Download presentation
Presentation is loading. Please wait.

1
Seente geneetika II Heidi Tamm 5. märts 2010

2
Suguline paljunemine Seened mudelorganismidena Tetraadide analüüs Komplementatsiooni test S. cerevisiae genoom Proteoomika

3
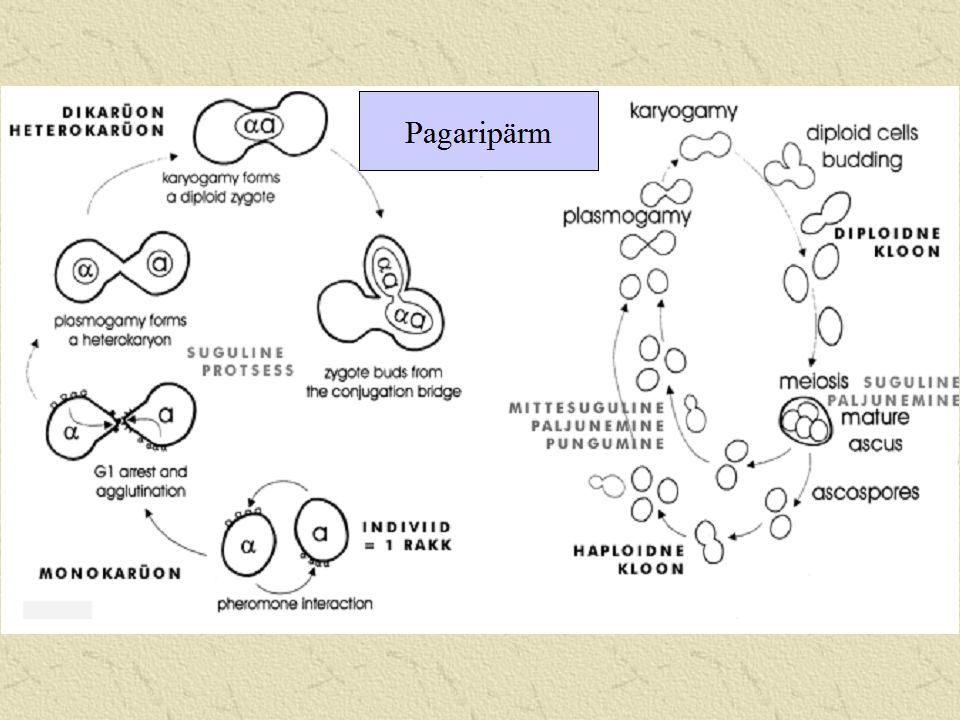
Suguline paljunemine Et saada maksimaalset kasu, peaksid ühinevate rakkude tuumad olema nii erinevad kui võimalik Tsütoplasmad peaksid olema nii sarnased kui võimalik (et vältida geneetilist saastust) Vegetatiivne vs. seksuaalne kokkusobivus

4
Isane või emane? ei ole võimalik eristada seened on hermafrodiitsed

5
Vegetatiivne kokkusobivus Heterokaarüonid tekivad vaid samasse vegetatiivse kokkusobivuse gruppi kuuluvate hüüfide vahel Kui hüüfid ei ole kokkusobivad, siis vahetult kontakteeruvad rakud surevad

6
Vegetatiivne kokkusobivus/sobimatus Liitumise kokkusobimatus (fusion incompatibility) - kokkusobivussüsteem määrab ära rakkude võime liituda. (hallitusseentel) Liitumisejärgne kokkusobimatus (post- fusion…) – määrab ära, kas rakud jäävad pärast liitumist ellu; kas koos tsütoplasmaga saavad migreeruda ka tuumad jm. organellid
 Liitumisejärgne kokkusobimatus (post- fusion…) – määrab ära, kas rakud jäävad pärast liitumist ellu; kas koos tsütoplasmaga saavad migreeruda ka tuumad jm. organellid.")
7
Ristumissüsteemid On määratud tuuma geenide poolt, mis takistavad kahe geneetiliselt identse mütseeli ristumist Heterotallism – süsteem, kus on takistatud ristumine kahe identse gameedi vahel Homotallism – kui miski ei takista identsete gameetide ristumist

8
Heterotallism anastomoos on võimalik ainult kahe erineva talluse vahel 90% kandseentest

9
Homotallism suguline paljunemine saab toimuda ilma interaktsioonita kahe talluse vahel iseviljastumine primaarne vs. sekundaarne homotallism

10
Primaarne homotallism idaneb ühetuumaline eos, moodustub kahetuumaline mütseel tuumad on ühesugused moodustuvad viljakehad

11
Sekundaarne homotallism kui peale meioosi satub ühte eosesse 2 tuuma

12
Ristumissüsteemid: unifaktoriaalne unifaktoriaalne ehk bipolaarne ristumist kontrollib üks lookus kahe alleeliga alleelide lahknemine meioosis tagab, et ühes eoses on vaid üks alleel. erinevad alleelid. Edukalt ristuvad vaid rakud, millel on ristumistüübi lookuses erinevad alleelid.

13
Ristumissüsteemid: unifaktoriaalne kui ühe viljakeha eostest kasvanud mütseele ristata omavahel, toimub dikaarüonide moodustumine 50% juhtudest kottseened, ikkesseened rooste- ja nõgiseened

14
Ristumissüsteemid: bifaktoriaalne Kaks omaette ristumistüübi lookust (A ja B). Bifaktoriaalne e. tetrapolaarne Edukaks ristumiseks on vaja erinevaid alleele mõlemas lookuses

15
Ristumissüsteemid: bifaktoriaalne Populatsioonis võib olla palju erinevaid alleele kummagi lookuse kohta 55% kandseentel Kui ühe viljakeha eostest kasvanud mütseele ristata omavahel, toimub dikaarüonide moodustumine 25% juhtudest

16
Ristumisfaktorid: bifaktoriaalne “A” geen kodeerib homeodomääni valke (transkriptsioonifaktoreid) “B” geen kodeerib feromone ja nende retseptoreid kummalgi geenil on alamlookused
 B geen kodeerib feromone ja nende retseptoreid kummalgi geenil on alamlookused")
17
S. cerevisiae ristumistüübi faktorid 2 ristumistüüpi: a ja α Ristumistüüp määratud: peptiidhormoonide (feromonide) poolt (α- ja a-faktorid) feromonispetsiifiliste retseptorite poolt
 poolt (α- ja a-faktorid) feromonispetsiifiliste retseptorite poolt.")
18
Kuidas ristumistüübi faktorid toimivad? Feromon seostub vastastüüpi raku pinnal oleva retseptoriga. Selle sündmuse tagajärjed: rakus algab aglutiniini tootmine, nii kleepuvad rakud kokku mõlema raku kasv peatub G1 faasis muutub rakuseina struktuur ja raku kuju

20
a-tüüpi rakkudes produtseeritakse: a-feromoni α-feromoni retseptorit ja vastupidi

21
MAT lookus Lookuses on 2 geeni (a1 ja a2 või α1 ja α2) MATa1 represseerib a-feromoni retseptori ja α-feromoni sünteesi MATa2 funktsioon teadmata MATα1 aktiveerib a-feromoni retseptori ja α-feromoni sünteesi MATα2 represseerib α-feromoni retseptori ja a-feromoni sünteesi
 MATa1 represseerib a-feromoni retseptori ja α-feromoni sünteesi MATa2 funktsioon teadmata MATα1 aktiveerib a-feromoni retseptori ja α-feromoni sünteesi MATα2 represseerib α-feromoni retseptori ja a-feromoni sünteesi")
22
Kui tuumad on ühinenud: Diploidid on alati MAT lookuse suhtes heterosügootsed a1 ja α2 valgud moodustavad heterodimeeri, mis aktiveerib meioosi ja sporulatsiooni ja represseerib haploidsed funktsioonid

23
Et asi oleks tõeliselt segane... Pärmi haploidsed rakud saavad oma ristumistüüpi muuta Kontrollib geen HO (endonukleaas) Dominantse alleeli korral toimub see peale igat raku jagunemist HMLMATHMR
 Dominantse alleeli korral toimub see peale igat raku jagunemist HMLMATHMR.")
24
(kott)seened mudelorganismidena Saccharomyces cerevisiae Aspergillus nidulans Schizosaccharomyces pombe Neurospora crassa
seened mudelorganismidena Saccharomyces cerevisiae Aspergillus nidulans Schizosaccharomyces pombe Neurospora crassa")
25
Saccharomyces cerevisiae kui mudelorganism Eukarüoot Lihtne kasvatada Väike genoom kergesti manipuleeritav Kasvab kiiresti Mutantide isoleerimise lihtsus Hästidefineeritud geneetiline süsteem Rekombinatsioon kõrge sagedusega

26
Saccharomyces cerevisiae kui mudelorganism Mitmekülgne DNA transformatsiooni süsteem Nii haploidne kui ka diploidne faas stabiilne Mittepatogeenne Metaboolsed ja regulatoorsed mehhanismid kõrgelt konserveerunud Inimeste haigustega seotud geenide ortoloogid...

27
Pärm: milliseid protsesse uuritakse? Geeniregulatsioon raku koostisosade biosüntees mRNA translatsioon valkude post-translatsiooniline modifitseerimine valkude sekreteerimine mitokondri biogenees võõrgeenide ekspressioon

28
Pärm: mida uuritakse? Rakutsükli regulatsioon Signaaliülekande mehhanismid Kromosoomi struktuur DNA replikatsioon Genoomika

29
Neurospora ja Aspergillus Haploidne vegetatiivne faas ja haploidsed koniidid - seeneniidistiku fenotüübi järgi saame teha järeldused genotüübi kohta

30
Neurospora

31
Aspergillus

32
Neurospora ja Aspergillus Iga eoskott sisaldab 8 eost peale meioosi toimub mitoos 2n tuuma iga üksik DNA ahel läheb ühe eose tuuma võimaldab algse DNA detailset analüüsi isegi heterodupleksi erinevused tulevad välja (osaline erinevus kaksikheeliksi ahelates)
")
33
Magnaporthe grisea Riisi närbumistõve tekitaja (rice blast fungus) Genoomi suurus 40.3 Mb 7 kromosoomi 10% genoomist kordusjärjestused (> 200 bp) retrotransposoonid sagedane rekombinatsioon
 Genoomi suurus 40.3 Mb 7 kromosoomi 10% genoomist kordusjärjestused (> 200 bp) retrotransposoonid sagedane rekombinatsioon")
34
Magnaporthe grisea apressor koniid

35
Magnaporthe grisea rekombinatsioonide tõttu genoom muutlik looduses paljuneb mittesuguliselt genoomi muutlikkus on seenele kasulik: peremehespetsiifilisuse muutumine kui kaotsi lähvad geenid, mis põhjustavad potentsiaalses peremehes kaitsereaktsioone

36
Seente transformatsioon Katsetatud paljudel seentel Head protokollid olemas üksikute jaoks: Saccharomyces Aspergillus Neurospora Ustilago

37
Võimalused transformatsiooniks Protoplast Terve raku töötlemine Ca 2+ ja Li + ioonidega Elektroporatsioon Geenipüss (gene gun, particle bombardment)
")
38
Mida on vaja transformatsiooni läbiviimiseks? Vektor, milles meid huvitaval geenil on sobiv promootor ja terminaator Seenerakud peavad olema kompetentsed Selektsioonisüsteem (Protoplastidest tuleb regenereerida mütseel)
.")
39
N. crassa – tetraadide analüüs Eoskotid kui korrastatud tetraadid - alleelide kombinatsioon eostes vastab meioosi I ja II jagunemisele meioos I - kahe (homoloogilise) kromosoomi lahknemine meioos II - kahe tütarkromatiidi lahknemine
 kromosoomi lahknemine meioos II - kahe tütarkromatiidi lahknemine.")
40
Tetraadide analüüs Saab jälgida rekombinatsiooni toimumist ja kindlaks teha aheldatuse gruppe Kottseened sobivad geenide meiootiliseks kaardistamiseks Pärmil on eoskotis 4 eost, mis on ühe meioosi tulemus Mitme alleeliga geenide aheldus Lookuse asukoht tsentromeeri suhtes

41
Tetraadide analüüs tänapäeval Et kindlaks teha mutatsiooni, mis vastab konkreetse lookuse muutmisele Et konstrueerida uusi tüvesid Et kindlaks teha geenide interaktsioone

42
Erinevad tetraaditüübid. Ristati AB × ab Eri tüüpide osakaal näitab, kas A ja B on aheldunud

43
Auksotroofid Auksotroofsed mutandid, kes pole ühe geeni muteerumise tõttu võimelised kasvama teatud söötmel Kasv taastub, kui söötmele lisada vajalik komponent Esineb spetsiifiline sõltuvus konkreetse geeni produkti ja metaboolse sammu vahel. “Üks geen - üks valk” hüpotees

44
Auksotroofne organism sisaldab mutatsiooni, mis muudab organismi toitumisvajadusi Prototroof – metsiktüüpi organism muutmata toitumisvajadustega

45
Pärmi auksotroofsed markerid Aminohapped Leu His Trp Nukleotiidid Uratsiil Adeniin Selektsioonivahend molek. geneetiliste katsete puhul

46
Mis on komplementatsioon? Kahe auksotroofi ristamisel saadud diploid on prototroof, kui defektid on erinevates lookustes Alleel, mis ühes haploidis on mutantne (retsessiivne), on teises haploidis funktsionaalne (dominantne) Mõlema haploidi wt alleelid kompenseerivad teise haploidi defektse alleeli
, on teises haploidis funktsionaalne (dominantne) Mõlema haploidi wt alleelid kompenseerivad teise haploidi defektse alleeli.")
47
Mis on komplementatsioon? Kui defektid on samades lookustes, siis kahe auksotroofi ristamisel tekkiv diploid on auksotroof Retsessiivne homosügoot Ka eri liikidelt pärit geenid võivad üksteist täiendada

48
Komplementatsiooni test Haploidsete auksotroofide genotüüpide kindlakstegemiseks Tundmatu genotüübiga haploid ristatakse teadaolevate genotüüpidega haploididega peavad olema erineva ristumistüübiga

49
Komplementatsioonigrupid Kui defektsete tüvede ristamisel ei toimu komplementatsiooni, kuuluvad nad samasse gruppi Iga grupp esindab ühte ensüümi biokeemilises rajas Suuremahulised projektid võimaldavad identifitseerida kõik kompl. grupid => kõik geenid metaboolses rajas Geeni defineerimine toime järgi

50
Alati pole kõik nii ilus… Kahe lookuse suhtes heterosügootne diploid võib fenotüübilt olla ikkagi defektne Alleelne ehk geenisisene komplementatsioon – mutatsioonid on mõlemas tüves samas lookuses

51
Geenisisene komplementatsioon Kui ensüüm koosneb kahest või rohkemast identsest subühikust Kaks erinevalt defektset valku kompenseerivad teineteise vead Kui ensüüm koosneb erinevatest domäänidest, mis täidavad erinevaid katalüütilisi funktsioone Pärmis on tavaline

52
Kas siis ikkagi 1 või 2 lookust? Tetraadide analüüs Rekombinatsiooni sagedus väga madal => mutatsioonid samas lookuses kuni 25% => mutatsioonid erinevates lookustes

53
S. cerevisiae genoom sekveneeriti 1996 Goffeau et al. 1996. Science 27 4: 546 esimene täielikult sekveneeritud eukarüootne genoom

54
Sekveneeritud seened Ascomycota Saccharomyces cerevisiae strain S288C12 Saccharomyces cerevisiae strain RM11-1a12 Saccharomyces pastorianus26 Candida guilliermondii12 Candida lusitaniae16 Candida tropicalis30 Candida glabrata12 Ashbya gossypii9 Debaryomyces hansenii10 Kluyveromyces lactis11 Pichia stipitis15 Yarrowia lipolytica21 Schizosaccharomyces pombe14 Aspergillus nidulans31 Aspergillus fumigatus32 Aspergillus oryzae37 Aspergillus niger37 Chaetomium globosum36 Coccidioides immitis29 Nectria haematococca52 Fusarium graminearum40 Fusarium verticillioides46 Magnaporthe grisea40 Neurospora crassa43 Stagonospora nodorum37 Trichoderma reesei33 Epichloë festucae35 Basidiomycota Coprinus cinereus 38 Cryptococcus neoformans20 Ustilago maydis20 Phanerochaete chrysosporium35 Zygomycota Rhizopus oryzae 40 Microsporidia Encephalitozoon cuniculi2.5 Dictyosteliomycota (Protozoa) Dictyostelium discoideum34 Oomycota (Chromista) Phytophthora ramorum65 Phytophthora sojae95 Mbp
 Dictyostelium discoideum34 Oomycota (Chromista) Phytophthora ramorum65 Phytophthora sojae95 Mbp")
55
Schizosaccharomyces pombe Genoomi suurus 13.8 Mb 3 kromosoomi Pooldub, mitte ei pungu Kasutatakse raku kasvamise ja jagunemise uurimiseks

56
S. cerevisiae genoom 12 Mb (~200 korda väiksem inimese genoomist, ~ 4 korda suurem E. coli genoomist) 16 kromosoomi Kromosoomide suurused 250-2500 kb ~ 6000 ORF-i
 16 kromosoomi Kromosoomide suurused kb ~ 6000 ORF-i.")
57
S. cerevisiae genoom Valke kodeerivad alad moodustavad 70% genoomist 3.8% ORF-e sisaldab introneid Järjestikku ~ 120 rRNA geenide komplekti kordust (XII kromosoomis)
.")
59
http://www.yeastgenome.org/cache/genomeSnapshot.html
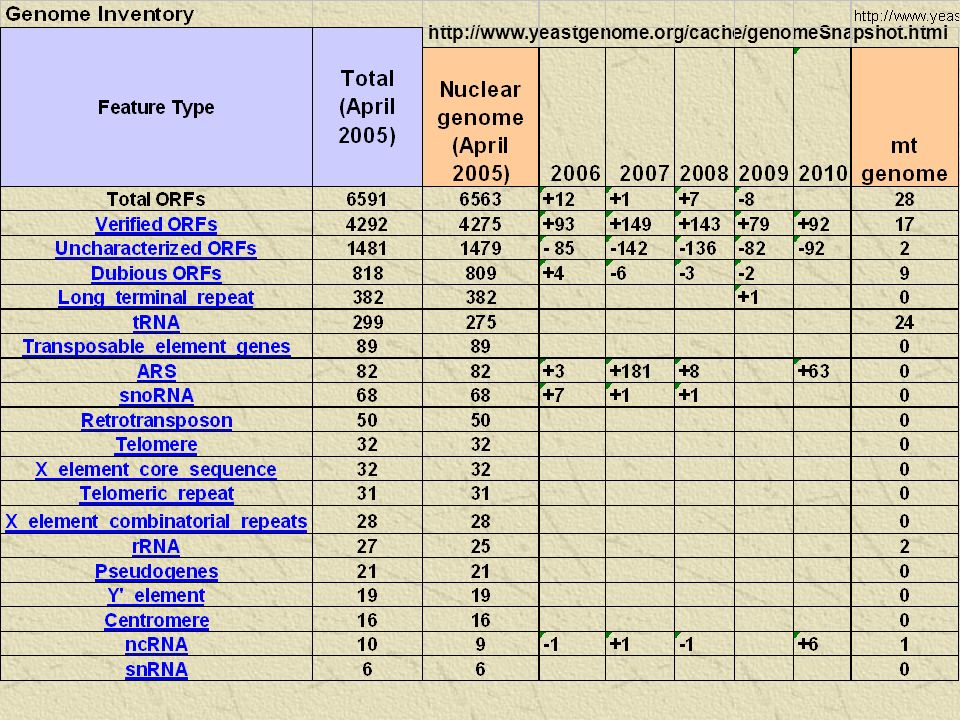
60
S. cerevisiae valke kodeerivad geenid

61
S. cerevisiae geeniproduktide jaotus vastavalt bioloogilistele protsessidele

62
S. cerevisiae kromosomaalsete geenide nomenklatuur Gene symbol Definition ARG + All wild-type alleles controlling arginine requirement ARG2 A locus or dominant allele arg2 A locus or recessive allele confering an arginine requirement arg2 – Any arg2 allele confering an arginine requirement ARG2 + The wild-type allele arg2-9 A specific allele or mutation Arg + A strain not requiring arginine Arg – A strain requiring arginine Arg2p The protein encoded by ARG2

63
S. cerevisiae mitokonder Mitokondris leiab aset suur hulk biokeemilisi reaktsioone Oksüdatiivse fosforüleerimise teel produtseeritakse ATP ja NAD + fakultatiivsed anaeroobid võivad elada ilma funktsionaalse hingamisahelata Pärmid on fakultatiivsed anaeroobid, mis tähendab, et nad saavad hakkama ka ilma molekulaarse hapnikuta => võivad elada ilma funktsionaalse hingamisahelata

64
Pärmi elu ilma hingamata Eeldab sobivat süsinikuallikat (suhkrud) ATP süntees glükolüüsi ja kääritamise käigus Rakud kasvavad aeglaselt, kolooniad on väikesed Kääritataval söötmel kasvab ka siis, kui mtDNA on mutantne või puudub üldse
 ATP süntees glükolüüsi ja kääritamise käigus Rakud kasvavad aeglaselt, kolooniad on väikesed Kääritataval söötmel kasvab ka siis, kui mtDNA on mutantne või puudub üldse")
65
Pärmi mitokondriaalne genoom Suurus ~ 80 kb (inimesel 16.5 kb) Lineaarne! Ühes haploidses pärmirakus ~ 50 mtDNA koopiat Kodeerib hingamisahela ensüüme Ülejäänud valgud kodeerib tuuma genoom Tsütoplasmaatiline pärilikkus

66
Pärmi mitokondriaalne genoom ehk ρ faktor Metsiktüüpi pärmirakud (‘grande’), mis hingavad normaalselt ja moodustavad suuri kolooniaid, on ρ + ‘Petite’ mutandid ei ole võimelised hingama ja nad moodustavad fermenteeritaval söötmel väikesi kolooniaid ρ 0 rakkudel puudub mtDNA üldse ρ - rakkudel esineb mtDNA, mis on moodustunud wt mitokondriaalse genoomi üksikutest fragmentidest, mis esinevad paljude kordustena
, mis hingavad normaalselt ja moodustavad suuri kolooniaid, on ρ + ‘Petite’ mutandid ei ole võimelised hingama ja nad moodustavad fermenteeritaval söötmel väikesi kolooniaid ρ 0 rakkudel puudub mtDNA üldse ρ - rakkudel esineb mtDNA, mis on moodustunud wt mitokondriaalse genoomi üksikutest fragmentidest, mis esinevad paljude kordustena")
67
‘petite’ - mutandid Saab eristada, ristates neid wt rakkudega ρ 0 rakud annavad wt (‘grande’) järglased keskmiselt supressiivsete ρ - rakkude järglased on pooled ‘grande’ ja pooled ‘petite’ hüpersupressiivsete ρ - (HS) rakkude järglased on peaaegu kõik ‘petite’
 järglased keskmiselt supressiivsete ρ - rakkude järglased on pooled ‘grande’ ja pooled ‘petite’ hüpersupressiivsete ρ - (HS) rakkude järglased on peaaegu kõik ‘petite’")
68
ρ - mtDNA Sisaldab wt mtDNA üksikuid fragmente, mis esinevad paljude kordustena HS ρ - mutantidel on selliseks fragmendiks arvatavasti replikatsiooni alguskoht (ori)
")
69
Pärm ja genoomiuuringud DNA kiipide kasutamine transkriptoomi uurimiseks Geeni funktsioonide analüüs geeni väljalülitamise teel Valkude lokalisatsiooni analüüs 2D protein maps Ensümaatilise aktiivsuse analüüs Valk-valk interaktsioonide analüüs

70
Pärmid ja proteoomika Geenijärjestuste analüüs vs. valkude analüüs Valgud ei tegutse tavaliselt üksinda Valgukomplekside analüüs

71
Pärmi proteoomi 2D-kaart

72
Kuidas analüüsida valk-valk interaktsioone? Pärmi kaksikhübriidsüsteem mRNA sünekspressioon Geneetilised interaktsioonid ehk sünteetiline letaalsus In silico meetodid Valgukomplekside afiinsuspuhastamine + mass-spektromeetria

73
Suguline paljunemine Seened mudelorganismidena Tetraadide analüüs Komplementatsiooni test S. cerevisiae genoom Proteoomika

Similar presentations

 CNRS / Aix-Marseille Université,>")


















