to the 2-series of prostaglandins and the 4-series of leukotrienes.")
64
The Synthesis of Leukotrienes
from Arachidonate

65
Although eicosanoids can also be synthesized from dihomo-y -linoleic acid (DGLA, 20:3; ω-6;), the immediate precursor of arachidonic acid, there is little synthesis of 1-series prostaglandins and 3-series leukotrienes in humans PGE1 has, however, been utilized extensively as a pharmacological agent; one application of its vasodilatory effects has been in the treatment of erectile dysfunction Eicosapentaenoic acid (EPA, 20:5; ω-3) is the other physiologically significant precursor of human eicosanoids in humans The main dietary source of EPA is fish oil, particularly oil from cold-water marine fish; some EPA is also synthesized from dietary α-linolenic acid EPA is the precursor to a family of eicosanoids, each of which has one more double bond than the corresponding eicosanoid derived from arachidonate
 is the other physiologically significant precursor of human eicosanoids in humans. The main dietary source of EPA is fish oil, particularly oil from cold-water marine fish; some EPA is also synthesized from dietary α-linolenic acid. EPA is the precursor to a family of eicosanoids, each of which has one more double bond than the corresponding eicosanoid derived from arachidonate.")
66
An increased dietary intake of fish oil can raise the ratio of membrane phospholipid EPA to arachidonate Although dietary fish oils have been shown to be cardioprotective, anti-inflammatory, and anticarcinogenic, it is still unclear how much these benefits are due to the partial replacement of arachidonate-derived eicosanoids with those synthesized from EPA Series of Eicosanoids

67
Sites of Synthesis of Eicosanoids
With the exception of red blood cells, eicosanoids are produced and released by nearly all human cells Gastrointestinal Tract Prostaglandins serve a cytoprotective role in the stomach PGE2 is synthesized by epithelial and smooth muscle cells in the stomach, where it reduces gastric acid secretion while stimulating the production of protective mucus For this reason, synthetic prostaglandins are helpful in promoting the healing of gastric ulcers Cardiovascular System In blood vessels, different prostaglandins have opposing effects For example, platelets produce TXA2, which promotes platelet aggregation, whereas vascular endothelial cells produce prostacyclin (PGI2), which inhibits platelet aggregation
, which inhibits platelet aggregation.")
68
Both PGE2 and PGI2 are vasodilators that lower systemic arterial pressure, thereby increasing local blood flow and decreasing peripheral resistance By contrast, both TXA2 and PGF2α, (produced by vascular smooth muscle) are vasoconstrictors The Kidneys PGE2 is the major prostaglandin in the kidney, and the collecting ducts are the main site of its production PGE2 dilates renal blood vessels and increases blood flow through the kidney It is also an important stimulator of renin release, thus contributing to the regulation of sodium excretion The Lungs Monocytes and neutrophils in the lungs produce LTB4, HETE, and SRS-A which are bronchoconstrictors LTC4 is more potent than histamine in contracting the smooth muscles of bronchi
 are vasoconstrictors. The Kidneys. PGE2 is the major prostaglandin in the kidney, and the collecting ducts are the main site of its production. PGE2 dilates renal blood vessels and increases blood flow through the kidney. It is also an important stimulator of renin release, thus contributing to the regulation of sodium excretion. The Lungs. Monocytes and neutrophils in the lungs produce LTB4, 5-HETE, and SRS-A which are bronchoconstrictors. LTC4 is more potent than histamine in contracting the smooth muscles of bronchi.")
69
Female Reproductive Tract
PGE2 within the ovarian follicle is essential for ovulation During parturition, prostaglandins soften tissues in the cervix and stimulate uterine contractions to expel the fetus Eicosanoids and Inflammation Prostaglandins, PGE2 in particular, are mediators of the edema, erythema (redness of the skin), and the fever and pain associated with inflammation Inflammatory reactions are most often treated with corticosteroids that inhibit prostaglandin synthesis PGE2, generated in immune cells (e.g., macrophages, mast cells, B cells), evokes chemotaxis of T cells It is thought that pyrogens (fever-inducing agents) activate the prostaglandin synthesis pathway with release of PGE2 in the hypothalamus, where body temperature is regulated Synthesis of leukotrienes and HETEs is also up-regulated under conditions of allergy and inflammation
, and the fever and pain associated with inflammation. Inflammatory reactions are most often treated with corticosteroids that inhibit prostaglandin synthesis. PGE2, generated in immune cells (e.g., macrophages, mast cells, B cells), evokes chemotaxis of T cells. It is thought that pyrogens (fever-inducing agents) activate the prostaglandin synthesis pathway with release of PGE2 in the hypothalamus, where body temperature is regulated. Synthesis of leukotrienes and HETEs is also up-regulated under conditions of allergy and inflammation.")
70
Regulation of Eicosanoid Synthesis and Activity
Regulation of Arachidonate Mobilization Activation of phospholipase A2 is crucial for the release of arachidonic acid, which serves as a substrate for eicosanoid synthesis. Phospholipase A2 is activated in a cell-specific manner by a variety of agonists (e.g., by thrombin in platelets) Steroidal anti-inflammatory drugs such as prednisone and betamethasone block prostaglandin release in part by inducing the synthesis of inhibitors of PLA2 Regulation of Prostaglandin G/H Synthase Prostaglandin G/H synthase represents the committed step in prostaglandin and thromboxane synthesis Whereas COX-1 is constitutive in many cell types, synthesis of COX-2 in various cells is induced by a variety of cytokines and lipid mediators
 Steroidal anti-inflammatory drugs such as prednisone and betamethasone block prostaglandin release in part by inducing the synthesis of inhibitors of PLA2. Regulation of Prostaglandin G/H Synthase. Prostaglandin G/H synthase represents the committed step in prostaglandin and thromboxane synthesis. Whereas COX-1 is constitutive in many cell types, synthesis of COX-2 in various cells is induced by a variety of cytokines and lipid mediators.")
71
Although glucocorticoids had long been assumed to block prostaglandin synthesis at the level of phospholipase A2, there is increasing evidence that they also suppress the synthesis of COX-2 in many cell types Prostaglandin synthase, a major target for pharmacological intervention, is inhibited by non-steroidal anti-inflammatory drugs (NSAIDs) such as aspirin (acetylsalicylic acid) and ibuprofen Most NSAIDs act as reversible inhibitors of the cyclo-oxygenase component of PGS Aspirin acts differently in that it irreversibly inhibits PGS by acetylating the hydroxyl group of a particular serine hydroxyl at the active site of cyclooxygenase Low-dose aspirin regimens are often used to decrease the risk of thrombosis and coronary heart disease in older persons
 such as aspirin (acetylsalicylic acid) and ibuprofen. Most NSAIDs act as reversible inhibitors of the cyclo-oxygenase component of PGS. Aspirin acts differently in that it irreversibly inhibits PGS by acetylating the hydroxyl group of a particular serine hydroxyl at the active site of cyclooxygenase. Low-dose aspirin regimens are often used to decrease the risk of thrombosis and coronary heart disease in older persons.")
72
This therapy is effective because circulating platelets are unable to synthesize more prostaglandin synthase to replace that which has been inactivated Recent pharmaceutical efforts have focused on the development of selective COX-2 inhibitors such as Celebrex (celecoxib), with the goal of developing anti-inflammatory drugs less likely to cause the gastric toxicity associated with chronic use of NSAIDs that block COX- 1 as well Both Celebrex and Vioxx (now withdrawn from the market) have been associated with increased adverse cardiovascular events Possible reasons for the adverse effects include: Decreased production of antithrombotic PGI2 while not inhibiting COX- 1 -mediated synthesis of thromboxane A2 in platelets COX 2 can be constitutive in the kidneys where PGE2 promotes dilation of arterioles and the excretion of Na+ and water
, with the goal of developing anti-inflammatory drugs less likely to cause the gastric toxicity associated with chronic use of NSAIDs that block COX- 1 as well. Both Celebrex and Vioxx (now withdrawn from the market) have been associated with increased adverse cardiovascular events. Possible reasons for the adverse effects include: Decreased production of antithrombotic PGI2 while not inhibiting COX- 1 -mediated synthesis of thromboxane A2 in platelets. COX 2 can be constitutive in the kidneys where PGE2 promotes dilation of arterioles and the excretion of Na+ and water.")
73
The Action of NSAIDs

74
Regulation of Leukotriene Metabolism
Current therapies for asthma include use of 5-lipoxygenase inhibitors such as zileuton, and cysteinyl-leukotriene (cysLT) receptor antagonists such as montelukast Therapeutic Uses of Eicosanoids Exogenous prostaglandins have a number of therapeutic uses. For example, in the fetus, PGE2 keeps ductus arteriosus open prior to birth In infants born with congenital abnormalities that can be corrected surgically, infusion of PGE2 will maintain blood flow through the ductus until surgery is performed Conversely, if the ductus remains open after birth in an otherwise normal infant, closure can be hastened by the COX inhibitor indomethacin PGE2 has also been used to induce cervical ripening and uterine contractions, leading to parturition
 receptor antagonists such as montelukast. Therapeutic Uses of Eicosanoids. Exogenous prostaglandins have a number of therapeutic uses. For example, in the fetus, PGE2 keeps ductus arteriosus open prior to birth. In infants born with congenital abnormalities that can be corrected surgically, infusion of PGE2 will maintain blood flow through the ductus until surgery is performed. Conversely, if the ductus remains open after birth in an otherwise normal infant, closure can be hastened by the COX inhibitor indomethacin. PGE2 has also been used to induce cervical ripening and uterine contractions, leading to parturition.")
75
Misoprostol is a synthetic PGE1 analog used to prevent NSAID-induced ulcers
In many countries, misoprostol is also used in combination with the synthetic steroid RU486 to block the action of progesterone and induce medical (as opposed to surgical) abortions Outroduction Trans Fatty Acids and Omega-3 vs 6 Fatty Acids Trans fatty acids are produced during the commercial hydrogenation of plant oils Some margarines contain these fatty acids, as do some commercially prepared snack foods (e.g. biscuits, cookies, cakes, chips) In addition, bacteria in the rumen of ruminants produce trans fatty acids, which are therefore present in dairy produce and meat
 abortions. Outroduction. Trans Fatty Acids and Omega-3 vs 6 Fatty Acids. Trans fatty acids are produced during the commercial hydrogenation of plant oils. Some margarines contain these fatty acids, as do some commercially prepared snack foods (e.g. biscuits, cookies, cakes, chips) In addition, bacteria in the rumen of ruminants produce trans fatty acids, which are therefore present in dairy produce and meat.")
76
Trans fatty acids can be incorporated into the phospholipids of the plasma membrane of endothelial and other cells, resulting in damage to the membranes Furthermore, these abnormal fatty acids can interfere in the production of thromboxanes, prostacyclins or leukotrienes Series-2 thromboxanes (from the ω-6 arachidonate) have strong positive effects on aggregation of platelets whereas those of the 3-series (from the ω-6 EPA) have much less effect Hence, a low chronic intake of omega-3 fatty acid (linolenic) and a high intake of omega-6 fatty acid (linoleic) increases the formation of thromboxanes of the series 2 and increases the formation of prostaglandin of series 2 which has a lower vasodilatory effect, which increases the risk of thrombosis ω -3 fatty acids are quantitatively important components of the membranes of neurones in the brain; important in development of the brain and the repair of damaged membranes Chronic deficiency could increase the risk of development of some disorders, including depression,schizophrenia and ADD
 have strong positive effects on aggregation of platelets whereas those of the 3-series (from the ω-6 EPA) have much less effect. Hence, a low chronic intake of omega-3 fatty acid (linolenic) and a high intake of omega-6 fatty acid (linoleic) increases the formation of thromboxanes of the series 2 and increases the formation of prostaglandin of series 2 which has a lower vasodilatory effect, which increases the risk of thrombosis. ω -3 fatty acids are quantitatively important components of the membranes of neurones in the brain; important in development of the brain and the repair of damaged membranes. Chronic deficiency could increase the risk of development of some disorders, including depression,schizophrenia and ADD.")
78
TRIACYLGLYCEROL TRANSPORT AND METABOLISM
The main function of TAG in the body is to provide a compact and relatively unlimited means for storing energy At 9 kcal/g, the energy content of TAG is more than twice that of the other major form of energy that humans store, namely glycogen (4.1 kcal/g) The fatty acids that comprise TAG are highly reduced. Except for the carboxyl group, most of the carbon atoms of a fatty acid have two hydrogen atoms attached to them and are bonded to another carbon atom It is the energy that is released during the oxidation of these C-H and C-C bonds that ultimately supports the synthesis of ATP by oxidative phosphorylation Furthermore, whereas glycogen binds more than twice its weight in water, TAG are hydrophobic, such that only a small part of the mass of adipose tissue is water
 The fatty acids that comprise TAG are highly reduced. Except for the carboxyl group, most of the carbon atoms of a fatty acid have two hydrogen atoms attached to them and are bonded to another carbon atom. It is the energy that is released during the oxidation of these C-H and C-C bonds that ultimately supports the synthesis of ATP by oxidative phosphorylation. Furthermore, whereas glycogen binds more than twice its weight in water, TAG are hydrophobic, such that only a small part of the mass of adipose tissue is water.")
79
This contrast in water-binding capacity between fat and glycogen means that on a weight basis, it is more economical to store energy in the form of fat than in the form of glycogen In the fed state, the TAG in circulating lipoproteins (both diet-derived (chylomicrons) and endogenously synthesized (VLDL)) are hydrolyzed in the capillaries of skeletal muscle, adipose, and other tissues by lipoprotein lipase, and the resulting free fatty acids are taken up by adipocytes and muscle cells Between meals or during a fast when glycogen stores have been depleted, TAG in adipocytes are hydrolyzed to free fatty acids and glycerol These fatty acids circulate bound to albumin rather than as components of triacylglycerol-containing lipoproteins Formation of non-covalent complexes between fatty acids and albumin solubilizes the long-chain fatty acids Transport on albumin also minimizes the potential damage to membranes by the detergent activity of free fatty acids
 and endogenously synthesized (VLDL)) are hydrolyzed in the capillaries of skeletal muscle, adipose, and other tissues by lipoprotein lipase, and the resulting free fatty acids are taken up by adipocytes and muscle cells. Between meals or during a fast when glycogen stores have been depleted, TAG in adipocytes are hydrolyzed to free fatty acids and glycerol. These fatty acids circulate bound to albumin rather than as components of triacylglycerol-containing lipoproteins. Formation of non-covalent complexes between fatty acids and albumin solubilizes the long-chain fatty acids. Transport on albumin also minimizes the potential damage to membranes by the detergent activity of free fatty acids.")
80
The fatty acids in the sn-1 and -3 positions of TAG tend to be long-chain saturated fatty acids (e.g., palmitic acid, stearic acid) or monounsaturated fatty acids (e.g., oleic acid), whereas those in the sn-2 position tend to be polyunsaturated fatty acids (e.g., linoleic acid, α-linolenic acid, arachidonic acid) The three most abundant fatty acids in the TAG of adipose tissue and plasma lipoproteins are palmitic acid, oleic acid, and linoleic acid Synthesis of Triacylglycerols The pathway for triacylglycerol synthesis in most tissues, including liver and adipocytes, utilizes glycerol 3-phosphate and fatty acyl-CoA

81
The activated fatty acids (i. e
The activated fatty acids (i.e., fatty acids attached to CoA) are derived either from endogenous de novo fatty acid synthesis or from dietary fats By contrast, triacylglycerol synthesis in the small intestine begins with 2-monoacylglycerol The sources of glycerol 3-phosphate, which provides the glycerol moiety for triacylglycerol synthesis, differ in liver and adipose tissue In liver, glycerol 3-phosphate is produced from the phosphorylation of glycerol by glycerol kinase or from the glycerol 3-phosphate dehydrogenase reduction of DHAP derived from glycolysis Adipose tissue lacks glycerol kinase and can produce glycerol 3-phosphate only from glucose via dihydroxyacetone phosphate In the fasting state, adipose and the liver also undergo glyceroneogenesis –synthesize glycerol 3-phosphate from gluconeogenic precursors, such as alanine, aspartate, malate
 are derived either from endogenous de novo fatty acid synthesis or from dietary fats. By contrast, triacylglycerol synthesis in the small intestine begins with 2-monoacylglycerol. The sources of glycerol 3-phosphate, which provides the glycerol moiety for triacylglycerol synthesis, differ in liver and adipose tissue. In liver, glycerol 3-phosphate is produced from the phosphorylation of glycerol by glycerol kinase or from the glycerol 3-phosphate dehydrogenase reduction of DHAP derived from glycolysis. Adipose tissue lacks glycerol kinase and can produce glycerol 3-phosphate only from glucose via dihydroxyacetone phosphate. In the fasting state, adipose and the liver also undergo glyceroneogenesis –synthesize glycerol 3-phosphate from gluconeogenic precursors, such as alanine, aspartate, malate.")
82
In both adipose tissue and liver, TAG are produced by a pathway in which glycerol 3-phosphate reacts with two fatty acyl-CoA molecules to form phosphatidic acid Dephosphorylation of phosphatidic acid produces diacylglycerol Another fatty acyl-CoA reacts with the diacylglycerol to form TAG The Synthesis of Triacylglycerols

83
Transport of Triacylglycerols in the Blood
Plasma contains a class of macromolecular aggregates called lipoproteins that transport otherwise highly water-insoluble lipids-cholesteryl esters and TAG in particular-in the circulation Lipoproteins also play a key role in the metabolism of these lipids and facilitate the two-way exchange of TAG between tissues and the blood In addition, cholesteryl esters, TAG and proteins undergo exchange between the various lipoproteins The surface of a lipoprotein particle is coated with proteins, phospholipids, and free (non-esterified) cholesterol The polar ends of the amphipathic lipids face the surface of the lipoprotein, whereas the hydrophobic portions are oriented toward the center of the particle
 cholesterol. The polar ends of the amphipathic lipids face the surface of the lipoprotein, whereas the hydrophobic portions are oriented toward the center of the particle.")
84
The core of the lipoproteins is composed of highly non-polar lipids such as TAG and cholesteryl esters The proteins (or apoproteins as they are called when separated from the lipids) serve a number of functions, including stabilization of the lipoprotein’s structure and as recognition sites for lipoprotein receptors on cell membranes The lipoproteins are differentiated on the basis of density The most dense lipoprotein class is called high-density lipoprotein (HDL) followed by low-density lipoprotein (LDL), very low-density lipoprotein (VLDL) and chylomicrons The major triacylglycerol-transporting lipoproteins are VLDL and chylomicrons LDL and HDL transport primarily cholesteryl esters HDL also contains some triacylglycerol; it is also the lipoprotein with the highest amount of protein
 serve a number of functions, including stabilization of the lipoprotein’s structure and as recognition sites for lipoprotein receptors on cell membranes. The lipoproteins are differentiated on the basis of density. The most dense lipoprotein class is called high-density lipoprotein (HDL) followed by low-density lipoprotein (LDL), very low-density lipoprotein (VLDL) and chylomicrons. The major triacylglycerol-transporting lipoproteins are VLDL and chylomicrons. LDL and HDL transport primarily cholesteryl esters. HDL also contains some triacylglycerol; it is also the lipoprotein with the highest amount of protein.")
85
The Basic Pattern of Lipoproteins

86
The Fate of Chylomicrons
The triacylglycerols of the chylomicrons are digested by lipoprotein lipase (LPL) attached to the proteoglycans in the basement membranes of endothelial cells that line the capillary walls. LPL is activated by apo C II LPL is produced mainly by adipose, muscle cells (particularly cardiac muscle), and cells of the lactating mammary gland The isozyme synthesized in adipose cells has a higher Km than the isozyme synthesized in muscle cells Therefore, adipose LPL is more active after a meal, when chylomicrons levels are elevated in the blood Insulin stimulates the synthesis and secretion of adipose LPL, such that after a meal, when triglyceride levels increase in circulation, LPL has been upregulated (through insulin release) to facilitate the hydrolysis of fatty acids from the TAG The fatty acids released from triacylglycerols by LPL are not very soluble in water
 attached to the proteoglycans in the basement membranes of endothelial cells that line the capillary walls. LPL is activated by apo C II. LPL is produced mainly by adipose, muscle cells (particularly cardiac muscle), and cells of the lactating mammary gland. The isozyme synthesized in adipose cells has a higher Km than the isozyme synthesized in muscle cells. Therefore, adipose LPL is more active after a meal, when chylomicrons levels are elevated in the blood. Insulin stimulates the synthesis and secretion of adipose LPL, such that after a meal, when triglyceride levels increase in circulation, LPL has been upregulated (through insulin release) to facilitate the hydrolysis of fatty acids from the TAG. The fatty acids released from triacylglycerols by LPL are not very soluble in water.")
87
They become soluble in blood by forming complexes with albumin
The major fate of the fatty acids is storage as triacylglycerol in adipose tissue. However, these fatty acids also may be oxidized for energy in muscle and other tissues The LPL in the capillaries of muscle cells has a lower Km than adipose LPL Thus, muscle cells can obtain fatty acids from blood lipoproteins whenever they are needed for energy, even if the concentration of the lipoproteins is low The glycerol released from chylomicron TAG by LPL may be used for triacylglycerol synthesis in the liver in the fed state The portion of a chylomicron that remains in the blood after LPL action is known as a chylomicron remnant This remnant binds to receptors on hepatocytes which recognize apo E, and is endocytosed

88
Transfer of Proteins Between
Lysosomes fuse with the endocytic vesicles, and the chylomicron remnants are degraded by lysosomal enzymes The products of lysosomal digestion (e.g., fatty acids, amino acids, glycerol, cholesterol, phosphate) can be reused As LPL is bound to the capillary endothelium through binding to proteoglycans, heparin also can bind to LPL and dislodge it from the capillary wall. This leads to loss of LPL activity and an increase of triglyceride content in the blood Transfer of Proteins Between HDL and Chylomicrons
 can be reused. As LPL is bound to the capillary endothelium through binding to proteoglycans, heparin also can bind to LPL and dislodge it from the capillary wall. This leads to loss of LPL activity and an increase of triglyceride content in the blood. Transfer of Proteins Between. HDL and Chylomicrons.")
89
The Metabolism of Chylomicrons

90
Fate of VLDL The TAG, which is produced in the smooth endoplasmic reticulum of the liver, is packaged with cholesterol, phospholipids, and proteins (synthesized in the rough endoplasmic reticulum) to form VLDL The microsomal triglyceride transfer protein (MTP), which is required for chylomicron assembly, is also required for VLDL assembly The major protein of VLDL is apoB-100. There is one long apoB-100 molecule wound through the surface of each VLDL particle ApoB-100 is encoded by the same gene as the apoB-48 of chylomicrons, but is a longer protein In intestinal cells, RNA editing produces a smaller mRNA and, thus, a shorter protein, apoB-48 VLDL is processed in the Golgi complex and secreted into the blood by the liver. The fatty acid residues of the triacylglycerols ultimately are stored in the triacylglycerols of adipose cells
 to form VLDL. The microsomal triglyceride transfer protein (MTP), which is required for chylomicron assembly, is also required for VLDL assembly. The major protein of VLDL is apoB-100. There is one long apoB-100 molecule wound through the surface of each VLDL particle. ApoB-100 is encoded by the same gene as the apoB-48 of chylomicrons, but is a longer protein. In intestinal cells, RNA editing produces a smaller mRNA and, thus, a shorter protein, apoB-48. VLDL is processed in the Golgi complex and secreted into the blood by the liver. The fatty acid residues of the triacylglycerols ultimately are stored in the triacylglycerols of adipose cells.")
91
In comparison to chylomicrons , VLDL particles are more dense, as they contain a lower percentage of triglyceride than do the chylomicrons Similar to chylomicrons, VLDL particles are first synthesized in a nascent form, and on entering the circulation they acquire apoproteins CII and E from HDL particles to become mature VLDL particles LPL cleaves the TAG in both VLDL and chylomicrons, forming fatty acids and glycerol The C-II apoprotein, which these lipoproteins obtain from HDL, activates LPL The low Km of the muscle LPL isozyme permits muscle to use the fatty acids of chylomicrons and VLDL as a source of fuel even when the blood concentration of these lipoproteins is very low. The isozyme in adipose tissue has a high Km and is most active after a meal, when blood levels of chylomicrons and VLDL are elevated

92
The fate of the VLDL particle after TAG has been removed by LPL is the generation of an IDL particle (intermediate-density lipoprotein), which can further lose triglyceride to become an LDL particle (low-density lipoprotein) The fate of the IDL and LDL particles is closely related with the metabolism of cholesterol A VLDL Particle

93
The Synthesis of VLDL

94
The Metabolism of VLDL (and Chylomicrons)
")
95
The Mobilization of Adipose Tracylglycerols
Hormone-sensitive lipase (HSL) is a key enzyme involved in the hydrolysis of adipocyte triacylglycerol Its name reflects the fact that the enzyme is activated by the signal transduction cascade involving cAMP and protein kinase A (PKA) In adipocytes, activation of PKA is initiated primarily by epinephrine and glucagon For decades, hormone-sensitive lipase was considered to be the key regulatory enzyme in the lipolysis pathway of adipose tissue However, it is now believed that a recently discovered lipase called desnutrin or adipose triglyceride lipase (ATGL) catalyzes the first step in triacylglycerol hydrolysis Desnutrin catalyzes the hydrolysis of TAG to diacylglycerols (DAG) and is the rate-limiting step in triacylglycerol hydrolysis
 is a key enzyme involved in the hydrolysis of adipocyte triacylglycerol. Its name reflects the fact that the enzyme is activated by the signal transduction cascade involving cAMP and protein kinase A (PKA) In adipocytes, activation of PKA is initiated primarily by epinephrine and glucagon. For decades, hormone-sensitive lipase was considered to be the key regulatory enzyme in the lipolysis pathway of adipose tissue. However, it is now believed that a recently discovered lipase called desnutrin or adipose triglyceride lipase (ATGL) catalyzes the first step in triacylglycerol hydrolysis. Desnutrin catalyzes the hydrolysis of TAG to diacylglycerols (DAG) and is the rate-limiting step in triacylglycerol hydrolysis.")
96
Hormone-sensitive lipase is more active against DAG than against TAG
Thus, the current hypothesis regarding the lipolytic cascade in adipocytes involves three esterases acting sequentially: desnutrin hydrolyzes the first ester bond in TAG generating DAG; then DAG are hydrolyzed by hormone-sensitive lipase to produce 2-monoacylglycerols; and finally, monoacylglycerol lipase removes the third fatty acid to produce glycerol

97
The Regulation of Triacylglycerol Metabolism
Insulin is the most important regulator of triacylglycerol metabolism Insulin enhances the rate of hydrolysis of lipoprotein-associated TAG by stimulating the synthesis and secretion of lipoprotein lipase by adipocytes Insulin also promotes TAG storage in adipocytes and TAG synthesis and VLDL export from hepatocytes Simultaneously, insulin inhibits the breakdown of TAG, in fat cells By contrast, hydrocortisone, epinephrine, and growth hormone oppose the action of insulin, inhibiting the synthesis of fatty acids in both hepatocytes and adipocytes, and promoting lipolysis in adipocytes in times of energy need, such as fasting and exercise

98
Regulation of TAG Synthesis
Insulin stimulates dephosphorylation and activation of acetyl-CoA carboxylase Insulin also promotes fatty acid synthesis by inducing enzymes of the fatty acid synthesis family In addition, insulin stimulates the catabolism of excess dietary carbohydrates, thereby increasing the supply of acetyl-CoA substrate for fatty acid and thus triacylglycerol synthesis Regulation of Adipocyte TAG Mobilization Lipolysis in adipocytes is under tight hormonal control The synthesis of desnutrin is induced by hydrocortisone and inhibited by insulin HSL is activated by epinephrine via a mechanism involving cAMP-dependent phosphorylation By contrast, insulin acts to dephosphorylate HSL, thereby inhibiting lipolysis

99
Adipocytes store TAG in the form of lipid droplets surrounded by a protein called perilipin
Like HSL, perilipin is phosphorylated by PKA In its unphosphorylated state, perilipin acts as a barrier that limits access of lipases to their substrates, thus maintaining a low rate of basal triacylglycerol hydrolysis Phosphorylation of perilipin causes the lipid droplets to fragment and disperse, permitting efficient hydrolysis of adipocyte TAG Regulation of Lipoprotein Lipase Activity LPL is active in both the fasted and fed states In the fasted state, it plays an important role in making fatty acids from VLDL TAG available to cardiac and skeletal muscles, In contrast, in the fed state, LPL directs fatty acids from both chylomicrons and VLDL to adipocytes for storage Adipocyte LPL expression is reduced during fasting while its expression in muscle is up-regulated

100
Conversely, adipocyte LPL expression is up-regulated in the fed state
In addition, muscle- and adipocyte-specific forms of LPL have different kinetic properties, with the muscle enzyme having a lower Km , for triacylglycerol than the adipocyte enzyme Thus, the active site of the LPL enzyme, which is localized to the surface of muscle capillaries, is saturated even during the fasted state, when circulating triacylglycerol-containing lipoprotein levels are low By contrast, the activity of LPL associated with adipose tissue capillaries increases in the fed state, when the levels of triacylglycerol-rich lipoproteins are relatively high The Triacylglycerol Cycle An additional factor in the balance between biosynthesis and degradation of triacylglycerols is that approximately 75% of all fatty acids released by lipolysis are reesterified to form TAG rather than used for fuel

101
This ratio persists even under starvation conditions, when energy metabolism is shunted from the use of carbohydrate to the oxidation of fatty acids Some of this fatty acid recycling takes place in adipose tissue, with the reesterification occurring before release into the bloodstream Some takes place via a systemic cycle in which free fatty acids are transported to liver, recycled to TAG, exported again to the adipose in the form of VLDL Flux through this triacylglycerol cycle between adipose tissue and liver may be quite low when other fuels are available and the release of fatty acids from adipose tissue is limited But the proportion of released fatty acids that are reesterified remains roughly constant at 75% under all metabolic conditions The function of the apparently futile triacylglycerol cycle is not well understood

102
One possibility is that, the excess capacity in the triacylglycerol cycle (the fatty acid that is eventually reconverted to TAG rather than oxidized as fuel) could represent an energy reserve in the bloodstream during fasting, one that would be more rapidly mobilized in a “fight or flight response” emergency than would stored TAG The glycerol 3-phosphate needed for the recycling of TAG in the starved state is provided by glyceroneogenesis Glyceroneogenesis is a shortened version of gluconeogenesis, from pyruvate to DHAP followed by conversion of the DHAP to glycerol 3- phosphate by cytosolic NAD+-linked glycerol 3-phosphate dehydrogenase In the adipose tissue, glyceroneogenesis coupled with reesterification of free fatty acids controls the rate of fatty acid release to the blood In addition, glyceroneogenesis in the liver alone can support the synthesis of enough G 3-P for 65% of reesterification

103
Flux through the triacylglycerol cycle between liver and adipose tissue is controlled to a large degree by the activity of PEP carboxykinase, which limits the rate of both gluconeogenesis and glyceroneogenesis Glucocorticoid hormones such as cortisol and dexamethasone (a synthetic glucocorticoid) regulate the levels of PEP carboxykinase reciprocally in the liver and adipose tissue These steroid hormones increase the expression of the gene encoding PEP carboxykinase in the liver, thus increasing gluconeogenesis and glyceroneogenesis Stimulation of glyceroneogenesis leads to an increase in the synthesis of triacylglycerol molecules in the liver and their release into the blood At the same time, glucocorticoids suppress the expression of the gene encoding PEP carboxykinase in adipose tissue This results in a decrease in glyceroneogenesis in adipose tissue
 regulate the levels of PEP carboxykinase reciprocally in the liver and adipose tissue. These steroid hormones increase the expression of the gene encoding PEP carboxykinase in the liver, thus increasing gluconeogenesis and glyceroneogenesis. Stimulation of glyceroneogenesis leads to an increase in the synthesis of triacylglycerol molecules in the liver and their release into the blood. At the same time, glucocorticoids suppress the expression of the gene encoding PEP carboxykinase in adipose tissue. This results in a decrease in glyceroneogenesis in adipose tissue.")
104
Recycling of fatty acids declines as a result and more free fatty acids are released into the blood
Thus glyceroneogenesis is regulated reciprocally in the liver and adipose tissue, affecting lipid metabolism in opposite ways A lower rate of glyceroneogenesis in adipose tissue leads to more fatty acid release (rather than recycling), whereas a higher rate in the liver leads to more synthesis and export of triacylglycerols High levels of free fatty acids in the blood interfere with glucose utilization in muscle and promote the insulin resistance that leads to type 2 diabetes A class of drugs called thiazolidinediones have been shown to reduce the levels of fatty acids circulating in the blood and increase sensitivity to insulin Thiazolidinediones bind to and activate a nuclear hormone receptor called peroxisome proliferator activated receptor γ (PPAR γ), leading to the induction in adipose tissue PEPCK
, whereas a higher rate in the liver leads to more synthesis and export of triacylglycerols. High levels of free fatty acids in the blood interfere with glucose utilization in muscle and promote the insulin resistance that leads to type 2 diabetes. A class of drugs called thiazolidinediones have been shown to reduce the levels of fatty acids circulating in the blood and increase sensitivity to insulin. Thiazolidinediones bind to and activate a nuclear hormone receptor called peroxisome proliferator activated receptor γ (PPAR γ), leading to the induction in adipose tissue PEPCK.")
105
A higher activity of PEPCK then leads to increased synthesis of the precursors of glyceroneogenesis
The therapeutic effect of thiazolidinediones is thus due, in part, to the increase in glyceroneogenesis, which in turn increases the resynthesis of triacylglycerol in adipose tissue and reduces the release of free fatty acid from adipose tissue into the blood The TAG Cycle

106
Diseases Involving Abnormalities in Triacylglycerol Metabolism
Obesity The amount of fat an individual can store depends on the number of fat cells in the body and the amount of triacylglycerol each cell can accommodate In obese individuals, both the number of fat cells and the size of the cells (i.e., the total storage capacity) is greater than in individuals with no history of obesity Fat cells begin to proliferate early in life, starting in the third trimester of gestation Proliferation essentially ceases before puberty, and thereafter fat cells change mainly in size However, some increase in the number of fat cells can occur in adulthood if preadipocytes are induced to proliferate by growth factors and changes in the nutritional state Weight reduction results in a decrease in the size of fat cells rather than a decrease in number
 is greater than in individuals with no history of obesity. Fat cells begin to proliferate early in life, starting in the third trimester of gestation. Proliferation essentially ceases before puberty, and thereafter fat cells change mainly in size. However, some increase in the number of fat cells can occur in adulthood if preadipocytes are induced to proliferate by growth factors and changes in the nutritional state. Weight reduction results in a decrease in the size of fat cells rather than a decrease in number.")
107
There are two major types of fat
There are two major types of fat. One is subcutaneous fat that is located just below the skin surface, most noticeable in the thighs, buttocks, arms and face The other is visceral fat which lies deep within the abdominal cavity and is responsible for the size of the waistline One way to predict if someone has too much body fat is to determine their body mass index (BMI) using a ratio of their weight and height It is generally accepted that a BMI value of less than 18.5 is considered underweight, is within the normal weight range, is overweight, and greater than 30 is obese BMI values do not provide information about the relative amounts of visceral fat and subcutaneous fat stores One of the best ways to predict an individual's disease risk is to use both their BMI value and the circumference of their waist in relationship to the size of their hips
 using a ratio of their weight and height. It is generally accepted that a BMI value of less than 18.5 is considered underweight, is within the normal weight range, is overweight, and greater than 30 is obese. BMI values do not provide information about the relative amounts of visceral fat and subcutaneous fat stores. One of the best ways to predict an individual s disease risk is to use both their BMI value and the circumference of their waist in relationship to the size of their hips.")
108
By determining a person's waist to hip ratio (WHR), it is possible to obtain an approximate measurement of the relative amounts of visceral and subcutaneous fat stored on their body A high WHR value corresponds to an "apple-shaped" body (more visceral fat in the waist than subcutaneous fat on the hips), whereas, a low WHR value leads to a "pear-shaped" body Body weight is determined by a balance between energy intake and energy expenditure The energy expenditure is required to maintain basal metabolic functions, absorption and digestion of foods , physical activity and, in children, linear growth and development If there is a net excess energy intake, BMI and WHR increase, eventually leading to overweight and ultimately to obesity and morbid obesity. In a population with stable genetic factors, increase in obesity is primarily attributable to consumption of excess food and decreased physical activity
, whereas, a low WHR value leads to a pear-shaped body. Body weight is determined by a balance between energy intake and energy expenditure. The energy expenditure is required to maintain basal metabolic functions, absorption and digestion of foods , physical activity and, in children, linear growth and development. If there is a net excess energy intake, BMI and WHR increase, eventually leading to overweight and ultimately to obesity and morbid obesity. In a population with stable genetic factors, increase in obesity is primarily attributable to consumption of excess food and decreased physical activity.")
109
Obesity is a worldwide health problem
Obesity is a worldwide health problem. It is a risk factor for development of diabetes mellitus, hypertension, and heart disease, all of which cause decreased quality of life and life expectancy Signals that initiate or inhibit feeding are extremely complex and include psychological and hormonal factors as well as neurotransmitter activity These signals are integrated and relayed through the hypothalamus Destruction of specific regions of the hypothalamus can lead to overeating and obesity or to anorexia and weight loss The adipocyte, in addition to storing triacylglycerol, secretes peptide hormones that regulate both glucose and fat metabolism The hormones leptin (Greek leptos, thin), resistin (resists insulin action), and adiponectin are all secreted from adipocytes under different conditions
, resistin (resists insulin action), and adiponectin are all secreted from adipocytes under different conditions.")
110
Leptin produced in adipocytes moves through the blood to the brain, where it acts on neuroendocrine system of the hypothalamus to curtail appetite (induce satiety) Leptin carries the message that fat reserves are sufficient, and it promotes a reduction in fuel intake and increased expenditure of energy Leptin-receptor interaction in the hypothalamus alters the release of neuronal signals to the region of the brain that affects appetite Leptin also stimulates the sympathetic nervous system, increasing blood pressure, heart rate, and thermogenesis by uncoupling electron transfer from ATP synthesis in the mitochondria of adipocytes Two types of neurons control fuel intake and metabolism. When leptin levels are low, the orexigenic (appetite-stimulating) neurons stimulate eating by producing and releasing neuropeptide Y (NPY)
 neurons stimulate eating by producing and releasing neuropeptide Y (NPY)")
111
NPY causes the next neuron in the circuit to send the signal to the brain, Eat!
When leptin levels are high, the anorexigenic (appetite-suppressing) neurons produce α-melanocyte-stimulating hormone (α-MSH) Release of α-MSH causes the next neuron in the circuit to send the signal to the brain, Stop eating! The amount of leptin released by adipose tissue depends on both the number and the size of adipocytes When weight loss decreases the mass of lipid tissue, leptin levels in the blood decrease, the production of NPY is increased Uncoupling is diminished, slowing thermogenesis and saving fuel, and fat mobilization slows Consumption of more food combined with more efficient utilization of fuel results in replenishment of the fat reserve in adipose, bringing the system back into balance
 neurons produce α-melanocyte-stimulating hormone (α-MSH) Release of α-MSH causes the next neuron in the circuit to send the signal to the brain, Stop eating! The amount of leptin released by adipose tissue depends on both the number and the size of adipocytes. When weight loss decreases the mass of lipid tissue, leptin levels in the blood decrease, the production of NPY is increased. Uncoupling is diminished, slowing thermogenesis and saving fuel, and fat mobilization slows. Consumption of more food combined with more efficient utilization of fuel results in replenishment of the fat reserve in adipose, bringing the system back into balance.")
112
The Mechanism of Action of Leptin

113
Human obesity may be the result of insufficient leptin production/action
Blood levels of leptin are usually much higher in obese animals (including humans) than in animals of normal body mass Some downstream element in the leptin response system may be defective in obese individuals, and the elevation in leptin could be the result of an (unsuccessful) attempt to overcome the leptin resistance In humans, adiponectin is secreted from adipocytes in inverse proportion to their adipose mass, lean individuals secreting more adiponectin than obese individuals This is the exact opposite of leptin secretion. Adiponectin circulates in the blood and powerfully affects the metabolism of fatty acids and carbohydrates in liver and muscle It increases the uptake of fatty acids from the blood by myocytes and the rate at which fatty acids undergo oxidation
 than in animals of normal body mass. Some downstream element in the leptin response system may be defective in obese individuals, and the elevation in leptin could be the result of an (unsuccessful) attempt to overcome the leptin resistance. In humans, adiponectin is secreted from adipocytes in inverse proportion to their adipose mass, lean individuals secreting more adiponectin than obese individuals. This is the exact opposite of leptin secretion. Adiponectin circulates in the blood and powerfully affects the metabolism of fatty acids and carbohydrates in liver and muscle. It increases the uptake of fatty acids from the blood by myocytes and the rate at which fatty acids undergo oxidation.")
114
It also blocks fatty acid synthesis and gluconeogenesis in hepatocytes, and it stimulates glucose uptake and catabolism in muscle and liver These effects of adiponectin occur indirectly, through activation of the key regulatory enzyme AMPK Mice with defective adiponectin genes are less sensitive to insulin than those with normal adiponectin, and they show poor glucose tolerance; ingestion of dietary carbohydrate causes a long-lasting rise in their blood glucose These metabolic defects resemble those of humans with type II diabetes, who also are insulin-insensitive and clear glucose from the blood only slowly Individuals with obesity or type II diabetes have lower blood adiponectin levels than non-diabetic controls Moreover, thiazolidinediones increase the expression of adiponectin mRNA in adipose tissue and increase blood adiponectin levels in experimental animals

115
The Mechanism of Action of Adiponectin
They also activate AMPK It appears that adiponectin, acting through AMPK, modulates the sensitivity of cells and tissues to insulin This hormone may prove to be one of the links between type II diabetes and its most important predisposing factor, obesity The Mechanism of Action of Adiponectin

116
The adipocytes have been shown to release a hormone known as resistin
This hormone may contribute to insulin resistance The mechanism by which resistin causes an insensitivity of cells to the actions of insulin is unknown Thiazolidinediones, suppress resistin transcription, reduce resistin levels, and increase sensitivity to insulin in these patients Proteins in a family of ligand-activated transcription factors, the peroxisome proliferator-activated receptors (PPARs), respond to changes in dietary lipid by altering the expression of genes involved in fat and carbohydrate metabolism These transcription factors were first recognized for their roles in peroxisome synthesis—thus their name Their normal ligands are fatty acids or fatty acid derivatives, but they can also bind synthetic agonists
, respond to changes in dietary lipid by altering the expression of genes involved in fat and carbohydrate metabolism. These transcription factors were first recognized for their roles in peroxisome synthesis—thus their name. Their normal ligands are fatty acids or fatty acid derivatives, but they can also bind synthetic agonists.")
117
PPARα, PPARδ, and PPARγ are members of the nuclear receptor superfamily
They act in the nucleus by forming heterodimers with another nuclear receptor, RXR (retinoid X receptor), binding to regulatory regions of DNA near the genes under their control and changing the rate of transcription of those genes PPAR γ , expressed primarily in liver and adipose tissue, is involved in turning on genes necessary to the differentiation of fibroblasts into adipocytes and genes that encode proteins required for lipid synthesis and storage in adipocytes The PPAR-y/RXR heterodimer regulates the transcription of genes encoding proteins such as acyl CoA-synthetases, lipoprotein lipase, GLUT4, mitochondrial uncoupling protein,… PPAR α in hepatocytes turns on the genes necessary for β-oxidation of fatty acids and formation of ketone bodies during fasting
, binding to regulatory regions of DNA near the genes under their control and changing the rate of transcription of those genes. PPAR γ , expressed primarily in liver and adipose tissue, is involved in turning on genes necessary to the differentiation of fibroblasts into adipocytes and genes that encode proteins required for lipid synthesis and storage in adipocytes. The PPAR-y/RXR heterodimer regulates the transcription of genes encoding proteins such as acyl CoA-synthetases, lipoprotein lipase, GLUT4, mitochondrial uncoupling protein,… PPAR α in hepatocytes turns on the genes necessary for β-oxidation of fatty acids and formation of ketone bodies during fasting.")
118
PPAR δ is a key regulator of fat oxidation, which acts by sensing changes in dietary lipid
It acts in liver and muscle, stimulating the transcription of at least nine genes encoding proteins for β-oxidation and for energy dissipation through uncoupling of mitochondria By stimulating fatty acid breakdown in uncoupled mitochondria, PPARδ causes fat depletion, weight loss, and thermogenesis Mode of Action of PPARs

119
Short-term eating behavior can be modified by hormones
Ghrelin is a peptide hormone produced in cells lining the stomach It is a powerful appetite stimulant that works on a shorter time scale (between meals) than leptin Ghrelin receptors are located in the pituitary gland (presumably mediating growth hormone release) and in the hypothalamus (affecting appetite), as well as in heart muscle and adipose tissue The concentration of ghrelin in the blood varies strikingly between meals, peaking just before a meal and dropping sharply just after the meal To summarize…. Although an increase in food intake beyond the daily requirements results in an increase in body weight and in fat stores, there is a large variation among individuals in the amount of weight gained for the excess calories consumed
 than leptin. Ghrelin receptors are located in the pituitary gland (presumably mediating growth hormone release) and in the hypothalamus (affecting appetite), as well as in heart muscle and adipose tissue. The concentration of ghrelin in the blood varies strikingly between meals, peaking just before a meal and dropping sharply just after the meal. To summarize…. Although an increase in food intake beyond the daily requirements results in an increase in body weight and in fat stores, there is a large variation among individuals in the amount of weight gained for the excess calories consumed.")
120
Both genetic and environmental factors influence the development of obesity
Still, although there are certainly wide variations in body structure and metabolism between individuals, the current epidemic of obesity in the many countries is due more to environmental than to genetic factors The efficient storage of excess calories as triacylglycerol may have been advantageous for our distant ancestors who were physically very active and for whom food scarcity was often the norm However, in the current context of sedentary lifestyles and in environments where there is an overabundance of calorically dense foods containing large amounts of fats and sugars, efficient fuel storage in the form of fat can result in obesity and its undesirable medical and social sequelae So, what should be done? MOVE!

121
Exercise Generating Maximum Power
ATP, phosphocreatine and anaerobic metabolism of muscular glycogen are the primary sources of energy High Intensity Endurance Exercise Phosphocreatine is the initial energy source; glycogenolysis ensues, anaerobically at first, with lactate production, but with increasing aerobic oxidation as oxygen availability increases Glycogen utilization is the major fuel for the first 15 minutes, but it is superseded by fatty acid and plasma glucose utilization During the exercise, branched-chain amino acid oxidation contributes some energy In endurance exercise, hepatic glycogen serves as a major fuel for muscle, and muscle glycogen is the initial fuel Low-Level Non-fatiguing Exercise This exercise is characteristic of an individual in normal occupational tasks that could reasonably be continued for long hours

122
The principal substrates used are similar to those in long endurance exercise but without depletion of phosphocreatine and with minimal muscle glycogen utilization The main source of energy is the aerobic oxidation of fatty acids, glucose, and branched-chain amino acids To the extent that one utilizes free fatty acids during exercise, one decreases fat stores proportionally Diabetes Mellitus Type I diabetes is caused by absent or insufficient insulin production Although the most common cause is autoimmune destruction of the β-cells of the pancreas, it can also result from chronic pancreatitis The resulting insulin deficiency has been described as “starvation in the midst of plenty,” with metabolic pathways active in fasting mode despite high plasma levels of nutrients in the blood

123
Diabetes affects lipid and protein metabolism as well as that of glucose
A lack of insulin results in fasting hyperglycemia with both overproduction of glucose by the liver and underutilization of glucose by both muscle and adipocytes There is going to be an increased catabolism of muscle proteins to provide gluconeogenetic precursors; hyperaminoacidemia will result Chronically raised levels of glucose in the blood bring damage to many organs, including the eyes, kidneys, blood vessels, and nerves The high glucagon/insulin ratio stimulates adipose triacylglycerol hydrolysis and increases the plasma free fatty acid concentration A great proportion of the fatty acids is going to travel to the liver and be metabolized in two main ways

124
Reesterification and release into the blood in the form of VLDL
But because insulin is lacking, LPL is not upregulated and hydrolysis of the triacylglycerols in chylomicrons and in VLDL is decreased, and hypertriglyceridemia results β-oxidation and formation of ketone bodies (since oxaloacetate is being diverted to gluconeogenesis) Higher than normal quantities of ketone bodies present in the blood or urine constitute ketonemia or ketonuria, respectively The overall condition is called ketosis The basic form of ketosis occurs in starvation and feeding of high-fat diets and involves depletion of available carbohydrate coupled with increased levels of free fatty acids This general pattern of metabolism is exaggerated to produce the pathologic states found in diabetes mellitus In uncontrolled diabetes mellitus, the concentration of plasma ketone bodies can rise to levels of 8 to 15 mmol/L or more (normal is in the range of 0.2–2 mmol/L)
 Higher than normal quantities of ketone bodies present in the blood or urine constitute ketonemia or ketonuria, respectively The overall condition is called ketosis. The basic form of ketosis occurs in starvation and feeding of high-fat diets and involves depletion of available carbohydrate coupled with increased levels of free fatty acids. This general pattern of metabolism is exaggerated to produce the pathologic states found in diabetes mellitus. In uncontrolled diabetes mellitus, the concentration of plasma ketone bodies can rise to levels of 8 to 15 mmol/L or more (normal is in the range of 0.2–2 mmol/L)")
125
Since both acetoacetate and β -hydroxybutyrate are organic acids, ketosis is a form of metabolic acidosis. In order to be excreted in the urine, the anionic metabolic acids in the urine must be counterbalanced by equivalent numbers of cations Therefore, ketosis may result in depletion of body stores of sodium and potassium and in some instances even in the loss of divalent cations such as calcium and magnesium The combined effect of the ketone bodies and the associated protons is known as diabetic ketoacidosis (DKA) Along with the effects of the hyperosmolarity of the blood (due to the excess glucose), DKA can lead to coma and death Currently, the only effective treatment for type I diabetes mellitus is insulin therapy Most people with diabetes mellitus have type II diabetes rather than type I, and it is type II diabetes that is now reaching epidemic incidence in many countries
 Along with the effects of the hyperosmolarity of the blood (due to the excess glucose), DKA can lead to coma and death. Currently, the only effective treatment for type I diabetes mellitus is insulin therapy. Most people with diabetes mellitus have type II diabetes rather than type I, and it is type II diabetes that is now reaching epidemic incidence in many countries.")
126
Type II diabetes mellitus is characterized by insulin resistance rather than primary insulin insufficiency A person has insulin resistance when larger-than-normal amounts of insulin are required to support insulin-dependent metabolic processes Insulin resistance is also commonly seen in the obese and in those with the metabolic syndrome In most instances of insulin resistance, insulin secretion is not impaired and insulin receptors are functional Although people in the early stages of insulin resistance can maintain normal blood glucose concentrations by increasing their insulin secretion, this compensation often becomes inadequate and they eventually progress to hyperglycemia and eventually to type II diabetes Like type I diabetes, type II diabetes is characterized by hyperglycemia, hypertriglyceridemia, hyperaminoacidemia, and elevated levels of free fatty acids

127
The high levels of free fatty acids in the blood are the result of increased TAG lipolysis by adipocytes Elevated free fatty acid levels, in turn, result in increased TAG synthesis by the liver and export of TAG-rich VLDL particles Unlike people with type I diabetes those with type II diabetes usually do not develop ketoacidosis, in part because - at least in the early stages of the disease - the liver is less insulin resistant than skeletal muscle or adipocytes Obesity often leads to insulin resistance in muscle In people with insulin resistance, the muscle cells do not sufficiently up-regulate the acyltransferases involved in triacylglycerol synthesis to cope with the increased availability of free fatty acids As a result, a high intracellular concentration of metabolic intermediates inhibits glucose uptake and glycolysis by muscle cells, and higher levels of insulin are required for glucose utilization

128
Although the mechanisms have not been fully elucidated, the relatively insulin-resistant state associated with obesity may be the result of an imbalance in adipokine production as well as elevated plasma levels of free fatty acids released from the excess adipocyte stores Exercise stimulates both TAG and glycogen synthesis in skeletal muscle and improves insulin sensitivity in both normal-weight and obese persons Among the mechanisms involved are up-regulation of GLUT4 transporters and induced expression of diacylglycerol acyltransferase Type II diabetes can be treated with a variety of drugs that stimulate insulin secretion (e.g., sulfonylureas) or increase insulin sensitivity (e.g, thiazolidinediones), or reduce hepatic gluconeogenesis (e.g., metformin) Many cases of type II diabetes, however, eventually progress to the point of pancreatic β-cell failure and dependence on insulin
 or increase insulin sensitivity (e.g, thiazolidinediones), or reduce hepatic gluconeogenesis (e.g., metformin) Many cases of type II diabetes, however, eventually progress to the point of pancreatic β-cell failure and dependence on insulin.")
129
Ethanol Metabolism Since there is no significant renal or pulmonary excretion of ethanol and no storage of ethanol in the body, whatever ethanol is consumed must be disposed of through metabolism The major site of alcohol metabolism is the liver, which contains both alcohol dehydrogenase (ADH) and the microsomal ethanol-oxidizing system (MEOS), the two enzymes most responsible for ethanol metabolism However, alcohol dehydrogenase activity is also present in the gastric mucosa (more so in men than women), and to a lesser extent in other organs, including the kidneys, lungs and small intestine ADH is a non-specific enzyme, able to use a wide range of simple alcohols, hydroxysteroids and retinol as substrates It also acts on ethanol produced by the fermentation of sugars by microorganisms in the colon
 and the microsomal ethanol-oxidizing system (MEOS), the two enzymes most responsible for ethanol metabolism. However, alcohol dehydrogenase activity is also present in the gastric mucosa (more so in men than women), and to a lesser extent in other organs, including the kidneys, lungs and small intestine. ADH is a non-specific enzyme, able to use a wide range of simple alcohols, hydroxysteroids and retinol as substrates. It also acts on ethanol produced by the fermentation of sugars by microorganisms in the colon.")
130
The overall pathway for metabolizing ethanol involves oxidation of the alcohol to acetaldehyde, which is then oxidized to acetate The acetate derived from ethanol oxidation is activated to acetyl-CoA by acetate thiokinase The resulting acetyl-CoA can be metabolized through the TCA cycle or utilized for fatty acid synthesis. There are three enzymes or enzyme systems that convert ethanol to acetaldehyde: ADH, MEOS, and catalase ADH NAD+-dependent ADH, which is a cytosolic enzyme converts ethanol to acetaldehyde with the concomitant release of NADH The NADH can be transported to the mitochondria and be used for ATP synthesis The gastric isozyme of ADH has a higher Km for ethanol than do the ADH isozymes in liver

131
MEOS The liver has a second pathway for oxidizing ethanol, which even though it can oxidize a variety of compounds in addition to ethanol, is designated the microsomal ethanol-oxidizing system Other substrates for MEOS include fatty acids, steroids, and certain drugs Ethanol and NADPH both donate electrons in the reaction, which reduces O2 to 2H2O The cytochrome P450 enzymes all have two major catalytic protein components: an electron-donating reductase system that transfers electrons from NADPH (cytochrome P450 reductase) and a cytochrome P450 The cytochrome P450 protein contains the binding sites for O2 and the substrate (e.g., ethanol) and carries out the reaction MEOS is part of the superfamily of cytochrome P450 enzymes, all of which catalyze similar oxidative reactions
 and a cytochrome P450. The cytochrome P450 protein contains the binding sites for O2 and the substrate (e.g., ethanol) and carries out the reaction. MEOS is part of the superfamily of cytochrome P450 enzymes, all of which catalyze similar oxidative reactions.")
132
Within the superfamily, at least 10 distinct gene families are found in mammals
More than 100 different cytochrome P450 isozymes exist within these 10 gene families Each isoenzyme has a distinct classification according to its structural relationship with other isoenzymes The isoenzyme that has the highest activity toward ethanol is called CYP2E1 A great deal of overlapping specificity exists among the various P450 isoenzymes, and ethanol is also oxidized by several other P450 isoenzymes “MEOS” refers to the combined ethanol oxidizing activity of all the P450 enzymes CYP2E1 has a much higher Km for ethanol than ADH. Thus, a greater proportion of ingested ethanol is metabolized through CYP2E1 at high levels of ethanol consumption than at low levels

133
The General Structure of
Cytochrome P450 Enzymes Catalase Catalase is also capable of oxidizing ethanol; however, its contribution to ethanol metabolism is minimal Oxidation of ethanol by catalase utilizes hydrogen peroxide: ethanol + H2O2 → acetaldehyde + 2H2O Acetaldehyde generated by ADH, MEOS, or catalase is oxidized to acetate by NAD+-dependent aldehyde dehydrogenase Although the major isozyme of aldehyde dehydrogenase is located in the mitochondria, there is also a cytosolic isozyme

134
acetate + CoASH + ATP → acetyl-CoA + AMP + PPi
The acetate produced by the oxidation of ethanol is activated by acetyl-CoA synthetase (acetate thiokinase) acetate + CoASH + ATP → acetyl-CoA + AMP + PPi The major liver isozyme of acetate thiokinase is cytosolic, and the acetyl-CoA it generates is used for fatty acid and cholesterol synthesis However, when these two pathways are inactive (due primarily to a high ratio of glucagon to insulin), acetate will diffuse out of the hepatocytes and be taken up and oxidized by heart and skeletal muscle which have high concentrations of mitochondrial acetyl-CoA synthetase Thus, if ethanol is consumed along with significant amounts of carbohydrate, the acetate generated from ethanol will be used mainly as a substrate for hepatic fatty acid synthesis If, however, ethanol is consumed in the absence of carbohydrate, the acetate derived from the oxidation of ethanol will be used mostly as fuel
 acetate + CoASH + ATP → acetyl-CoA + AMP + PPi. The major liver isozyme of acetate thiokinase is cytosolic, and the acetyl-CoA it generates is used for fatty acid and cholesterol synthesis. However, when these two pathways are inactive (due primarily to a high ratio of glucagon to insulin), acetate will diffuse out of the hepatocytes and be taken up and oxidized by heart and skeletal muscle which have high concentrations of mitochondrial acetyl-CoA synthetase. Thus, if ethanol is consumed along with significant amounts of carbohydrate, the acetate generated from ethanol will be used mainly as a substrate for hepatic fatty acid synthesis. If, however, ethanol is consumed in the absence of carbohydrate, the acetate derived from the oxidation of ethanol will be used mostly as fuel.")
135
Regulation of Ethanol Metabolism
Chronic consumption of ethanol can increase hepatic levels of CYP2E1 many fold When induction of MEOS increases the rate of metabolism of ethanol, the increased production of acetaldehyde may exceed the ability of the acetaldehyde dehydrogenases to further oxidize acetaldehyde Gender differences and genetic variants in the enzymes responsible for metabolizing ethanol may account for some of the individual variation in tolerance to ethanol As noted earlier, women normally have lower levels of gastric ADH than men The lower level of gastric ADH activity in women, as well as gender-based differences in body size and total body-water space, are believed to account for the lower tolerance to ethanol in women relative to men

136
A number of genetic polymorphisms in ethanol-metabolizing enzymes have been characterized
The inducibility of CYP2E1 can vary as much as 10-fold between persons Similarly, many persons of East Asian descent have an inactive or less active form of ALDH2, the hepatic mitochondrial isozyme of acetaldehyde dehydrogenase When people with mutation in ALDH2 consume ethanol, they are more susceptible to flushing, headache, and nausea, apparently because of acetaldehyde accumulation The drug Antabuse (disulfiram), which is used to discourage alcoholics from drinking, acts by inhibiting acetaldehyde dehydrogenase; people who consume ethanol while taking Antabuse develop symptoms similar to those of people who have a genetic lack of ALDH2 activity
, which is used to discourage alcoholics from drinking, acts by inhibiting acetaldehyde dehydrogenase; people who consume ethanol while taking Antabuse develop symptoms similar to those of people who have a genetic lack of ALDH2 activity.")
137
Toxic Effects of Alcohol Metabolism
Alcohol-induced liver disease, a common and sometimes fatal consequence of chronic ethanol abuse, may manifest itself in three forms: fatty liver, alcohol-induced hepatitis, and cirrhosis Each may occur alone, or they may be present in any combination in a given patient Acetaldehyde and free radicals generated from ethanol metabolism can result in alcohol-induced hepatitis, a condition in which the liver is inflamed and cells become necrotic and die However, ethanol ingestion also has acute effects on liver metabolism, including inhibition of fatty acid oxidation and stimulation of triacylglycerol synthesis, leading to a fatty liver It also can result in ketoacidosis or lactate acidosis and cause hypoglycemia Changes in Fatty Acid Metabolism The high NADH/NAD+ ratio generated from ethanol oxidation inhibits the oxidation of fatty acids in the liver

138
The source of the fatty acids can be dietary fat, fatty acids synthesized in the liver, or fatty acids released from adipose tissue stores These fatty acids are re-esterified into TAG by combining with glycerol 3-P The increased NADH/NAD+ ratio increases the availability of glycerol 3-P by promoting its synthesis from intermediates of glycolysis The TAG are incorporated into VLDL which accumulate in the liver and enter the blood, resulting in an ethanol-induced hyperlipidemia Fatty acids that are oxidized are converted to acetyl-CoA and subsequently to ketone bodies Enough NADH is generated from oxidation of ethanol and fatty acids that there is no need to oxidize acetyl-CoA in the TCA cycle. And the very high NADH/NAD+ ratio shifts the oxaloacetate in the TCA cycle to malate

139
The acetyl-CoA enters the pathway for ketone body synthesis instead of the TCA cycle
Although ketone bodies are being produced at a high rate, their metabolism in other tissues is restricted by the supply of acetate, which is the preferred fuel Thus, the blood concentration of ketone bodies may be much higher than found under normal fasting conditions Acetaldehyde Toxicity Accumulation of acetaldehyde, produced both by alcohol dehydrogenase and MEOS, is believed to be responsible for most of the alcohol-induced liver damage known as cirrhosis By virtue of its aldehyde group, acetaldehyde is a highly reactive molecule that can form adducts with many different intracellular proteins In particular, reaction of acetaldehyde with tubulin impairs secretion of serum proteins from hepatocytes, damaging these cells further

140
Increased oxidative stress, resulting from production of free radicals by CYP2E1, also contributes to liver damage in chronic alcoholics Acetaldehyde binds directly to glutathione and diminishes its ability to protect against H2O2 and prevent lipid peroxidation. It also binds to free radical defense enzymes Liver injury is irreversible at the stage that hepatic cirrhosis develops During the development of cirrhosis, many of the normal metabolic functions of the liver are lost, including biosynthetic and detoxification pathways Synthesis of blood proteins, including blood coagulation factors and serum albumin, is decreased. The capacity to incorporate amino groups into urea is decreased, resulting in the accumulation of toxic levels of ammonia in the blood. Conjugation and excretion of bilirubin (from heme degra-dation) is diminished, and bilirubin accumulates in the blood
 is diminished, and bilirubin accumulates in the blood.")
142
The Synthesis of Membrane Lipids
Glycerophospholipids The pathway for synthesis of glycerophospholipids, like that for TAG, starts with glycerol 3-phosphate Glycerol 3-phosphate reacts with two fatty acyl-CoA molecules to form phosphatidic acid Two different mechanisms are then used to add a head group to the molecule In the first mechanism, phosphatidic acid is cleaved by a phosphatase to form diacylglycerol (DAG) DAG then reacts with an activated head group. Head groups are activated by reacting them with CTP Phosphatidylcholine (PC) and phosphatidylethanolamine (PE) are synthesized using this mechanism In the second mechanism, phosphatidic acid reacts with CTP to form CDP-diacylglycerol
 DAG then reacts with an activated head group. Head groups are activated by reacting them with CTP. Phosphatidylcholine (PC) and phosphatidylethanolamine (PE) are synthesized using this mechanism. In the second mechanism, phosphatidic acid reacts with CTP to form CDP-diacylglycerol.")
143
CDP-DAG can react with phosphatidylglycerol (which itself is formed from the condensation of CDP-DAG and glycerol 3-phosphate) to produce cardiolipin or with inositol to produce phosphatidylinositol (PI) Human cells do not synthesize phosphatidylserine (PS) through any one of the above listed mechanisms Instead, PS is produced by a novel mechanism that involves polar head group exchange between PE and free serine PS can be converted back to PE by a decarboxylation reaction. PC can be synthesized from dietary choline when it is available If dietary choline is not available, choline can be synthesized from glucose But the rate of synthesis is inadequate to provide for the necessary amounts of choline. Thus, choline has been classified as an essential nutrient PE can also be methylated to form PC The donor of the methyl group is S-adenosylmethionine (SAM)
 through any one of the above listed mechanisms. Instead, PS is produced by a novel mechanism that involves polar head group exchange between PE and free serine. PS can be converted back to PE by a decarboxylation reaction. PC can be synthesized from dietary choline when it is available. If dietary choline is not available, choline can be synthesized from glucose. But the rate of synthesis is inadequate to provide for the necessary amounts of choline. Thus, choline has been classified as an essential nutrient. PE can also be methylated to form PC. The donor of the methyl group is S-adenosylmethionine (SAM)")
144
The Synthesis of PC, PE and PS

145
The Synthesis of PG, Cardiolipin and PI
Ether Lipids The pathway for synthesis of ether lipids starts with acylation of C-1 of the glycolytic intermediate DHAP The fatty acid is then replaced by a fatty alcohol The carbonyl group on C-2 of DHAP is then reduced by an NADPH dependent enzyme and acylated At this point, an alkyl analog of phosphatidate has been made

146
Head groups are added in a manner similar to that of PC
Formation of a double bond between carbons 1 and 2 of the alkyl group produces a plasmalogen Platelet-activating factor (PAF) is similar to choline plasmalogen except that an acetyl group replaces the fatty acyl group at carbon 2 of the glycerol moiety, and the alkyl group on carbon 1 is saturated PAF is released from phagocytic blood cells in response to various stimuli It causes platelet aggregation, edema, and hypotension, and it is involved in the allergic response Plasmalogen synthesis occurs within peroxisomes, and, in individuals with Zellweger’s syndrome , plasmalogen synthesis is compromised If severe enough, this syndrome leads to death at an early age
 is similar to choline plasmalogen except that an acetyl group replaces the fatty acyl group at carbon 2 of the glycerol moiety, and the alkyl group on carbon 1 is saturated. PAF is released from phagocytic blood cells in response to various stimuli. It causes platelet aggregation, edema, and hypotension, and it is involved in the allergic response. Plasmalogen synthesis occurs within peroxisomes, and, in individuals with Zellweger’s syndrome , plasmalogen synthesis is compromised. If severe enough, this syndrome leads to death at an early age.")
147
The Synthesis of Ethanolamine Plasmalogen

148
Sphingolipid Synthesis
The synthesis of sphingolipids begins with the formation of ceramide The starting materials for the synthesis of palmitoyl-CoA and serine Ceramide reacts with phosphatidylcholine to form sphingomyelin Ceramide also reacts with UDP-sugars to form cerebrosides (which contain a single monosaccharide, usually galactose or glucose) Galactocerebroside may react with 3-phosphoadenosine 5-phosphosulfate (PAPS, an active sulfate donor) to form sulfatides, the major sulfolipids of the brain Additional sugars may be added to ceramide to form globosides, and gangliosides are produced by the addition of N-acetylneuraminic acid (NANA) as branches from the oligosaccharide chains
 Galactocerebroside may react with 3-phosphoadenosine 5-phosphosulfate (PAPS, an active sulfate donor) to form sulfatides, the major sulfolipids of the brain. Additional sugars may be added to ceramide to form globosides, and gangliosides are produced by the addition of N-acetylneuraminic acid (NANA) as branches from the oligosaccharide chains.")
149
The Synthesis of Sphingolipids
from Ceramide

150
The sphingolipidoses (lipid storage diseases) are a group of inherited diseases that are caused by a genetic defect in the catabolism of lipids containing sphingosine They are part of a larger group of lysosomal disorders and exhibit several constant features: (1) Complex lipids containing ceramide accumulate in cells, particularly neurons, causing neurodegeneration and shortening the life span (2) The rate of synthesis of the stored lipid is normal (3) The enzymatic defect is in the lysosomal degradation pathway of sphingolipids (4) The extent to which the activity of the affected enzyme is decreased is similar in all tissues There is no effective treatment for many of the diseases, although some success has been achieved with enzyme replacement therapy and bone marrow transplantation in the treatment of Gaucher’s and Fabry’s diseases
 Complex lipids containing ceramide accumulate in cells, particularly neurons, causing neurodegeneration and shortening the life span. (2) The rate of synthesis of the stored lipid is normal. (3) The enzymatic defect is in the lysosomal degradation pathway of sphingolipids. (4) The extent to which the activity of the affected enzyme is decreased is similar in all tissues. There is no effective treatment for many of the diseases, although some success has been achieved with enzyme replacement therapy and bone marrow transplantation in the treatment of Gaucher’s and Fabry’s diseases.")
151
Examples of Sphingolipidoses

152
CHOLESTEROL METABOLISM
Cholesterol is a ubiquitous and essential component of mammalian cell membranes It is also present in small amounts in the outer membrane of mitochondria Cholesterol is especially abundant in myelinated structures of the central nervous system, with 25% of the body’s cholesterol located in the brain In contrast to plasma, where most of the circulating cholesterol exists esterified to a fatty acid, most cholesterol in cellular membranes is present in the free (unesterified) form The fluidity of membranes is determined by the degree of unsaturation of the hydrocarbon chains of the phospholipids and by the amount of cholesterol in the membrane Hydrocarbon chains with cis-double bonds produce kinks and allow a greater degree of freedom of movement for the neighboring alkyl side chains
 form. The fluidity of membranes is determined by the degree of unsaturation of the hydrocarbon chains of the phospholipids and by the amount of cholesterol in the membrane. Hydrocarbon chains with cis-double bonds produce kinks and allow a greater degree of freedom of movement for the neighboring alkyl side chains.")
153
Hence, these unsaturated chains give rise to more fluidity than do saturated alkyl chains, which associate in ordered arrays Cholesterol is packed between fatty alkyl chains, the ring bearing the polar hydroxyl group interacting with the polar groups of phospho- and glycolipids The presence of cholesterol disrupts the orderly stacking of alkyl side chains, restricts their mobility, and causes increased membrane viscosity Cholesterol is abundant in bile. The solubilization of free cholesterol in bile is achieved in part by the detergent property of phosphatidylcholine (lecithin), which is produced in liver and secreted into bile Bile acids, which are metabolites of cholesterol, also aid in solubilizing cholesterol in bile Increased biliary secretion of cholesterol or decreased secretion of phospholipids or bile acids into bile may lead to deposition of cholesterol-rich gallstones
, which is produced in liver and secreted into bile. Bile acids, which are metabolites of cholesterol, also aid in solubilizing cholesterol in bile. Increased biliary secretion of cholesterol or decreased secretion of phospholipids or bile acids into bile may lead to deposition of cholesterol-rich gallstones.")
154
Indeed, the name cholesterol was derived some 200 years ago from the Greek words chole (bile) + stereos (solid) A little more than half the cholesterol of the body arises by synthesis (about 700 mg/d), and the remainder is provided by the average diet Virtually all tissues containing nucleated cells are capable of cholesterol synthesis However , the liver and intestine account for approximately 10% each of total synthesis in humans In addition, the adrenal cortex, and reproductive tissues, including ovaries, testes, and placenta have got a high capacity of cholesterol synthesis Cholesterol synthesis occurs in the endoplasmic reticulum and the cytosol Humans cannot metabolize cholesterol to CO2 and water. Excretion of cholesterol and of bile acids synthesized from cholesterol occurs by way of the liver, gallbladder, and intestine
, and the remainder is provided by the average diet. Virtually all tissues containing nucleated cells are capable of cholesterol synthesis. However , the liver and intestine account for approximately 10% each of total synthesis in humans. In addition, the adrenal cortex, and reproductive tissues, including ovaries, testes, and placenta have got a high capacity of cholesterol synthesis. Cholesterol synthesis occurs in the endoplasmic reticulum and the cytosol. Humans cannot metabolize cholesterol to CO2 and water. Excretion of cholesterol and of bile acids synthesized from cholesterol occurs by way of the liver, gallbladder, and intestine.")
155
Cholesterol Synthesis
Acetyl-CoA is the source of all carbon atoms in cholesterol The biosynthesis of cholesterol may be divided into five steps: Step 1: Synthesis of Mevalonate HMG-CoA is formed by the reactions used in mitochondria to synthesize ketone bodies However, since cholesterol synthesis is extramitochondrial, the two pathways are distinct Initially, two molecules of acetyl-CoA condense to form acetoacetyl-CoA catalyzed by cytosolic thiolase Acetoacetyl-CoA condenses with an additional molecule of acetyl-CoA catalyzed by HMG-CoA synthase to form HMG-CoA, which is reduced to mevalonate by NADPH catalyzed by HMG-CoA reductase This is the rate-limiting step in the pathway of cholesterol synthesis

156
Step 2: Formation of Isoprenoid Units
Mevalonate is phosphorylated sequentially by ATP by three kinases, and after decarboxylation the active isoprenoid unit, isopentenyl diphosphate, is formed Step 3: Formation of Squalene Isopentenyl diphosphate is isomerized by a shift of the double bond to form dimethylallyl diphosphate, then condensed with another molecule of isopentenyl diphosphate to form the ten-carbon intermediate geranyl diphosphate A further condensation forms farnesyl diphosphate Step 1

157
Two molecules of farnesyl diphosphate condense at the diphosphate end to form squalene
The polyisoprenoids dolichol and ubiquinone are formed from farnesyl diphosphate by the further addition of up to 16 (dolichol) or 3–7 (ubiquinone) isopentenyl diphosphate residues Some GTP-binding proteins in the cell membrane are prenylated with farnesyl or geranylgeranyl (20 carbon) residues Protein prenylation is believed to facilitate the anchoring of proteins into lipoid membranes and may also be involved in protein-protein interactions and membrane-associated protein trafficking Step 4: Formation of Lanosterol Squalene can fold into a structure that closely resembles the steroid nucleus Step 5: Formation of Cholesterol The steroid nucleus and the side chain of lanosterol (30 C) are modified by at least eight enzymes to give cholesterol (27 C)
 or 3–7 (ubiquinone) isopentenyl diphosphate residues. Some GTP-binding proteins in the cell membrane are prenylated with farnesyl or geranylgeranyl (20 carbon) residues. Protein prenylation is believed to facilitate the anchoring of proteins into lipoid membranes and may also be involved in protein-protein interactions and membrane-associated protein trafficking. Step 4: Formation of Lanosterol. Squalene can fold into a structure that closely resembles the steroid nucleus. Step 5: Formation of Cholesterol. The steroid nucleus and the side chain of lanosterol (30 C) are modified by at least eight enzymes to give cholesterol (27 C)")
158
Step 2 Step 3

159
Two key enzymes can esterify cholesterol
One is an intracellular enzyme, acyl-CoA: cholesterol acyltransferase (ACAT), which transfers a fatty acyl group from its CoA carrier The other is an extracellular enzyme called lecithin:cholesterol acyltransferase (LCAT), which esterifies cholesterol with a fatty acyl derived from phosphatidylcholine Steps 4 and 5
, which transfers a fatty acyl group from its CoA carrier. The other is an extracellular enzyme called lecithin:cholesterol acyltransferase (LCAT), which esterifies cholesterol with a fatty acyl derived from phosphatidylcholine. Steps 4 and 5.")
160
The Transport of Cholesterol
The total fasting cholesterol concentration in plasma of healthy people is usually 150 to 200 mg per 100 mL, which is about twice the normal plasma glucose concentration Such a high concentration of cholesterol in plasma is possible due to cholesterol-rich plasma lipoproteins that solubilize and transport cholesterol Only about 30% of the total plasma cholesterol is free (unesterified); the rest is esterified with a long-chain fatty acid, usually linoleic acid, which increases the hydrophobicity of cholesterol Chylomicrons are the main vehicle for transporting dietary-derived cholesterol and cholesteryl esters As the chylomicrons circulate in the plasma, they lose TAG and shrink to become chylomicron remnants The chylomicron remnants then deliver cholesterol and other lipophilic molecules (e.g., fat-soluble vitamins) to the liver
; the rest is esterified with a long-chain fatty acid, usually linoleic acid, which increases the hydrophobicity of cholesterol. Chylomicrons are the main vehicle for transporting dietary-derived cholesterol and cholesteryl esters. As the chylomicrons circulate in the plasma, they lose TAG and shrink to become chylomicron remnants. The chylomicron remnants then deliver cholesterol and other lipophilic molecules (e.g., fat-soluble vitamins) to the liver.")
161
Although the main function of VLDL is to export endogenous TAG that are made in the liver, VLDL is also involved in the transport of both free cholesterol and cholesteryl esters between tissues VLDL are the main vehicle for exporting both dietary-derived and endogenously synthesized cholesterol from hepatocytes into the plasma Like chylomicrons, VLDL changes its composition and size as it circulates; it loses TAG through hydrolysis by LPL and by acquiring additional cholesteryl esters from HDL As their TAG component undergoes hydrolysis, VLDL particles become remnants of various sizes (sometimes called IDL, or intermediate-density lipoproteins) The VLDL remnants contain apo E; approximately two-thirds of the VLDL remnants are removed from the circulation by the liver
 The VLDL remnants contain apo E; approximately two-thirds of the VLDL remnants are removed from the circulation by the liver.")
162
The remaining IDL are converted in the circulation to LDL as a result of the actions of LPL and hepatic triacyglycerol lipase (HTGL) LDL is the major lipoprotein that transports cholesterol in blood Unlike chylomicrons and VLDL that are rich in TAG, the core of LDL contains primarily cholesteryl esters The surface of each LDL particle contains one molecule of apo B100 Since its concentration in plasma is positively correlated with cardiovascular disease (stroke, myocardial infarction, blood clots), LDL-cholesterol is popularly termed the “bad” cholesterol LDL functions primarily to deliver cholesterol and cholesteryl esters to peripheral tissues such as the adrenal glands, testes, and ovaries
, LDL-cholesterol is popularly termed the bad cholesterol. LDL functions primarily to deliver cholesterol and cholesteryl esters to peripheral tissues such as the adrenal glands, testes, and ovaries.")
163
Since LDL contains some cholesteryl esters derived from HDL, it also contributes to reverse cholesterol transport, whereby cholesteryl esters are transported from peripheral tissues to the liver for excretion as cholesterol or as bile salts Both hepatocytes and peripheral cells express LDL receptors which recognize apo B 100 and internalize LDL via receptor-mediated endocytosis The receptors are then recycled back to the cell surface, and the LDL is transported to lysosomes, where hydrolysis of cholesteryl esters generates free cholesterol The main role of HDL is reverse cholesterol transport whereby HDL extracts cholesterol from peripheral tissues and transports that cholesterol to the liver for excretion Circulating HDL can also donate cholesteryl esters to other lipoproteins such as VLDL and IDL HDL also plays a central role in lipoprotein metabolism by donating proteins such as apo C2 and apo E to chylo. and VLDL

164
LDL Metabolism

165
Since the concentration of HDL is inversely correlated with cardiovascular disease, HDL-cholesterol is described as the “good” cholesterol The primary apoprotein in HDL is apo A The major form of apo A is apo A 1 , which is synthesized by both the liver and the intestine There are three major structural forms of apo A1 circulating in the plasma: (1) amorphous or lipid-free HDL (apo A1), which does contain some phospholipid; (2) nascent or discoidal, lipid-poor HDL; and ( 3 ) mature, spherical HDL (HDL2, HDL3), which is rich in cholesteryl esters Lipid-free apo A1 is secreted by the liver and intestine and acquires free cholesterol and phospholipids in the plasma, thereby becoming nascent HDL As nascent HDL acquires additional free cholesterol, the cholesterol is esterified by the action of LCAT to generate cholesteryl esters –HDL2 is formed
 amorphous or lipid-free HDL (apo A1), which does contain some phospholipid; (2) nascent or discoidal, lipid-poor HDL; and ( 3 ) mature, spherical HDL (HDL2, HDL3), which is rich in cholesteryl esters. Lipid-free apo A1 is secreted by the liver and intestine and acquires free cholesterol and phospholipids in the plasma, thereby becoming nascent HDL. As nascent HDL acquires additional free cholesterol, the cholesterol is esterified by the action of LCAT to generate cholesteryl esters –HDL2 is formed.")
166
There are two mechanisms by which circulating HDL gives up some of its cholesteryl esters
One is the exchange of cholesteryl esters in HDL for TAG in VLDL, IDL, or, to a lesser extent, in LDL The exchange process is mediated by cholesteryl ester transfer protein (CETP) Cholesteryl esters may also be removed by selective uptake by the liver, which is mediated by a scavenger receptor class B1 (SR-B1) and occurs without the intracellular uptake of HDL proteins The resulting HDL3 particles are smaller and have a higher ratio of TAG to cholesteryl esters than HDL2 has, and can be cleared by the liver or the kidney In the process of HDL3 formation from HDL2 some of the excess phospholipid is released from the surface of the particles, thus regenerating lipid-poor HDL. The lipid-poor HDL can then acquire and esterify additional cholesterol
 Cholesteryl esters may also be removed by selective uptake by the liver, which is mediated by a scavenger receptor class B1 (SR-B1) and occurs without the intracellular uptake of HDL proteins. The resulting HDL3 particles are smaller and have a higher ratio of TAG to cholesteryl esters than HDL2 has, and can be cleared by the liver or the kidney. In the process of HDL3 formation from HDL2 some of the excess phospholipid is released from the surface of the particles, thus regenerating lipid-poor HDL. The lipid-poor HDL can then acquire and esterify additional cholesterol.")
167
ABCA-1 and ABCG-1 are members of the ATP-binding cassette transporter (ABC) family of transporters
LCAT acquired from the circulation and activated by apo A1 HDL Metabolism

168
Receptor-Mediated Endocytosis of Lipoproteins
Receptor-mediated endocytosis of lipoproteins provides a mechanism both for their clearance from the circulation and for the delivery of key lipid components to target cells Targeting of lipoproteins to sites of metabolism and removal is mediated primarily by the apoproteins on their surfaces The LDL receptor (LDLR) is a transmembrane glycoprotein with an apo B-100-binding domain LDL receptors are expressed on liver cells and extrahepatic tissues and they recognize apo B 100 but not the smaller apo B48 molecule present on chylomicrons and chylomicron remnants Once the LDL receptor is occupied by LDL, the LDL : LDLR complex clusters in coated pits, which are then internalized by receptor-mediated endocytosis Intracellularly, as the clathrin-coated vesicles lose their clathrin the LDL receptors are recycled back to the plasma membrane
 is a transmembrane glycoprotein with an apo B-100-binding domain. LDL receptors are expressed on liver cells and extrahepatic tissues and they recognize apo B 100 but not the smaller apo B48 molecule present on chylomicrons and chylomicron remnants. Once the LDL receptor is occupied by LDL, the LDL : LDLR complex clusters in coated pits, which are then internalized by receptor-mediated endocytosis. Intracellularly, as the clathrin-coated vesicles lose their clathrin the LDL receptors are recycled back to the plasma membrane.")
169
The LDL-containing endosomes then fuse with lysosomes to form endolysosomes
Within the endolysosomes, the cholesteryl esters are hydrolyzed by “acid lipase” to free cholesterol and fatty acids, while the apo B-100 is hydrolyzed to amino acids LDL Receptor-Related Protein (LRP) is expressed on the surface of hepatocytes but not peripheral cells. Its function is to bind and clear chylomicron remnants Scavenger Receptors A (SR-A’s) are a family of molecules that are expressed on tissue macrophages, Kupffer cells, and various extrahepatic endothelial cells Scavenger Receptors B 1 (SR-B1’s) are different in that they are not internalized by receptor-mediated endocytosis Instead, SR-B 1 permit hepatocytes (and the adrenal glands, ovaries, and other steroidogenic tissues) to selectively remove and internalize HDL-associated cholesteryl esters
 is expressed on the surface of hepatocytes but not peripheral cells. Its function is to bind and clear chylomicron remnants. Scavenger Receptors A (SR-A’s) are a family of molecules that are expressed on tissue macrophages, Kupffer cells, and various extrahepatic endothelial cells. Scavenger Receptors B 1 (SR-B1’s) are different in that they are not internalized by receptor-mediated endocytosis. Instead, SR-B 1 permit hepatocytes (and the adrenal glands, ovaries, and other steroidogenic tissues) to selectively remove and internalize HDL-associated cholesteryl esters.")
170
Receptor-Mediated Endocytosis of LDL

171
The Regulation of Cholesterol Metabolism
Regulation of HMG-CoA Reductase The activity of HMG-CoA reductase is under strict metabolic control The simultaneous regulation of HMG-CoA reductase synthesis and degradation can alter steady-state levels of the enzyme 200-fold Sterol regulatory element binding protein (SREBP) plays a central role in regulating the expression of HMG-CoA reductase levels SREBPs, after synthesis, are integral ER proteins associated with SCAP (SREBP cleavage-activating protein). The active component of the protein is released by two proteases, S1P (site 1 protease) and S2P. Once released, the active amino terminal component travels to the nucleus to bind to SREs. Cholesterol and cholesterol derivatives block the proteolytic activation and transport of SREBP
 plays a central role in regulating the expression of HMG-CoA reductase levels. SREBPs, after synthesis, are integral ER proteins associated with SCAP (SREBP cleavage-activating protein). The active component of the protein is released by two proteases, S1P (site 1 protease) and S2P. Once released, the active amino terminal component travels to the nucleus to bind to SREs. Cholesterol and cholesterol derivatives block the proteolytic activation and transport of SREBP.")
172
Transcription of the LDL receptor is also regulated by the intracellular cholesterol concentration through SREBP There are multiple isoforms of SREBP. One of these, SREBP-2, selectively activates transcription of cholesterol biosynthetic genes and the LDL receptor gene By contrast, SREBP-1 also activates transcription of acetyl-CoA carboxylase, fatty acid synthase,… SREBP-1 thus controls not only cholesterol synthesis but also the synthesis of fatty acids, TAG, and phospholipids In sterol-depleted cells, HMG-CoA reductase is slowly degraded with a half-life greater than 12 hours In the presence of abundant sterols the degradation of HMG-CoA reductase is accelerated HMG-CoA reductase is also inhibited when it is phosphorylated by AMP-activated kinase (AMPK) Glucagon and sterols activate AMPK while insulin favors the action of phosphatase
 Glucagon and sterols activate AMPK while insulin favors the action of phosphatase.")
173
Transcriptional Control
Regulation by Proteolysis Regulation by Phosphorylation The Regulation of HMG-CoA Reductase

174
Abnormalities in Cholesterol Metabolism
Hypercholesterolemia Hypercholesterolemia refers to plasma levels of cholesterol that exceed the normal range The risk of coronary heart disease (CHD) is correlated with LDL-cholesterol (LDL-C) level, while a high fasting HDL-C level is a negative risk factor for CHD Since the risk of developing CHD is related mainly to the LDL-component, treatment is aimed at decreasing the level of LDL Hypercholesterolemia could have genetic origins A deficiency of LDLR is the most common cause of familial hypercholesterolemia (FH) FH heterozygotes usually exhibit an elevated fasting plasma LDL-C concentration and a normal triglyceride level They have an increased risk of CHD, with onset in the fourth or fifth decade. Patients with heterozygous FH are generally responsive to treatment
 is correlated with LDL-cholesterol (LDL-C) level, while a high fasting HDL-C level is a negative risk factor for CHD. Since the risk of developing CHD is related mainly to the LDL-component, treatment is aimed at decreasing the level of LDL. Hypercholesterolemia could have genetic origins. A deficiency of LDLR is the most common cause of familial hypercholesterolemia (FH) FH heterozygotes usually exhibit an elevated fasting plasma LDL-C concentration and a normal triglyceride level. They have an increased risk of CHD, with onset in the fourth or fifth decade. Patients with heterozygous FH are generally responsive to treatment.")
175
Persons who are homozygous for FH usually have very high plasma LDL-C levels, even in early childhood These patients invariably have cutaneous deposits of cholesterol called xanthomas on the hands, wrists, elbows, and/or knees Coronary heart disease usually manifests within the first two decades of life Due to the lack of functional receptors, patients with homozygous FH are largely unresponsive to drug therapies Currently, the preferred treatment is the selective removal of VLDL, IDL and LDL from the plasma Atherogenic Dyslipidemia The dyslipidemic or atherogenic profile is a combination of three abnormalities in plasma lipoprotein levels: high VLDL triacylglycerol , low HDL-C, and the presence of relatively small, dense LDL particles

176
Independent of the concentration of either total plasma cholesterol or LDL-C, the dyslipidemic profile is a major risk factor for coronary artery disease The dyslipidemic profile is commonly associated with insulin resistance, type II diabetes, central obesity, and hypertension, which are included in a constellation of findings that have been termed the metabolic syndrome Tangier Disease The role of ABC 1-transporter in cholesterol efflux is exemplified by an autosomal recessive disease known as Tangier disease where mutations in the gene encoding the ABC-1 transporter lead to accumulation of cholesterol esters in the tissues with almost complete absence of HDL cholesterol Alzheimer's Disease Abnormal cholesterol metabolism may be a factor in Alzheimer’s disease (AD)
")
177
Of the several genotypes for apo E, the acquisition of two E4 alleles may increase the risk for Alzheimer's disease up to eightfold and shifts the onset to the lower ages Treatment of Hypercholesterolemia Treatment of persons with moderate hypercholesterolemia usually begins with dietary and other behavioral changes (e.g., exercise) Diets low in total fat, saturated fat, and cholesterol and relatively high in oleic acid (as in olive oil) tend to lower both the total cholesterol level and the LDL-C level If additional reductions in cholesterol are needed, various medications are available Statin drugs decrease cholesterol synthesis by inhibiting HMG-CoA activity. This, in turn, results in up-regulation of the LDLR, particularly in hepatocytes On a mass basis, the major metabolic products of cholesterol metabolism are the bile salts
 Diets low in total fat, saturated fat, and cholesterol and relatively high in oleic acid (as in olive oil) tend to lower both the total cholesterol level and the LDL-C level. If additional reductions in cholesterol are needed, various medications are available. Statin drugs decrease cholesterol synthesis by inhibiting HMG-CoA activity. This, in turn, results in up-regulation of the LDLR, particularly in hepatocytes. On a mass basis, the major metabolic products of cholesterol metabolism are the bile salts.")
178
Most bile salts are normally reabsorbed in the distal ileum
Soluble dietary fiber, such as that found in fruits and oat bran, binds bile salts and decrease their absorption This, in turn, increases synthesis of bile salts in the liver, consuming cholesterol in the process and decreasing the concentration of cholesterol in hepatocytes Bile-acid sequestrants are insoluble polymers that bind bile acids strongly inside the resin matrix; the bile acid-sequestrant complex is excreted in the feces Depletion of the body's bile acid pool results in the increased conversion of cholesterol to bile acids The depletion of hepatic cholesterol increases the expression of LDLR and lowers the plasma LDL-cholesterol concentration However, since the decreased intrahepatic concentration of cholesterol also stimulates synthesis of HMG-CoA reductase, statins may be prescribed in conjunction with resins

179
Plant sterols interfere with the absorption of dietary cholesterol
Plant sterols interfere with the absorption of dietary cholesterol. Increased intake of vegetables and therapeutic doses of plant sterols have been used to lower the plasma cholesterol level Cholesterol absorption can also be reduced by Ezetimibe which inhibits the transporter that moves cholesterol from the intestinal lumen into enterocytes In addition, niacin and a class of drugs known as fibrates increase HDL levels and lower plasma TAG levels The Development of Atherosclerosis Atherosclerosis is a degenerative condition of the arteries in which fat, yellowish plaque, known as atheroma, is present in medium and large arteries The current view is that atheroma starts with injury to endothelial cells in an artery and that his affects the immune system in a similar manner to the development of chronic inflammation. This is the injury– inflammation hypothesis

180
Many factors are suspected of causing injury to the endothelial cells, including pollutants, chemicals in tobacco smoke, various lipids (e.g. cholesterol, long-chain fatty acids), proinflammatory cytokines and bacteria Disturbance in the pattern of blood flow at bends and branchpoints in the major arteries can also lead to endothelial injury, particularly if the blood pressure is raised Such injuries result in the appearance of adhesion molecules on the surface of the endothelial cells to which monocytes adhere before entering the subendothelial space, where they develop into macrophages Factors that are released from these cells attract more monocytes and other immune cells, and the process of inflammation begins A central event in the generation of plaque is the uptake of LDL by macrophages in the subendothelial space. LDL enters this space through the damaged endothelial cells

181
The uptake occurs mainly by endocytosis mediated by the non-specific SR-A1 receptors
The uptake is not feedback regulated; LDL keep on entering the macrophage Damage to LDL prevents its recognition by the normal receptor and facilitates its uptake by the scavenger receptors Damage to LDL is caused by high blood glucose levels, which cause glycation of apo B 100, and free radicals, which oxidize unsaturated fatty acids in the phospholipids of the LDL Within the macrophage, the LDL is degraded and the resultant free cholesterol is esterified to form cholesterol ester This accumulates and then damages and eventually kills the macrophages to produce what are known as foam cells The dead and dying macrophages secrete cytokines and chemotactic agents, which encourage the entry of more monocytes and lymphocytes into the developing plaque

182
The process, therefore, has the characteristics of a vicious circle
They also secrete growth factors that stimulate proliferation of smooth muscle cells to further increase the size of the plaque The process, therefore, has the characteristics of a vicious circle Injury to Endothelial Cells

183
Derivatives of Cholesterol
Oxygenated derivatives of cholesterol play many roles in the body Cholesterol is the precursor of two important classes of molecules, bile acids and steroid hormones. In addition, 7-dehydrocholesterol, the immediate precursor of cholesterol, can be converted to cholecalciferol (vitamin D3), which ultimately produces 1,25- dihydroxycholecalciferol [ 1,25-(OH)2D3, calcitriol], the active hormone that regulates calcium metabolism Formation of Plaque
, which ultimately produces 1,25- dihydroxycholecalciferol [ 1,25-(OH)2D3, calcitriol], the active hormone that regulates calcium metabolism. Formation of Plaque.")
184
Many of the enzymes that catalyze oxygenation of cholesterol and cholesterol derivatives are monooxygenases (mixed-function oxidases) The monooxygenases that modify cholesterol are members of the cytochrome P450 superfamily of enzymes and usually utilize NADPH as a cofactor These monooxygenases are all membrane-bound and localized to either the endoplasmic reticulum or the inner mitochondrial membrane Cholesterol is a 27-carbon lipid containing a fused four-ring structure and a hydrocarbon chain Except for the one hydroxyl group at C3, cholesterol is completely non-polar By contrast, bile acids contain 24-carbon atoms and are more polar than cholesterol. The steroid ring of bile acids contains one or more additional hydroxyl groups and the shorter hydrocarbon side chain terminates in a carboxyl group

185
The so-called “bile salts” are actually bile acids that contain an amino acid which is conjugated in amide linkage to the side chain of the carboxyl group of the bile acid The two amino acids used most commonly by the liver to conjugate human bile acids are glycine, and the sulfur amino acid taurine Conjugated bile acids are more ionized at the pH of the intestinal lumen than their non-conjugated counterparts They are therefore better emulsifying agents Bile salts play a major role in the digestion and absorption of triacylglycerols and cholesteryl esters Bile salts emulsify dietary lipids in the gastrointestinal tract and stabilize the resulting mixed micelles Along with lecithin, bile salts solubilize the cholesterol and bile pigments present in bile, preventing formation of precipitates (stones) of cholesterol or bilirubin in the gallbladder and bile ducts
 of cholesterol or bilirubin in the gallbladder and bile ducts.")
186
In addition, formation of bile salts represents the major metabolic mechanism for eliminating excess cholesterol from the body The synthesis of the primary bile acids, cholic acid and chenodeoxycholic acid, and their conjugation with taurine or glycine to form bile salts occurs exclusively in the liver The conjugated bile acids are reabsorbed in intestine and transported through the blood back to the liver Intestinal bacteria deconjugate and dehydroxylate some of the bile salts; the products are called secondary bile acids The secondary bile acids are less soluble and, therefore, less readily reabsorbed from the intestinal lumen than the bile salts that have not been subjected to bacterial action Lithocholic acid, a secondary bile salt that has a hydroxyl group only at position 3, is the least soluble bile salt. Its major fate is excretion

187
Greater than 95% of the bile salts are resorbed in the ileum and return to the liver via the enterohepatic circulation In addition to secondary bile acids, the feces contains a mixture of cholesterol and cholesterol metabolites, such as cholestanol and coprostanol, generated by intestinal bacteria The Synthesis of Bile Acids

188
The Conjugation and Deconjugation of Bile Acids

189
Five types of hormones are produced by the oxidation and the removal of the side chain of cholesterol: glucocorticoids, mineralcorticoids, androgens, estrogens and progestins The synthesis of steroid hormones begins with the formation of pregnenolone through the action of the mitochondrial enzyme desmolase Desmolase removes 6 carbons from cholesterol (27 C) Pregnenolone moves from the mitochondria to the SER where it is changed to all the different types of steroid hormones The cholesterol used for steroid hormone synthesis is either synthesized in the tissues from acetyl CoA, extracted from intracellular cholesterol ester pools, or taken up by the cell in the form of cholesterol-containing lipoproteins (either internalized by the LDL-receptor, or absorbed by the SR-B1 receptor)
 Pregnenolone moves from the mitochondria to the SER where it is changed to all the different types of steroid hormones. The cholesterol used for steroid hormone synthesis is either synthesized in the tissues from acetyl CoA, extracted from intracellular cholesterol ester pools, or taken up by the cell in the form of cholesterol-containing lipoproteins (either internalized by the LDL-receptor, or absorbed by the SR-B1 receptor)")
190
The Synthesis of Steroid Hormones

191
Cholecalciferol (vitamin D3) can either be obtained from the diet or formed in the skin by the non-enzymatic action of ultraviolet (UV) light on 7-dehydrocholesterol Ergocalciferol (vitamin D2) is formed by the action of UV light on a structurally similar plant sterol (ergosterol) Vitamin D3 is preferred for vitamin D supplementation Conversion of cholecalciferol to its active hormonal form is a multiorgan process First, cholecalciferol is hydroxylated in the liver by 25-hydroxylase to form 25-hydroxycholecalciferol A second hydroxylation step, catalyzed by 1α-hydroxylase in the kidney, generates the active hormone 1,25- dihydroxycholecalciferol Kidney, bone, cartilage, and intestine contain a 24-hydroxylase that converts 25-dihydroxycholecalciferol to the inactive 24,25-dihydroxycholecalciferol,thus preventing formation of excess active calcitriol
 is formed by the action of UV light on a structurally similar plant sterol (ergosterol) Vitamin D3 is preferred for vitamin D supplementation. Conversion of cholecalciferol to its active hormonal form is a multiorgan process. First, cholecalciferol is hydroxylated in the liver by 25-hydroxylase to form 25-hydroxycholecalciferol. A second hydroxylation step, catalyzed by 1α-hydroxylase in the kidney, generates the active hormone 1,25- dihydroxycholecalciferol. Kidney, bone, cartilage, and intestine contain a 24-hydroxylase that converts 25-dihydroxycholecalciferol to the inactive 24,25-dihydroxycholecalciferol,thus preventing formation of excess active calcitriol.")
192
The Synthesis of Active Vitamin D

Similar presentations

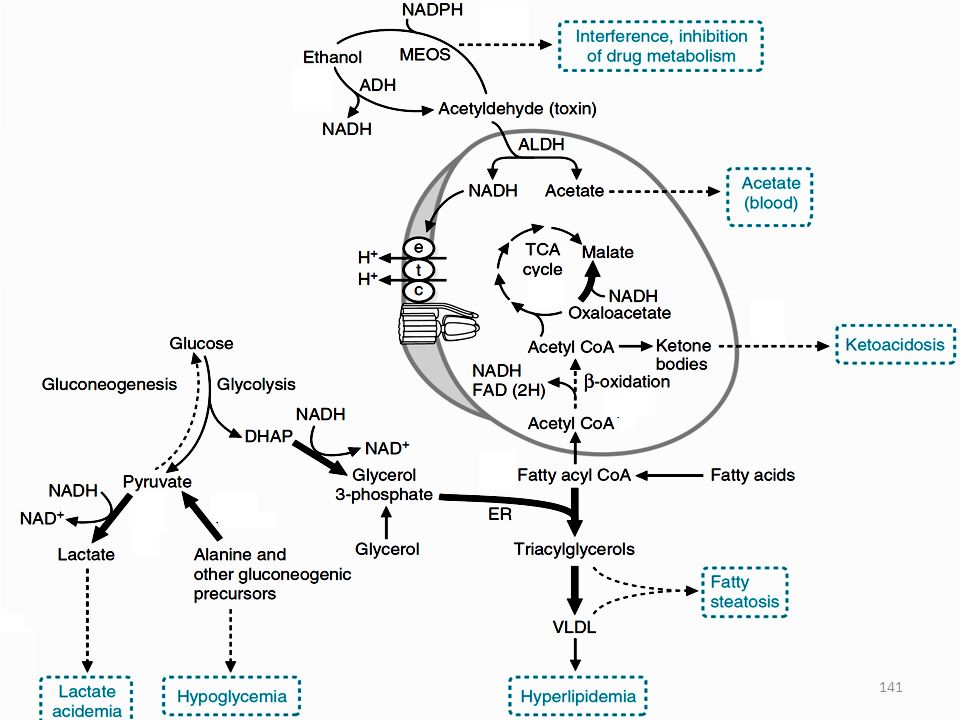


>")
Biosynthesis of glycerol. 2)Biosynthesis of fatty acids. 3)Biosynthesis of the triacylglycerol. It occurs in.>")









 Fatty acid Catabolism ( -oxidation)>")
>")
>")



